
The Effects of High-Protein Diet and Resistance Training on Glucose Control and Inflammatory Profile of Visceral Adipose Tissue in Rats

 , , , ,
, , , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Study Design
2.2. Resistance Training Protocol
2.3. Metabolic Analysis
2.4. Histological Analysis
2.5. Biomarkers Analysis
2.6. Statistical Analysis
3. Results
3.1. Diet Characteristics
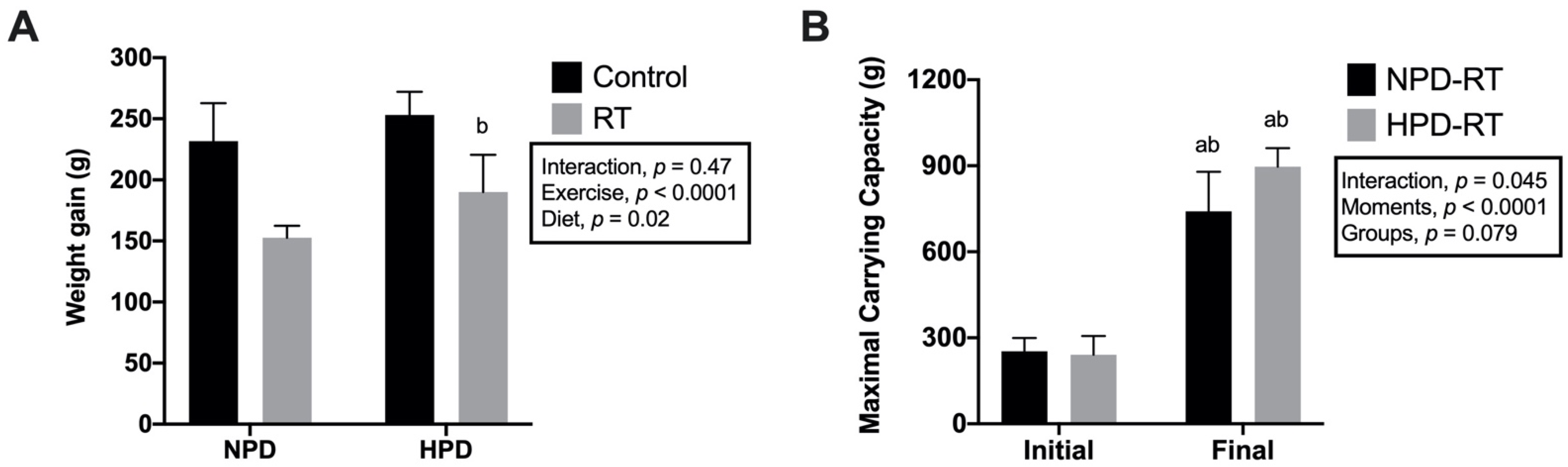
3.2. Body Weight Gain and Strength Development
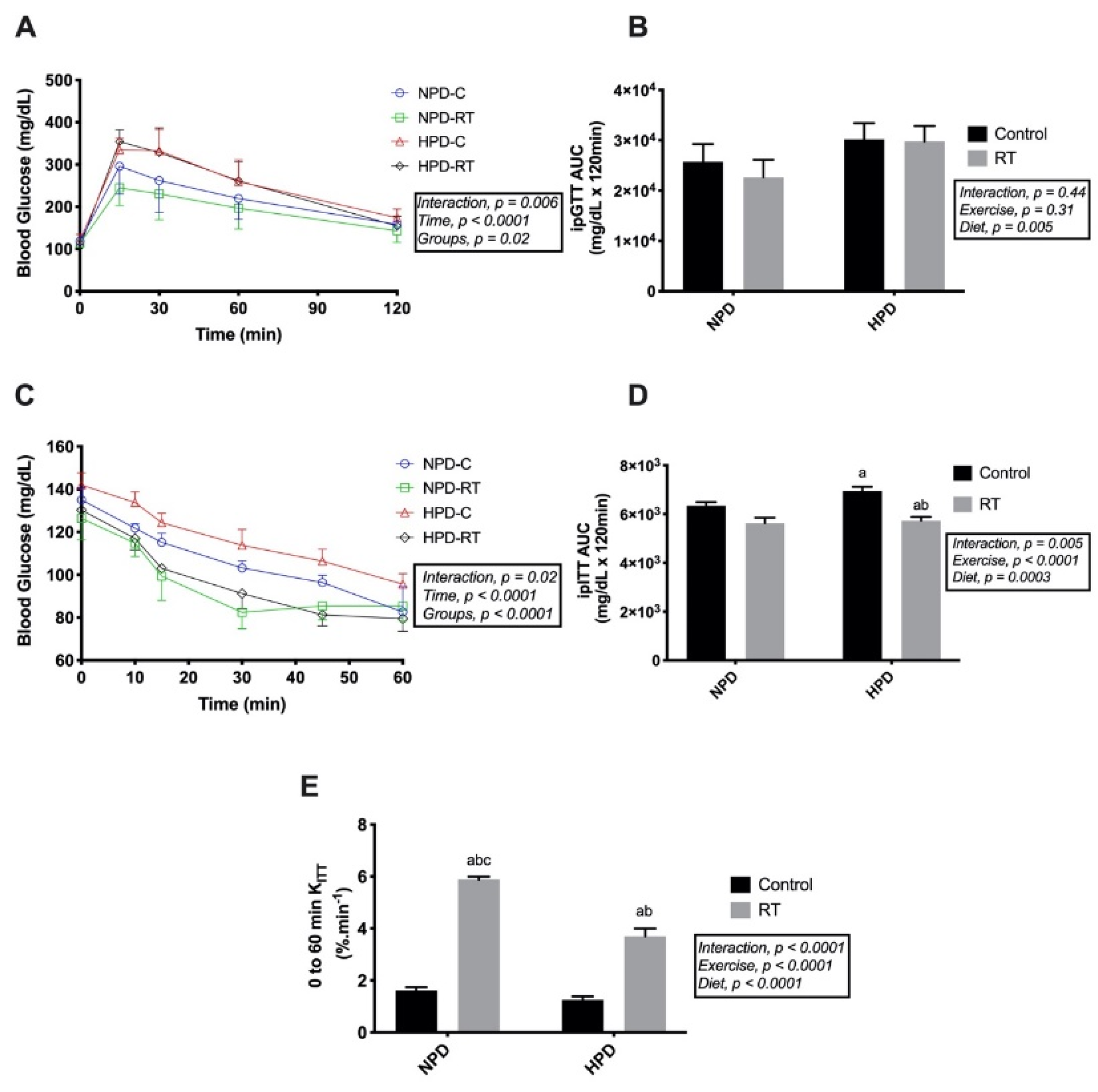
3.3. Whole-Body Glucose and Insulin Tolerance Tests
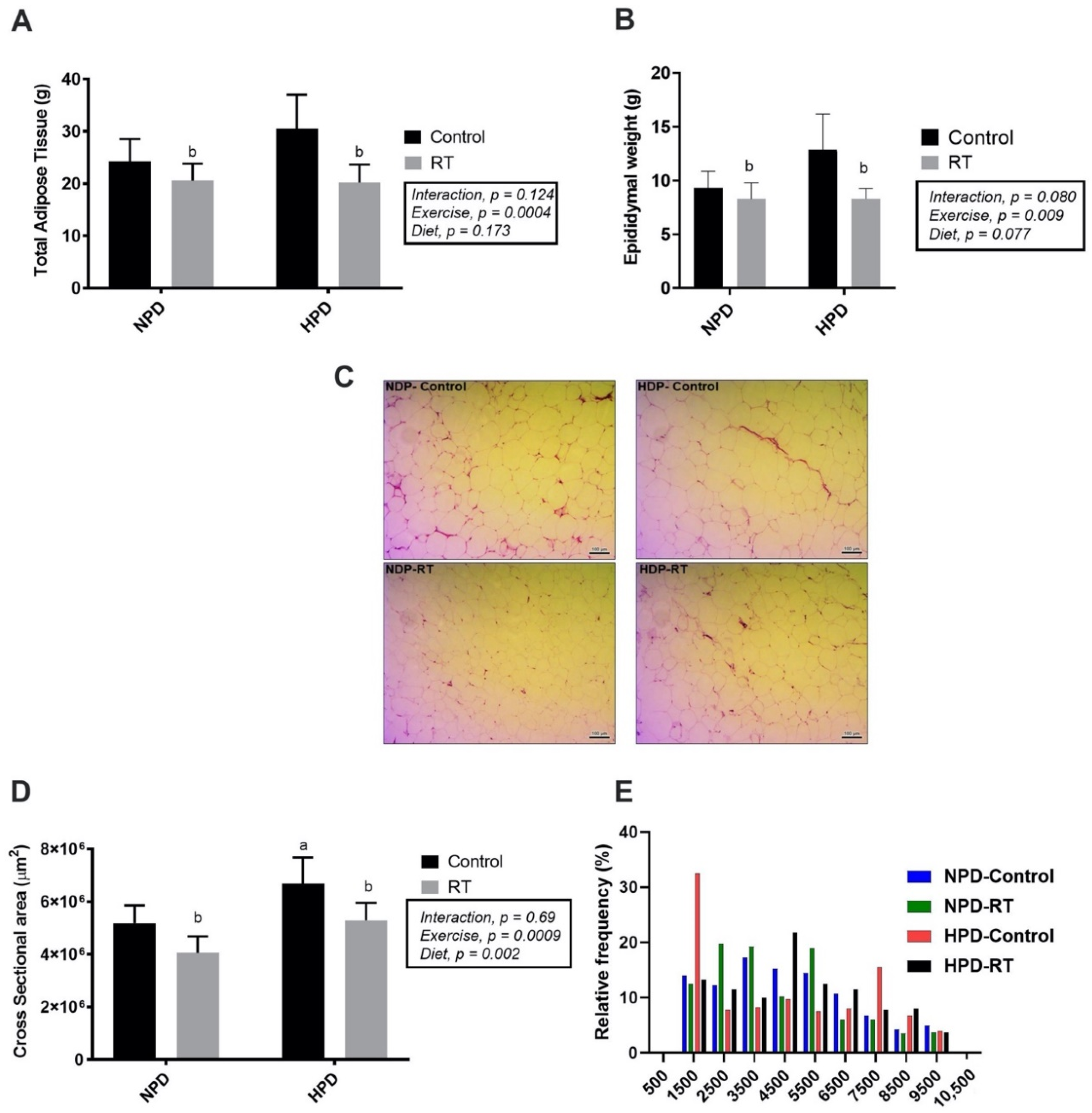
3.4. Effects of HPD Diet and RT on Adipose Tissue Weight and Adipocyte Size
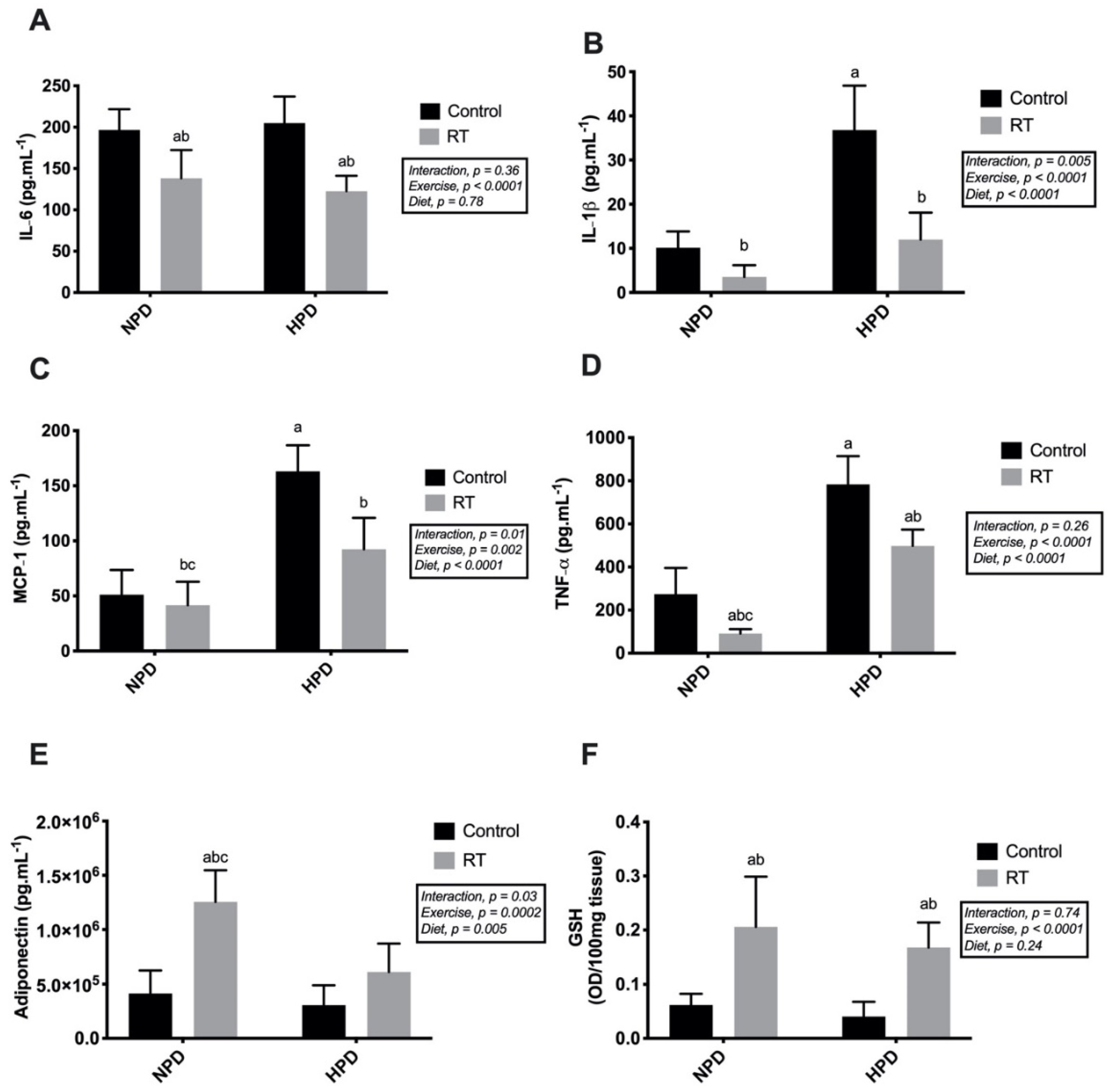
3.5. Cytokines and GSH Levels in the Visceral Adipose Tissue in Response to HPD Diet and RT
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Pesta, D.H.; Samuel, V.T. A high-protein diet for reducing body fat: Mechanisms and possible caveats. Nutr. Metab. 2014, 11, 53. [Google Scholar] [CrossRef] [Green Version]
- Ancu, O.; Mickute, M.; Guess, N.D.; Hurren, N.M.; Burd, N.A.; Mackenzie, R.W. Does high dietary protein intake contribute to the increased risk of developing prediabetes and type 2 diabetes? Appl. Physiol. Nutr. Metab. 2021, 46, 1–9. [Google Scholar] [CrossRef]
- Mitra, S.R.; Tan, P.Y. Effect of an individualised high-protein, energy-restricted diet on anthropometric and cardio-metabolic parameters in overweight and obese Malaysian adults: A 6-month randomised controlled study. Br. J. Nutr. 2019, 121, 1002–1017. [Google Scholar] [CrossRef]
