
Radioprotective Effect of Whey Hydrolysate Peptides against γ-Radiation-Induced Oxidative Stress in BALB/c Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Animals and Treatments
2.3. Irradiation Process
2.4. Experimental Assays
2.4.1. Survival Experiment
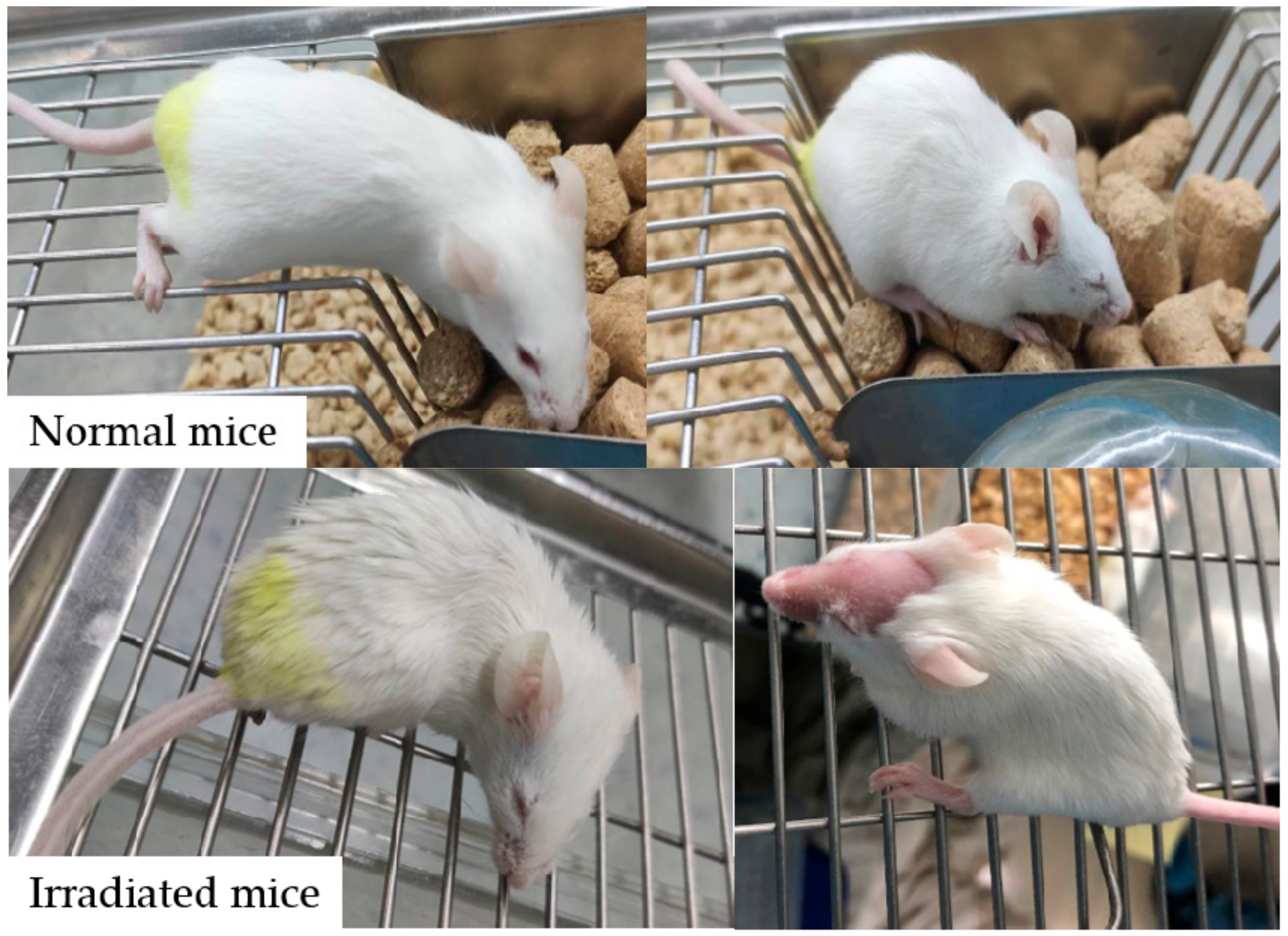
2.4.2. General Status, Body Weight and Organ Index
2.4.3. Peripheral White Blood Cell Count and Bone Marrow DNA Content
2.4.4. Analysis of Superoxide Dismutase (SOD), Glutathione Peroxidase (GSH-PX) and Malondialdehyde (MDA) Levels
2.4.5. Analysis of Tumor Necrosis Factor Alpha (TNF-α) and Interleukin-6 (IL-6) Levels
2.4.6. Histopathology of Mouse Small Intestine
2.4.7. Expression of Occludin and Zona Occludens 1(ZO-1) in Intestine
2.4.8. Analysis of Serum D-Lactic Acid, Diamine Oxidase (DAO) and Lipopolysaccharide (LPS) Levels
2.5. Statistical Analysis
3. Results
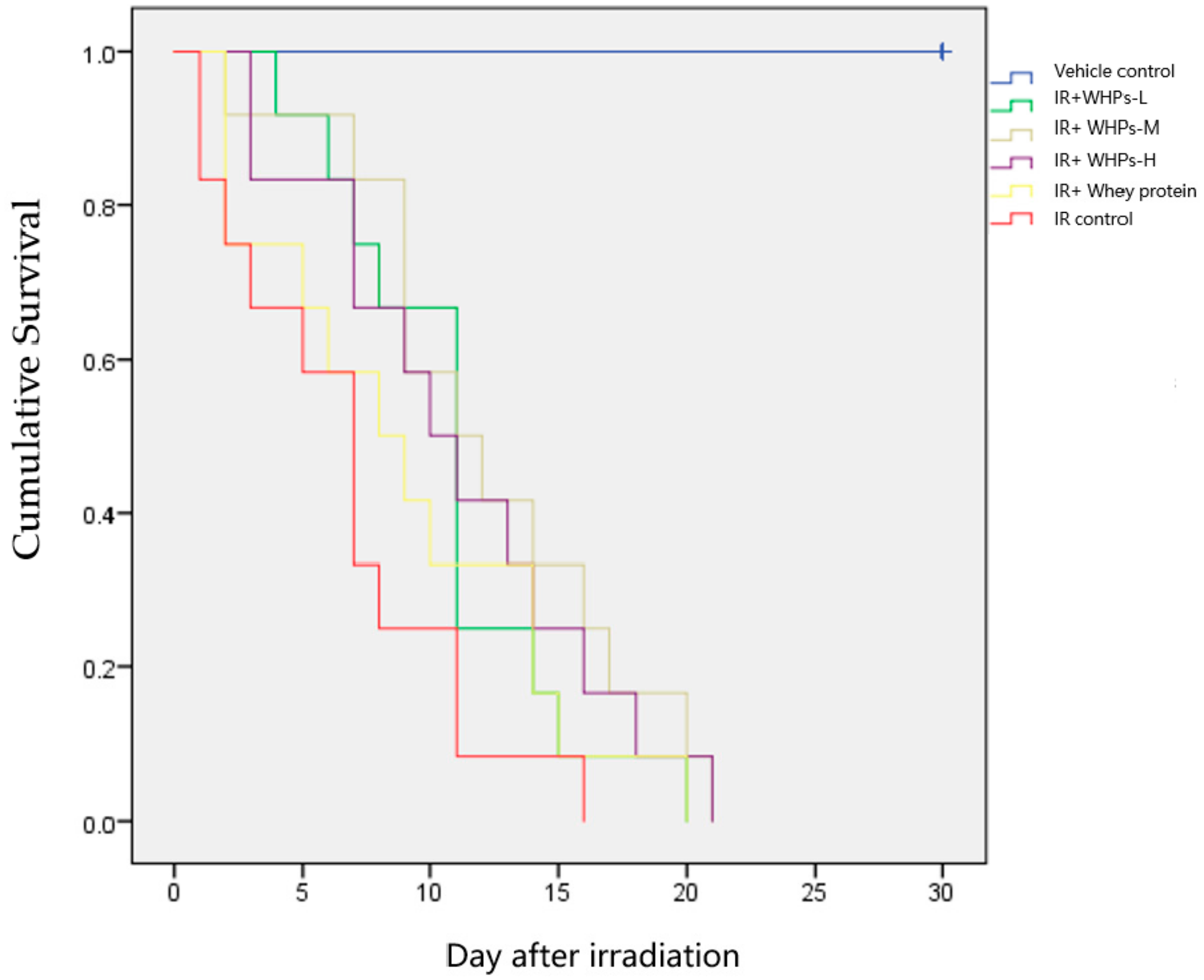
3.1. WHPs Prolong the Survival Time of Irradiated Mice
3.2. WHPs Restore Body Weight in Irradiated Mice
3.3. WHPs Accelerate the Recovery of Peripheral WBC in Irradiated Mice
3.4. WHPs Promote the Recovery of Bone Marrow DNA Damage in Irradiated Mice
3.5. WHPs Improve the Antioxidant Capacity of Irradiated Mice
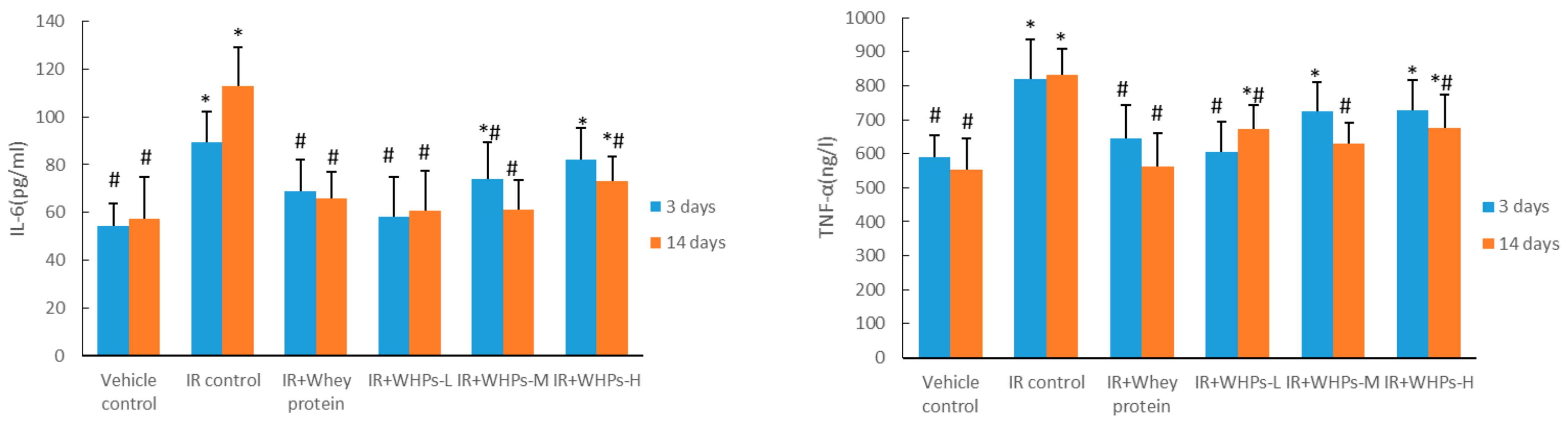
3.6. WHPs Regulate the Level of Inflammatory Cytokines in Irradiated Mice
3.7. WHPs Protect Intestinal Morphology and Structure in Irradiated Mice
3.8. WHPs Increase Intestinal Tight Junction Protein Expression in the Acute Phase of Radiation Injury
3.9. Effect of WHPs on Permeability in Irradiated Mice Intestinal Permeability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, K.L.; Marks, L.B.; Sibley, G.S.; Clough, R.W.; Garst, J.L.; Crawford, J.; Shafman, T.D. Routine use of approximately 60 Gy once-daily thoracic irradiation for patients with limited-stage small-cell lung cancer. Int. J. Radiat. Oncol. Biol. Phys. 2003, 56, 355–359. [Google Scholar] [CrossRef]
