
Dietary Supplementation with Fermented Brassica rapa L. Stimulates Defecation Accompanying Change in Colonic Bacterial Community Structure


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Fermented B. rapa L. for 2 Experiments
2.2. Subjects and Study Design of Human Volunteer Study
2.3. Serum Biochemistry and Hematology Analyses
2.4. Preparation of Peripheral Blood Mononuclear Cells
2.5. Flow Cytometric Analysis
2.6. Natural Killer Cell Activity
2.7. Bowel Function Questionnaire
2.8. Mouse Experiment
2.9. Measurement of Cytokine Production from Mouse Spleen Cells
2.10. Microbial Analysis
2.11. Statistical Analysis
3. Results
3.1. General Summary of Biochemical Characteristics
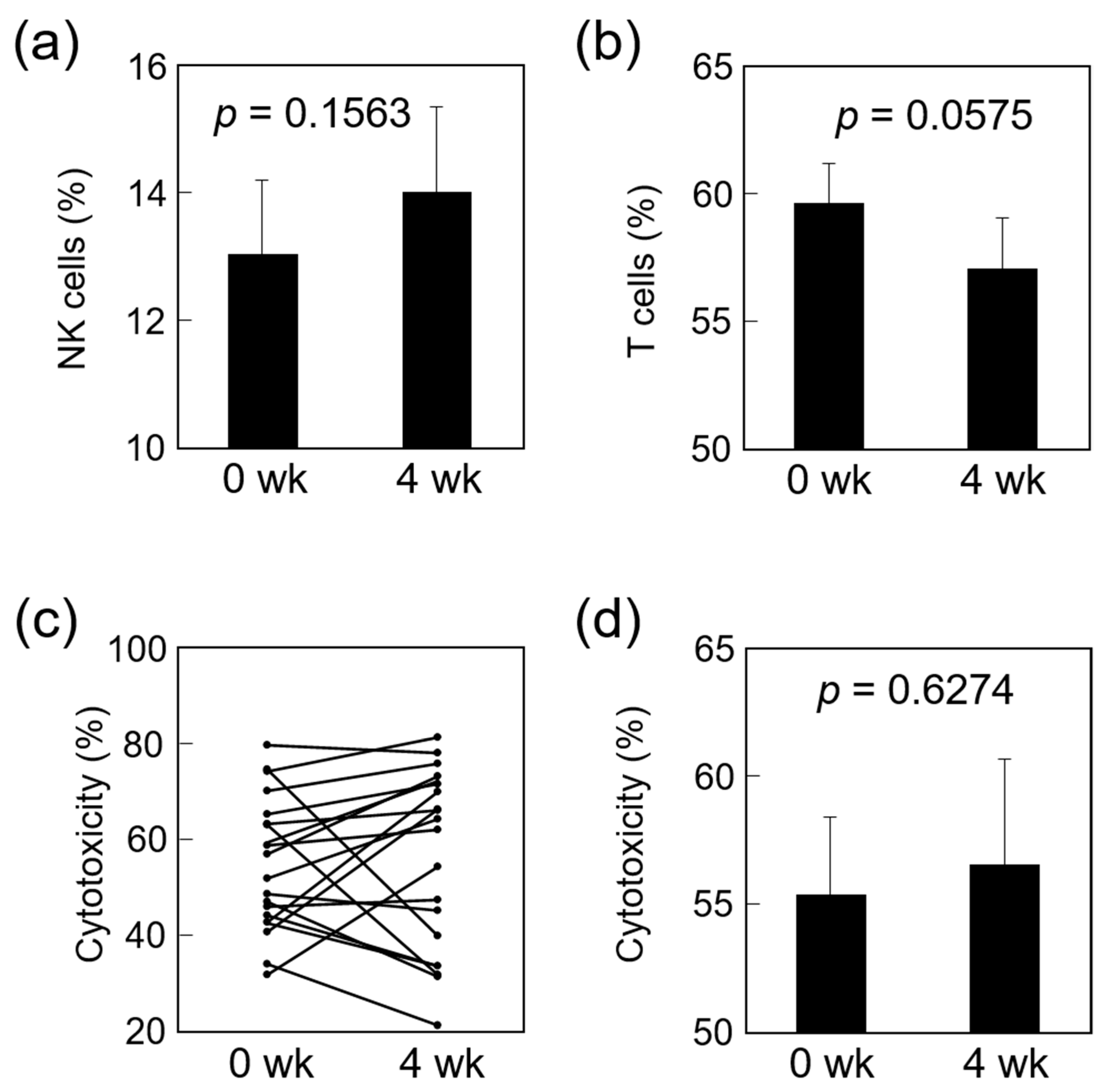
3.2. Effects on Immune Function of Study Subjects
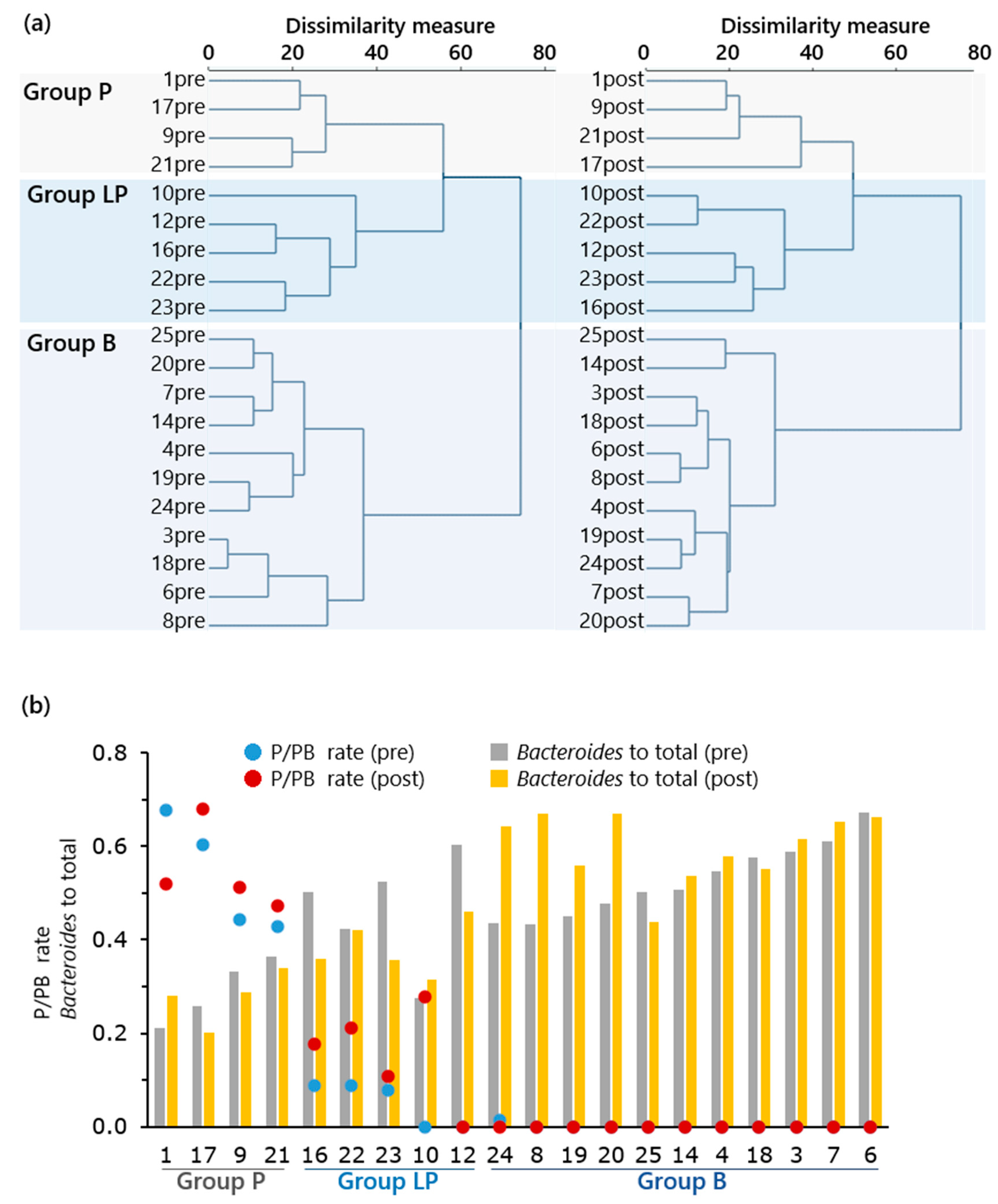
3.3. Effects on Stool Microbiota and Bowel Function of Study Subjects
3.4. Mouse Experiment
4. Discussion
4.1. Implications from Results of Volunteer Study
4.2. Detail Analysis of the Effect of Feeding Fermented B. rapa L. on Gut Bacteria in Mouse
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oikawa, T.; Uneyama, H. WASHOKU and health: New approach with dashi/umami in the medical and nutritional health care. Yakugaku zasshi: J. Pharm. Soc. Jpn. 2016, 136, 1325–1326. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Yamamoto, K.; Yamada, K.; Furuya, K.; Uyeno, Y. Relationship of enhanced butyrate production by colonic butyrate-producing bacteria to immunomodulatory effects in normal mice fed an insoluble fraction of Brassica rapa L. Appl. Environ. Microbiol. 2016, 82, 2693–2699. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Yamamoto, K.; Hamajima, C.; Takahashi, F.; Yamada, K.; Furuya, K.; Uyeno, Y. Changes in gut microbial ecology and immunological responses of mice fed the insoluble fraction of Brassica rapa L. that was fermented or not. Microbes Environ. 2017, 32, 268–274. [Google Scholar] [CrossRef] [Green Version]