- Antonio, J.; Ellerbroek, A.; Silver, T.; Orris, S.; Scheiner, M.; Gonzalez, A.; Peacock, C.A. A high protein diet (3.4 g/kg/d) combined with a heavy resistance training program improves body composition in healthy trained men and women—A follow-up investigation. J. Int. Soc. Sports Nutr. 2015, 12, 39. [Google Scholar] [CrossRef] [Green Version]
- Batista, A.; Monteiro, C.P.; Borrego, R.; Matias, C.N.; Teixeira, F.J.; Valamatos, M.J.; Oliveira, A.C.; Reis, J.F.; Mendes, L.; Sardinha, L.B. Association between whey protein, regional fat mass, and strength in resistance-trained men: A cross-sectional study. Appl. Physiol. Nutr. Metab. 2019, 44, 7–12. [Google Scholar] [CrossRef]
- Santesso, N.; Akl, E.A.; Bianchi, M.; Mente, A.; Mustafa, R.; Heels-Ansdell, D.; Schunemann, H.J. Effects of higher- versus lower-protein diets on health outcomes: A systematic review and meta-analysis. Eur. J. Clin. Nutr. 2012, 66, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Freudenberg, A.; Petzke, K.J.; Klaus, S. Comparison of high-protein diets and leucine supplementation in the prevention of metabolic syndrome and related disorders in mice. J. Nutr. Biochem. 2012, 23, 1524–1530. [Google Scholar] [CrossRef]
- Kiilerich, P.; Myrmel, L.S.; Fjaere, E.; Hao, Q.; Hugenholtz, F.; Sonne, S.B.; Derrien, M.; Pedersen, L.M.; Petersen, R.K.; Mortensen, A.; et al. Effect of a long-term high-protein diet on survival, obesity development, and gut microbiota in mice. Am. J. Physiol. Endocrinol. Metab. 