- Roach, M.C.; Bradley, J.D.; Robinson, C.G. Optimizing radiation dose and fractionation for the definitive treatment of locally advanced non-small cell lung cancer. J. Thorac. Dis. 2018, S2465–S2473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, D.S.; Lasley, F.D.; Das, I.J.; Mendonca, M.S.; Dynlacht, J.R. Acute Effects of Total Body Irradiation (TBI). In Basic Radiotherapy Physics and Biology; Springer: Cham, Switzerland, 2014; pp. 259–263. [Google Scholar]
- Mishra, K.N.; Moftah, B.A.; Alsbeih, G.A. Appraisal of mechanisms of radioprotection and therapeutic approaches of radiation countermeasures. Biomed. Pharmacother. 2018, 106, 610–617. [Google Scholar] [CrossRef]
- Jakubczyk, A.; Karaś, M.; Rybczyńska-Tkaczyk, K.; Zielińska, E.; Zieliński, D. Current Trends of Bioactive Peptides—New Sources and Therapeutic Effect. Foods 2020, 9, 846. [Google Scholar] [CrossRef] [PubMed]
- Arends, J.; Bachmann, P.; Baracos, V.; Barthelemy, N.; Bertz, H.; Bozzetti, F.; Fearon, K.; Hütterer, E.; Isenring, E.; Kaasa, S. ESPEN guidelines on nutrition in cancer patients. Clin. Nutr. 2017, 36, 11–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González de Llano, D.; Polo Sánchez, C. PEPTIDES. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 4468–4473. [Google Scholar]
- Semen, Z.; Arif, A. Bioactive Peptides in Milk and Its Biological Importance. J. Turk. Vet. Med. Assoc. 2016, 15, 67–84. [Google Scholar]
- Nakamura, K.; Sasayama, A.; Takahashi, T.; Yamaji, T. An Immune-Modulating Diet in Combination with Chemotherapy Prevents Cancer Cachexia by Attenuating Systemic Inflammation in Colon 26 Tumor-Bearing Mice. Nutr. Cancer 2015, 67, 912–920. [Google Scholar] [CrossRef]
- Saint-Sauveur, D.; Gauthier, S.F.; Boutin, Y.; Montoni, A. Immunomodulating properties of a whey protein isolate, its enzymatic digest and peptide fractions. Int. Dairy J. 2008, 18, 260–270. [Google Scholar] [CrossRef]
- Kunwar, A.; Verma, P.; Bhilwade, H.N.; Iwaoka, M.; Priyadarsini, K.I. Dihydroxyselenolane (DHS) supplementation improves survival following whole-body irradiation (WBI) by suppressing tissue-specific inflammatory responses. Mutat Res. Genet. Toxicol. Environ. Mutagen. 2016, 807, 33–46. [Google Scholar] [CrossRef]