- Shida, K.; Suzuki, T.; Kiyoshima-Shibata, J.; Shimada, S.-I.; Nanno, M. Essential roles of monocytes in stimulating human peripheral blood mononuclear cells with Lactobacillus casei to produce cytokines and augment natural killer cell activity. Clin. Vaccine Immunol. 2006, 13, 997–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, S.J.; Heaton, K.W. Stool form scale as a useful guide to intestinal transit time. Scand. J. Gastroenterol. 1997, 32, 920–924. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, K.; Hisada, T.; Sato, M.; Mochizuki, J. Application of new primer-enzyme combinations to terminal restriction fragment length polymorphism profiling of bacterial populations in human feces. Appl. Environ. Microbiol. 2003, 69, 1251–1262. [Google Scholar] [CrossRef] [Green Version]
- Uyeno, Y.; Akiyama, K.; Hasunuma, T.; Yamamoto, H.; Yokokawa, H.; Yamaguchi, T.; Kawashima, K.; Itoh, M.; Kushibiki, S.; Hirako, M. Effects of supplementing an active dry yeast product on rumen microbial community composition and on subsequent rumen fermentation of lactating cows in the mid-to-late lactation period. Anim. Sci. J. 2017, 88, 119–124. [Google Scholar] [CrossRef]
- Gorvitovskaia, A.; Holmes, S.P.; Huse, S.M. Interpreting Prevotella and Bacteroides as biomarkers of diet and lifestyle. Microbiome 2016, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Kurasawa, S.i.; Haack, V.S.; Marlett, J.A. Plant residue and bacteria as bases for increased stool weight accompanying consumption of higher dietary fiber diets. J. Am. Coll. Nutr. 2000, 19, 426–433. [Google Scholar] [CrossRef]
- Yang, J.; Wang, H.-P.; Zhou, L.; Xu, C.-F. Effect of dietary fiber on constipation: A meta analysis. World J. Gastroenterol. WJG 2012, 18, 7378. [Google Scholar] [CrossRef]
- Uyeno, Y.; Katayama, S.; Nakamura, S. Changes in mouse gastrointestinal microbial ecology with ingestion of kale. Benef. Microbes 2014, 5, 345–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielińska, D.; Rzepkowska, A.; Radawska, A.; Zieliński, K. In vitro screening of selected probiotic properties of Lactobacillus strains isolated from traditional fermented cabbage and cucumber. Curr. Microbiol. 