2016, 310, E886–E899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Rosa Lima, T.; Avila, E.T.P.; Fraga, G.A.; de Souza Sena, M.; de Souza Dias, A.B.; de Almeida, P.C.; Dos Santos Trombeta, J.C.; Junior, R.C.V.; Damazo, A.S.; Navalta, J.W.; et al. Effect of administration of high-protein diet in rats submitted to resistance training. Eur. J. Nutr. 2018, 57, 1083–1096. [Google Scholar] [CrossRef]
- Nogueira, M.E.; Sousa Neto, I.V.; Motta-Santos, D.; Cantuaria, A.P.C.; Lima, S.M.F.; Rezende, T.M.B.; Santana, H.A.P.; Petriz, B.A.; Marqueti, R.C.; Almeida, J.A. High-protein diet associated with resistance training reduces cardiac TNF-alpha levels and up-regulates MMP-2 activity in rats. Arch. Physiol. Biochem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kurki, E.; Shi, J.; Martonen, E.; Finckenberg, P.; Mervaala, E. Distinct effects of calorie restriction on adipose tissue cytokine and angiogenesis profiles in obese and lean mice. Nutr. Metab. 2012, 9, 64. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Quintela, A.; Churruca, I.; Portillo, M.P. The role of dietary fat in adipose tissue metabolism. Public Health Nutr. 2007, 10, 1126–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiloulis, T.; Watt, M.J. Exercise and the Regulation of Adipose Tissue Metabolism. Prog. Mol. Biol. Transl. Sci. 2015, 135, 175–201. [Google Scholar] [CrossRef]
- Ziegler, A.K.; Damgaard, A.; Mackey, A.L.; Schjerling, P.; Magnusson, P.; Olesen, A.T.; Kjaer, M.; Scheele, C. An anti-inflammatory phenotype in visceral adipose tissue of old lean mice, augmented by exercise. Sci. Rep. 2019, 9, 12069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bluher, M. Adipose tissue dysfunction in obesity. Exp. Clin. Endocrinol. Diabetes 2009, 117, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Auger, C.; Jeschke, M.G. Adipose Tissue Metabolic Function and Dysfunction: Impact of Burn Injury. Front. Cell Dev. Biol. 2020, 8, 599576. [Google Scholar] [CrossRef]