- Raikos, V.; Dassios, T. Health-promoting properties of bioactive peptides derived from milk proteins in infant food: A review. Dairy Sci. Technol. 2014, 94, 91–101. [Google Scholar] [CrossRef] [Green Version]
- Kindekov, I.; Mileva, M.; Krastev, D.; Vassilieva, V.; Raynova, Y.; Doumanova, L.; Aljakov, M.; Idakieva, K. Radioprotective effect of Rapana thomasiana hemocyanin in gamma induced acute radiation syndrome. Biotechnol. Biotec. Eq. 2014, 28, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Verma, P.; Kunwar, A.; Indira Priyadarsini, K. Effect of Low-Dose Selenium Supplementation on the Genotoxicity, Tissue Injury and Survival of Mice Exposed to Acute Whole-Body Irradiation. Biol. Trace Elem. Res. 2017, 179, 130–139. [Google Scholar] [CrossRef]
- Khan, S.; Adhikari, J.S.; Rizvi, M.A.; Chaudhury, N.K. Radioprotective potential of melatonin against 60Co γ-ray-induced testicular injury in male C57BL/6 mice. J. Biomed. Sci. 2015, 22, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, T.; Vares, G.; Ninomiya, Y.; Wang, B.; Katsube, T.; Tanaka, K.; Maruyama, K.; Nenoi, M. Diallyl Disulfide Mitigates DNA Damage and Spleen Tissue Effects After Irradiation. Med. Sci. Monit. 2019, 25, 8920–8927. [Google Scholar] [CrossRef] [PubMed]
- Gui, C.; Cheng, Z.; Cecil, E.; Sloan, L.; Deek, M.P.; Lakshminarayanan, P.; Guo, Y.; Han, P.; McNutt, T.R.; Quon, H. Low Peripheral Blood Leukocyte Counts during Radiation Therapy for Head and Neck Cancer Predict Distant Relapse and Dysphagia. Int. J. Radiat. Oncol. Biol. Phys. 2019, 105, E421. [Google Scholar] [CrossRef]
- Nilsson, R.; Liu, N.-A. Nuclear DNA damages generated by reactive oxygen molecules (ROS) under oxidative stress and their relevance to human cancers, including ionizing radiation-induced neoplasia part II: Relation between ROS-induced DNA damages and human cancer. Radiat. Med. Prot. 2020, 1, 196–204. [Google Scholar] [CrossRef]
- Daly, M.J. Death by protein damage in irradiated cells. DNA Repair 2012, 11, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Wang, N.; Dong, X.; Wang, C.; Du, Z.; Mei, L.; Yong, Y.; Huang, C.; Li, Y.; Gu, Z.; et al. Graphdiyne Nanoparticles with High Free Radical Scavenging Activity for Radiation Protection. ACS Appl. Mater. Inter. 2019, 11, 2579–2590. [Google Scholar] [CrossRef]