2015, 70, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.H.; Tsai, C.C.; Hsih, H.Y.; Tsen, H.Y. Screening from pickled vegetables the potential probiotic strains of lactic acid bacteria able to inhibit the Salmonella invasion in mice. J. Appl. Microbiol. 2008, 104, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Nyblom, H.; Berggren, U.; Balldin, J.; Olsson, R. High AST/ALT ratio may indicate advanced alcoholic liver disease rather than heavy drinking. Alcohol Alcohol. 2004, 39, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Hjorth, M.F.; Blædel, T.; Bendtsen, L.Q.; Lorenzen, J.K.; Holm, J.B.; Kiilerich, P.; Roager, H.M.; Kristiansen, K.; Larsen, L.H.; Astrup, A. Prevotella-to-Bacteroides ratio predicts body weight and fat loss success on 24-week diets varying in macronutrient composition and dietary fiber: Results from a post-hoc analysis. Int. J. Obes. 2019, 43, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Long, W.; Zhang, C.; Liu, S.; Zhao, L.; Hamaker, B.R. Fiber-utilizing capacity varies in Prevotella- versus Bacteroides-dominated gut microbiota. Sci. Rep. 2017, 7, 2594. [Google Scholar] [CrossRef]
- Fischer, F.; Romero, R.; Hellhund, A.; Linne, U.; Bertrams, W.; Pinkenburg, O.; Eldin, H.S.; Binder, K.; Jacob, R.; Walker, A.; et al. Dietary cellulose induces anti-inflammatory immunity and transcriptional programs via maturation of the intestinal microbiota. Gut Microbes 2020, 12, 1–17. [Google Scholar] [CrossRef]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef] [Green Version]
- Mowat, A.M.; Agace, W.W. Regional specialization within the intestinal immune system. Nat. Rev. Immunol. 2014, 14, 667–685. [Google Scholar] [CrossRef]
- Ginaldi, L.; Loreto, M.F.; Corsi, M.P.; Modesti, M.; De Martinis, M. Immunosenescence and infectious diseases. Microb. Infect. 2001, 3, 851–857. [Google Scholar] [CrossRef]
- Takeda, K.; Okumura, K. Effects of a fermented milk drink containing Lactobacillus casei strain Shirota on the human NK-cell activity. J. Nutr. 2007, 137, 791S–793S. [Google Scholar] [CrossRef] [PubMed]