- Matafome, P.; Seica, R. Function and Dysfunction of Adipose Tissue. Adv. Neurobiol. 2017, 19, 3–31. [Google Scholar] [CrossRef]
- Meln, I.; Wolff, G.; Gajek, T.; Koddebusch, J.; Lerch, S.; Harbrecht, L.; Hong, W.; Bayindir-Buchhalter, I.; Krunic, D.; Augustin, H.G.; et al. Dietary calories and lipids synergistically shape adipose tissue cellularity during postnatal growth. Mol. Metab. 2019, 24, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Vidal, P.; Stanford, K.I. Exercise-Induced Adaptations to Adipose Tissue Thermogenesis. Front. Endocrinol. 2020, 11, 270. [Google Scholar] [CrossRef] [PubMed]
- Groussard, C.; Maillard, F.; Vazeille, E.; Barnich, N.; Sirvent, P.; Otero, Y.F.; Combaret, L.; Madeuf, E.; Sourdrille, A.; Delcros, G.; et al. Tissue-Specific Oxidative Stress Modulation by Exercise: A Comparison between MICT and HIIT in an Obese Rat Model. Oxid. Med. Cell. Longev. 2019, 2019, 1965364. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, T.; Ogasawara, J.; Shirato, K.; Izawa, T.; Oh-Ishi, S.; Ishibashi, Y.; Radak, Z.; Ohno, H.; Kizaki, T. Exercise Training Attenuates the Dysregulated Expression of Adipokines and Oxidative Stress in White Adipose Tissue. Oxid. Med. Cell. Longev. 2017, 2017, 9410954. [Google Scholar] [CrossRef]
- Strasser, B.; Arvandi, M.; Siebert, U. Resistance training, visceral obesity and inflammatory response: A review of the evidence. Obes. Rev. 2012, 13, 578–591. [Google Scholar] [CrossRef] [PubMed]