- Wang, X.-Q.; Wang, W.; Peng, M.; Zhang, X.-Z. Free radicals for cancer theranostics. Biomaterials 2021, 266. [Google Scholar] [CrossRef]
- Shaban, N.Z.; Ahmed Zahran, A.M.; El-Rashidy, F.H.; Abdo Kodous, A.S. Protective role of hesperidin against γ-radiation-induced oxidative stress and apoptosis in rat testis. J. Biol. Res. Thessalon 2017, 24, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishanth, R.P.; Jyotsna, R.G.; Schlager, J.J.; Hussain, S.M.; Reddanna, P. Inflammatory responses of RAW 264.7 macrophages upon exposure to nanoparticles: Role of ROS-NFκB signaling pathway. Nanotoxicology 2011, 5, 502–516. [Google Scholar] [CrossRef] [PubMed]
- Gorbunov, N.V.; Sharma, P. Protracted Oxidative Alterations in the Mechanism of Hematopoietic Acute Radiation Syndrome. Antioxidants 2015, 4, 134–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, R.P.; Fouquerel, E.; Opresko, P.L. The impact of oxidative DNA damage and stress on telomere homeostasis. Mech. Ageing Dev. 2019, 177, 37–45. [Google Scholar] [CrossRef]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef]
- Kantara, C.; Moya, S.M.; Houchen, C.W.; Umar, S.; Ullrich, R.L.; Singh, P.; Carney, D.H. Novel regenerative peptide TP508 mitigates radiation-induced gastrointestinal damage by activating stem cells and preserving crypt integrity. Lab. Investig. 2015, 95, 1222–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homma, T.; Fujii, J. Application of Glutathione as Anti-Oxidative and Anti-Aging Drugs. Curr. Drug Metab. 2015, 16, 560–571. [Google Scholar] [CrossRef]
- Bounous, G. Whey protein concentrate (WPC) and glutathione modulation in cancer treatment. Anticancer Res. 2000, 20, 4785–4792. [Google Scholar]
- Power, O.; Jakeman, P.; FitzGerald, R.J. Antioxidative peptides: Enzymatic production, in vitro and in vivo antioxidant activity and potential applications of milk-derived antioxidative peptides. Amino Acids 2013, 44, 797–820. [Google Scholar] [CrossRef]