- Moro-García, M.A.; Alonso-Arias, R.; Baltadjieva, M.; Benítez, C.F.; Barrial, M.A.F.; Ruisánchez, E.D.; Santos, R.A.; Sánchez, M.Á.; Miján, J.S.; López-Larrea, C. Oral supplementation with Lactobacillus delbrueckii subsp. bulgaricus 8481 enhances systemic immunity in elderly subjects. Age 2013, 35, 1311–1326. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Otani, H. Stimulatory effect of lactic acid bacteria from commercially available nozawana-zuke pickle on cytokine expression by mouse spleen cells. Biosci. Biotech. Biochem. 2006, 70, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Bouladoux, N.; Hall, J.; Grainger, J.; Dos Santos, L.; Kann, M.; Nagarajan, V.; Verthelyi, D.; Belkaid, Y. Regulatory role of suppressive motifs from commensal DNA. Mucosal Immunol. 2012, 5, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Abraham, C.; Medzhitov, R. Interactions between the host innate immune system and microbes in inflammatory bowel disease. Gastroenterology 2011, 140, 1729–1737. [Google Scholar] [CrossRef] [Green Version]
- Delzenne, N.M.; Neyrinck, A.M.; Cani, P.D. Gut microbiota and metabolic disorders: How prebiotic can work? Br. J. Nutr. 2013, 109, S81–S85. [Google Scholar] [CrossRef] [Green Version]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; Rudensky, A.Y. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef]
- Martins, E.M.F.; Ramos, A.M.; Vanzela, E.S.L.; Stringheta, P.C.; de Oliveira Pinto, C.L.; Martins, J.M. Products of vegetable origin: A new alternative for the consumption of probiotic bacteria. Food Res. Int. 2013, 51, 764–770. [Google Scholar] [CrossRef]
- Yamamoto, K.; Furuya, K.; Yamada, K.; Takahashi, F.; Hamajima, C.; Tanaka, S. Enhancement of natural killer activity and IFN-γ production in an IL-12-dependent manner by a Brassica rapa L. Biosci. Biotech. Biochem. 2018, 82, 654–668. [Google Scholar] [CrossRef] [Green Version]
- Kwak, J.H.; Baek, S.H.; Woo, Y.; Han, J.K.; Kim, B.G.; Kim, O.Y.; Lee, J.H. Beneficial immunostimulatory effect of short-term Chlorella supplementation: Enhancement of natural killer cell activity and early inflammatory response (randomized, double-blinded, placebo-controlled trial). Nutr. J. 2012, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Kiecolt-Glaser, J.K.; McGuire, L.; Robles, T.F.; Glaser, R. Psychoneuroimmunology: Psychological influences on immune function and health. J. Consult. Clin. Psychol. 2002, 70, 537. [Google Scholar] [CrossRef] [PubMed]