- Donatto, F.F.; Neves, R.X.; Rosa, F.O.; Camargo, R.G.; Ribeiro, H.; Matos-Neto, E.M.; Seelaender, M. Resistance exercise modulates lipid plasma profile and cytokine content in the adipose tissue of tumour-bearing rats. Cytokine 2013, 61, 426–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Almeida, J.A.; Santana, H.A.P.; Motta-Santos, D.; Nogueira, M.E.; Silva, K.K.S.; Miotto, H.; Medeiros, C.S.; Faria-Ravagnani, C.C.; Voltarelli, F.A.; Guimaraes, R.C.A. High-Protein Diet Associated with Bocaiuva Supplementation Decreases Body Fat and Improves Glucose Tolerance in Resistance-Trained Rats. J. Med. Food 2020, 23, 258–265. [Google Scholar] [CrossRef]
- De Sousa Neto, I.V.; Tibana, R.A.; da Cunha Nascimento, D.; Vieira, D.C.; Durigan, J.L.; Pereira, G.B.; Navalta, J.W.; de Cassia Marqueti, R.; Prestes, J. Effects of Resistance Training Volume on MMPs in Circulation, Muscle and Adipose Tissue. Int. J. Sports Med. 2017, 38, 307–313. [Google Scholar] [CrossRef]
- De Sousa Neto, I.V.; Tibana, R.A.; da Silva, L.G.O.; de Lira, E.M.; do Prado, G.P.G.; de Almeida, J.A.; Franco, O.L.; Durigan, J.L.Q.; Adesida, A.B.; de Sousa, M.V.; et al. Paternal Resistance Training Modulates Calcaneal Tendon Proteome in the Offspring Exposed to High-Fat Diet. Front. Cell Dev. Biol. 2020, 8, 380. [Google Scholar] [CrossRef]
- De Sousa Neto, I.V.; Durigan, J.L.Q.; Guzzoni, V.; Tibana, R.A.; Prestes, J.; de Araujo, H.S.S.; Marqueti, R.C. Effects of Resistance Training on Matrix Metalloproteinase Activity in Skeletal Muscles and Blood Circulation During Aging. Front. Physiol. 2018, 9, 190. [Google Scholar] [CrossRef] [PubMed]