- Kleekayai, T.; Cermeño, M.; FitzGerald, R.J. The Production of Bioactive Peptides from Milk Proteins. In Agents of Change: Enzymes in Milk and Dairy Products; Kelly, A.L., Larsen, L.B., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 447–497. [Google Scholar]
- Mohammadian, M.; Salami, M.; Emam-Djomeh, Z.; Alavi, F. Chapter 25—Nutraceutical Properties of Dairy Bioactive Peptides. In Dairy in Human Health and Disease across the Lifespan; Watson, R.R., Collier, R.J., Preedy, V.R., Eds.; Academic Press: Oxford, UK, 2017; pp. 325–342. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Content (g/100 g) |
|---|---|
| Glutamic Acid | 14.4 |
| Aspartic Acid | 8.9 |
| Leucine | 8.6 |
| Lysine | 8.1 |
| Threonine | 5.8 |
| Isoleucine | 5.4 |
| Proline | 5.0 |
| Valine | 4.6 |
| Alanine | 4.3 |
| Serine | 4.2 |
| Phenylalanine | 2.6 |
| Tyrosine | 2.5 |
| Arginine | 2.3 |
| Cystine/Cysteine | 2.0 |
| Methionine | 1.8 |
| Tryptophan | 1.7 |
| Glycine | 1.5 |
| Histidine | 1.5 |
| Hydroxyproline | <0.1 |
| Group | n | Median | Q25 | Q75 |
|---|---|---|---|---|
| Vehicle control | 12 | 30 # | 30 | 30 |
| IR control | 12 | 7 * | 2 | 8 |
| IR+Whey protein | 12 | 8 * | 2 | 14 |
| IR+WHPs-L | 12 | 11 * | 7 | 11 |
| IR+WHPs-M | 12 | 11 *# | 9 | 16 |
| IR+WHPs-H | 12 | 10 * | 7 | 14 |
| Group | 3rd Day Post-Irradiation | 14th Day Post-Irradiation | ||
|---|---|---|---|---|
| Initial Weight (g) | Final Weight (g) | Initial Weight (g) | Final Weight (g) | |
| Vehicle control | 21.25 ± 0.75 | 21.90 ± 2.02 | 21.18 ± 0.98 | 20.60 ± 0.91 # |
| IR control | 21.26 ± 1.12 | 20.68 ± 1.24 | 20.91 ± 2.31 | 18.59 ± 1.75 * |
| IR+Whey protein | 21.21 ± 1.12 | 20.70 ± 1.91 | 20.98 ± 2.31 | 18.72 ± 1.09 |
| IR+WHPs-L | 21.23 ± 0.83 | 20.73 ± 2.10 | 21.02 ± 2.06 | 18.93 ± 1.17 |
| IR+WHPs-M | 21.35 ± 0.61 | 21.42 ± 1.58 | 20.84 ± 2.33 | 19.73 ± 1.45 # |
| IR+WHPs-H | 21.23 ± 1.01 | 21.72 ± 1.35 | 21.00 ± 1.56 | 19.16 ± 1.85 |
| Group | Spleen Index (mg/g) | Thymus Index (mg/g) | Liver Index (mg/g) |
|---|---|---|---|
| Vehicle control | 4.05 ± 0.31 # | 2.15 ± 0.61 # | 43.27 ± 3.18 |