- Aiso, I.; Inoue, H.; Seiyama, Y.; Kuwano, T. Compared with the intake of commercial vegetable juice, the intake of fresh fruit and komatsuna (Brassica rapa L. var. perviridis) juice mixture reduces serum cholesterol in middle-aged men: A randomized controlled pilot study. Lipids Health Dis. 2014, 13, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0 wk | 4 wk | p-Value | |
|---|---|---|---|
| Age (year) | 52.00 ± 1.44 | ||
| Male/female (n) | 11/9 | ||
| Weight (kg) | 72.88 ± 2.27 | 72.39 ± 2.25 | 0.0616 |
| Fat percentage (%) | 31.83 ± 1.45 | 31.80 ± 1.38 | 0.6291 |
| Systolic blood pressure (mmHg) | 133.95 ± 4.58 | 131.75 ± 4.09 | 0.3041 |
| Diastolic blood pressure (mmHg) | 83.30 ± 3.32 | 83.60 ± 3.04 | 1.0000 |
| 0 wk | 4 wk | p-Value | |
|---|---|---|---|
| Glucose (mg/dL) | 92.75 ± 5.04 | 94.4 ± 5.56 | 0.2463 |
| Total protein (g/dL) | 7.44 ± 0.08 | 7.52 ± 0.08 | 0.2852 |
| Triglycerides (mg/dL) | 124.90 ± 14.76 | 137.50 ± 13.68 | 0.3225 |
| Total cholesterol (mg/dL) | 229.25 ± 9.53 | 225.95 ± 8.45 | 0.6274 |
| HDL cholesterol (mg/dL) | 57.95 ± 3.01 | 57.50 ± 3.96 | 0.7318 |
| LDL cholesterol (mg/dL) | 144.00 ± 8.07 | 139.75 ± 6.97 | 0.2861 |
| AST (U/L) | 27.60 ± 2.76 | 23.65 ± 1.97 | 0.0088 |
| ALT (U/L) | 31.45 ± 3.96 | 26.95 ± 3.43 | 0.0259 |
| Creatinine (mg/dL) | 0.78 ± 0.04 | 0.77 ± 0.04 | 0.125 |
| Uric acid (mg/dL) | 5.98 ± 0.35 | 5.95 ± 0.36 | 0.9679 |
| 0 wk | 4 wk | p-Value | |
|---|---|---|---|
| RBCs (104/mL) | 480.65 ± 10.92 | 478.90 ± 9.65 | 0.7933 |
| Hemoglobin (g/dL) | 14.64 ± 0.35 | 14.55 ± 0.33 | 0.6044 |
| Hematocrit (%) | 43.77 ± 0.81 | 43.45 ± 0.78 | 0.5062 |
| MCV (fL) | 91.20 ± 1.01 | 90.85 ± 1.03 | 0.2527 |
| Platelets (104/mL) | 25.79 ± 1.28 | 26.23 ± 1.40 | 0.5015 |
| WBCs (103/mL) | 5.76 ± 0.33 | 5.42 ± 0.25 | 0.1256 |
| Item | Group P | Group LP | Group B |
|---|---|---|---|
| Number of subjects (Subject ID) | 4 (1, 9, 17, 21) | 5 (10, 12, 16, 22, 23) | 11 (3, 4, 6, 7, 8, 14, 18, 19, 20, 24, 25) |