- Bonora, E.; Moghetti, P.; Zancanaro, C.; Cigolini, M.; Querena, M.; Cacciatori, V.; Corgnati, A.; Muggeo, M. Estimates of in vivo insulin action in man: Comparison of insulin tolerance tests with euglycemic and hyperglycemic glucose clamp studies. J. Clin. Endocrinol. Metab. 1989, 68, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, R.E.; Huber, J.S.; Frendo-Cumbo, S.; Simpson, J.A.; Wright, D.C. Adipose Tissue Insulin Action and IL-6 Signaling after Exercise in Obese Mice. Med. Sci. Sports Exerc. 2015, 47, 2034–2042. [Google Scholar] [CrossRef] [Green Version]
- Coutinho, J.V.; Freitas-Lima, L.C.; Freitas, F.F.; Freitas, F.P.; Podratz, P.L.; Magnago, R.P.; Porto, M.L.; Meyrelles, S.S.; Vasquez, E.C.; Brandao, P.A.; et al. Tributyltin chloride induces renal dysfunction by inflammation and oxidative stress in female rats. Toxicol. Lett. 2016, 260, 52–69. [Google Scholar] [CrossRef]
- Newgard, C.B. Interplay between lipids and branched-chain amino acids in development of insulin resistance. Cell Metab. 2012, 15, 606–614. [Google Scholar] [CrossRef] [Green Version]
- Rietman, A.; Schwarz, J.; Tome, D.; Kok, F.J.; Mensink, M. High dietary protein intake, reducing or eliciting insulin resistance? Eur. J. Clin. Nutr. 2014, 68, 973–979. [Google Scholar] [CrossRef] [Green Version]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose Tissue Dysfunction as Determinant of Obesity-Associated Metabolic Complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [Green Version]