| IR control | 1.69 ± 0.11 * | 0.61 ± 0.18 * | 42.38 ± 2.69 |
| IR+Whey protein | 1.67 ± 0.18 * | 0.74 ± 0.16 * | 42.32 ± 3.78 |
| IR+WHPs-L | 1.77 ± 0.35 * | 0.78 ± 0.27 * | 44.10 ± 9.51 |
| IR+WHPs-M | 1.62 ± 0.14 * | 0.85 ± 0.15 * | 41.04 ± 2.16 |
| IR+WHPs-H | 1.62 ± 0.19 * | 0.72 ± 0.17 * | 40.86 ± 3.78 |
| Group | Spleen Index (mg/g) | Thymus Index (mg/g) | Liver Index (mg/g) |
|---|---|---|---|
| Vehicle control | 4.31 ± 0.50 | 2.05 ± 0.33 | 45.68 ± 4.81 |
| IR control | 4.94 ± 1.09 | 1.96 ± 0.39 | 51.73 ± 9.46 |
| IR+Whey protein | 5.26 ± 0.91 * | 2.21 ± 0.59 | 50.36 ± 6.73 |
| IR+WHPs-L | 4.68 ± 1.62 | 2.21 ± 0.84 | 43.02 ± 14.50 # |
| IR+WHPs-M | 4.64 ± 0.87 | 2.06 ± 0.49 | 45.46 ± 6.93 |
| IR+WHPs-H | 4.53 ± 1.10 | 2.15 ± 0.56 | 44.96 ± 5.36 |
| Group | Number of WBCs at 3rd Day Post-Irradiation (109/mL) | Number of WBCs at 14th Day Post-Irradiation (109/mL) |
|---|---|---|
| Vehicle control | 11.5 ± 3.2 # | 10.5 ± 1.2 # |
| IR control | 1.8 ± 0.6 * | 8.4 ± 2.1 * |
| IR+Whey protein | 2.2 ± 0.6 * | 9.1 ± 1.9 |
| IR+WHPs-L | 2.2 ± 0.6 * | 8.9 ± 1.3 |
| IR+WHPs-M | 2.3 ± 0.8 * | 9.1 ± 1.7 |
| IR+WHPs-H | 2.0 ± 0.0 * | 10.2 ± 2.3 # |
| Group | Liver SOD (ng/g) | Serum SOD (ng/mL) | Liver GSH-PX (ng/g) | Serum GSH-PX (ng/mL) | Liver MDA (nmol/g) | Serum MDA (nmol/mL) |
|---|---|---|---|---|---|---|
| Vehicle control | 76.18 ± 12.74 # | 9.00 ± 1.33 # | 1403.24 ± 226.73 # | 165.04 ± 20.93 # | 80.63 ± 18.77 # | 10.30 ± 1.83 # |
| IR control | 45.95 ± 11.78 * | 4.94 ± 0.46 * | 732.13 ± 206.88 * | 83.81 ± 14.01 * | 137.37 ± 19.42 * | 15.72 ± 2.43 * |
| IR+Whey protein | 67.77 ± 12.29 # | 6.60 ± 1.06 *# | 1155.86 ± 255.98 *# | 125.71 ± 25.19 *# | 115.08 ± 19.92 *# | 11.90 ± 0.91 # |
| IR+WHPs-L | 60.37 ± 10.43 *# | 8.10 ± 1.26 # | 1268.84 ± 234.70 # | 132.78 ± 23.91 *# | 86.68 ± 21.02 # | 13.70 ± 0.90 *# |
| IR+WHPs-M | 54.05 ± 10.80 * | 7.26 ± 0.81 *# | 1045.10 ± 81.61 *# | 115.8 ± 22.75 *# | 104.01 ± 20.10 *# | 13.66 ± 2.39 *# |
| IR+WHPs-H | 53.26 ± 7.48 * | 6.24 ± 1.36 *# | 909.92 ± 86.49 * | 114.33 ± 18.3 *# | 105.84 ± 17.11 *# | 13.75 ± 1.73 * |