| Occupation of Prevotella 1 | High (>26%) | Low (<5%) or zero | Zero |
| Signaturing RFs (Length of the fragment) 1 | Clostridium cluster IX (110) Prevotella (317) | Bacteroides (366) Clostridium subcluster XIVa (494, 940, 990) Clostridium cluster IV (749) | Bacteroides (469) Clostridium cluster XI (338) Clostridium subcluster XIVa (754) |
| Item, Period | Group P | Group LP | Group B | Contrast |
|---|---|---|---|---|
| Defecation frequency (times/day) | ||||
| −7 d to 0 d | 0.39 ± 0.12 | 0.51 ± 0.11 | 0.40 ± 0.18 | Group, p = 0.12 |
| 1 d to 14 d | 0.82 ± 0.56 | 0.83 ± 0.25 | 0.52 ± 0.27 | Period, p = 0.02 A |
| 15 d to 28 d | 0.77 ± 0.28 | 0.89 ± 0.27 | 0.68 ± 0.26 | Group × Period, p = 0.92 |
| Stool consistency 1 | ||||
| −7 d to 0 d | 3.1 ± 0.9 | 2.8 ± 0.7 | 2.5 ± 1.2 | Group, p = 0.27 |
| 1 d to 14 d | 3.9 ± 0.1 | 3.1 ± 0.9 | 3.3 ± 0.8 | Period, p = 0.21 |
| 15 d to 28 d | 3.5 ± 0.7 | 3.2 ± 1.0 | 3.6 ± 1.0 | Group × Period, p = 0.87 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, S.; Yamamoto, K.; Hamajima, C.; Takahashi, F.; Endo, K.; Uyeno, Y. Dietary Supplementation with Fermented Brassica rapa L. Stimulates Defecation Accompanying Change in Colonic Bacterial Community Structure. Nutrients 2021, 13, 1847. https://doi.org/10.3390/nu13061847
Tanaka S, Yamamoto K, Hamajima C, Takahashi F, Endo K, Uyeno Y. Dietary Supplementation with Fermented Brassica rapa L. Stimulates Defecation Accompanying Change in Colonic Bacterial Community Structure. Nutrients. 2021; 13(6):1847. https://doi.org/10.3390/nu13061847
Chicago/Turabian StyleTanaka, Sachi, Kana Yamamoto, Chisato Hamajima, Fuka Takahashi, Katsunori Endo, and Yutaka Uyeno. 2021. "Dietary Supplementation with Fermented Brassica rapa L. Stimulates Defecation Accompanying Change in Colonic Bacterial Community Structure" Nutrients 13, no. 6: 1847. https://doi.org/10.3390/nu13061847
APA StyleTanaka, S., Yamamoto, K., Hamajima, C., Takahashi, F., Endo, K., & Uyeno, Y. (2021). Dietary Supplementation with Fermented Brassica rapa L. Stimulates Defecation Accompanying Change in Colonic Bacterial Community Structure. Nutrients, 13(6), 1847. https://doi.org/10.3390/nu13061847