| Group | Liver SOD (ng/g) | Serum SOD (ng/mL) | Liver GSH-PX (ng/g) | Serum GSH-PX (ng/mL) | Liver MDA (nmol/g) | Serum MDA (nmol/mL) |
|---|---|---|---|---|---|---|
| Vehicle control | 72.75 ± 16.01 # | 8.07 ± 1.11 # | 1351.07 ± 201.14 # | 155.12 ± 13.32 # | 97.39 ± 11.80 # | 10.48 ± 2.74 # |
| IR control | 39.18 ± 12.17 * | 4.74 ± 1.06 * | 558.30 ± 209.47 * | 90.73 ± 16.65 * | 152.20 ± 15.03 * | 16.81 ± 2.51 * |
| IR+Whey protein | 63.24 ± 5.50 # | 7.57 ± 1.66 # | 1067.09 ± 130.32 *# | 133.32 ± 7.11 *# | 104.52 ± 17.50 # | 11.84 ± 2.03 # |
| IR+WHPs-L | 73.07 ± 12.91 # | 8.94 ± 1.11 # | 1227.39 ± 174.08 # | 150.04 ± 21.71 # | 92.22 ± 14.96 # | 12.01 ± 1.71 # |
| IR+WHPs-M | 63.71 ± 10.46 # | 7.24 ± 1.20 # | 1166.78 ± 219.12 # | 130.57 ± 26.04 *# | 94.53 ± 22.25 # | 13.21 ± 1.92 *# |
| IR+WHPs-H | 56.39 ±12.65 *# | 6.69 ± 1.28 *# | 992.34 ± 193.28 *# | 122.71 ± 16.75 *# | 116.90 ± 15.73 *# | 12.01 ± 1.50 # |
| Group | Villus Height (3 days) | Villus Height (14 days) | Crypt Depth (3 days) | Crypt Depth (14 days) |
|---|---|---|---|---|
| Vehicle control | 311.5 ± 30.8 | 377.6 ± 43.6 | 81.8 ± 10.4 | 71.0 ± 3.0 # |
| IR control | 281.7 ± 22.2 | 336.7 ± 19.3 | 87.6 ± 10.8 | 90.1 ± 15.0 * |
| IR+Whey protein | 330.5 ± 29.9 # | 381.6 ± 43.1 | 85.8 ± 10.8 | 81.1 ± 8.4 |
| IR+WHPs-L | 303.2 ± 31.0 | 350.4 ± 51.2 | 89.9 ± 8.6 | 88.5 ± 12.1 * |
| IR+WHPs-M | 315.2 ± 41.4 | 369.1 ± 31.0 | 92.4 ± 12.2 | 84.3 ± 15.8 |
| IR+WHPs-H | 313.9 ± 39.7 # | 364.4 ± 52.5 | 77.7 ± 15.0 # | 92.0 ± 13.8 * |
| Group | Occludin (3 days) | ZO-1 (3 days) | Occludin (14 days) | ZO-1 (14 days) |
|---|---|---|---|---|
| Vehicle control | 0.175 ± 0.016 # | 0.165 ± 0.010 # | 0.181 ± 0.011 | 0.164 ± 0.010 |
| IR control | 0.156 ± 0.010 * | 0.152 ± 0.008 * | 0.170 ± 0.007 | 0.155 ± 0.011 |
| IR+Whey protein | 0.162 ± 0.008 | 0.149 ± 0.016 * | 0.172 ± 0.022 | 0.155 ± 0.013 |
| IR+WHPs-L | 0.163 ± 0.013 | 0.164 ± 0.009 | 0.186 ± 0.027 | 0.163 ± 0.013 |
| IR+WHPs-M | 0.168 ± 0.024 | 0.168 ± 0.020 # | 0.184 ± 0.042 | 0.160 ± 0.014 |
| IR+WHPs-H | 0.165 ± 0.011 | 0.162 ± 0.007 | 0.178 ± 0.020 | 0.161 ± 0.021 |
| Group | D-Lactate (μmol/L) | DAO (pg/mL) | LPS (EU/L) |
|---|---|---|---|
| Vehicle control | 31.58 ± 5.96 | 757.54 ± 265.71 | 14.52 ± 2.46 |
| IR control | 49.36 ± 4.27 | 1398.84 ± 166.12 | 23.12 ± 2.14 |
| IR+Whey protein | 39.02 ± 6.53 | 1069.59 ± 187.01 | 19.97 ± 3.11 |
| IR+WHPs-L | 40.53 ± 4.49 | 969.10 ± 185.02 | 17.90 ± 2.80 |
| IR+WHPs-M | 40.04 ± 4.18 | 1068.36 ± 167.25 | 20.46 ± 2.69 |
| IR+WHPs-H | 42.54 ± 5.20 | 1123.73 ± 238.61 | 19.18 ± 2.63 |
| Group | D-Lactate (μmol/L) | DAO (pg/mL) | LPS (EU/L) |
|---|---|---|---|
| Vehicle control | 33.58 ± 4.01 | 796.50 ± 206.96 | 14.82 ± 3.12 |
| IR control | 50.29 ± 6.06 | 1490.52 ± 237.45 | 23.49 ± 1.80 |
| IR+Whey protein | 36.59 ± 4.60 | 767.24 ± 175.31 | 17.93 ± 2.32 |
| IR+WHPs-L | 35.23 ± 6.23 | 924.16 ± 233.64 | 18.56 ± 1.88 |
| IR+WHPs-M | 38.01 ± 5.36 | 976.59 ± 197.92 | 18.51 ± 2.68 |
| IR+WHPs-H | 38.93 ± 6.06 | 965.65 ± 101.67 | 19.44 ± 1.82 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.-R.; Zhu, N.; Hao, Y.-T.; Yu, X.-C.; Li, Z.; Mao, R.-X.; Liu, R.; Kang, J.-W.; Hu, J.-N.; Li, Y. Radioprotective Effect of Whey Hydrolysate Peptides against γ-Radiation-Induced Oxidative Stress in BALB/c Mice. Nutrients 2021, 13, 816. https://doi.org/10.3390/nu13030816
Liu X-R, Zhu N, Hao Y-T, Yu X-C, Li Z, Mao R-X, Liu R, Kang J-W, Hu J-N, Li Y. Radioprotective Effect of Whey Hydrolysate Peptides against γ-Radiation-Induced Oxidative Stress in BALB/c Mice. Nutrients. 2021; 13(3):816. https://doi.org/10.3390/nu13030816
Chicago/Turabian StyleLiu, Xin-Ran, Na Zhu, Yun-Tao Hao, Xiao-Chen Yu, Zhen Li, Rui-Xue Mao, Rui Liu, Jia-Wei Kang, Jia-Ni Hu, and Yong Li. 2021. "Radioprotective Effect of Whey Hydrolysate Peptides against γ-Radiation-Induced Oxidative Stress in BALB/c Mice" Nutrients 13, no. 3: 816. https://doi.org/10.3390/nu13030816
APA StyleLiu, X. -R., Zhu, N., Hao, Y. -T., Yu, X. -C., Li, Z., Mao, R. -X., Liu, R., Kang, J. -W., Hu, J. -N., & Li, Y. (2021). Radioprotective Effect of Whey Hydrolysate Peptides against γ-Radiation-Induced Oxidative Stress in BALB/c Mice. Nutrients, 13(3), 816. https://doi.org/10.3390/nu13030816