- Naufahu, J.; Elliott, B.; Markiv, A.; Dunning-Foreman, P.; McGrady, M.; Howard, D.; Watt, P.; Mackenzie, R.W.A. High-Intensity Exercise Decreases IP6K1 Muscle Content and Improves Insulin Sensitivity (SI2*) in Glucose-Intolerant Individuals. J. Clin. Endocrinol. Metab. 2018, 103, 1479–1490. [Google Scholar] [CrossRef] [PubMed]
- Burd, N.A.; West, D.W.; Moore, D.R.; Atherton, P.J.; Staples, A.W.; Prior, T.; Tang, J.E.; Rennie, M.J.; Baker, S.K.; Phillips, S.M. Enhanced amino acid sensitivity of myofibrillar protein synthesis persists for up to 24 h after resistance exercise in young men. J. Nutr. 2011, 141, 568–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aragon, A.A.; Schoenfeld, B.J.; Wildman, R.; Kleiner, S.; VanDusseldorp, T.; Taylor, L.; Earnest, C.P.; Arciero, P.J.; Wilborn, C.; Kalman, D.S.; et al. International society of sports nutrition position stand: Diets and body composition. J. Int. Soc. Sports Nutr. 2017, 14, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morato, P.N.; Lollo, P.C.; Moura, C.S.; Batista, T.M.; Camargo, R.L.; Carneiro, E.M.; Amaya-Farfan, J. Whey protein hydrolysate increases translocation of GLUT-4 to the plasma membrane independent of insulin in wistar rats. PLoS ONE 2013, 8, e71134. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, M.B.T.; Guzzoni, V.; Hord, J.M.; Lopes, G.N.; Marqueti, R.C.; de Andrade, R.V.; Selistre-de-Araujo, H.S.; Durigan, J.L.Q. Resistance training regulates gene expression of molecules associated with intramyocellular lipids, glucose signaling and fiber size in old rats. Sci. Rep. 2017, 7, 8593. [Google Scholar] [CrossRef] [PubMed]
- Rohling, M.; Herder, C.; Stemper, T.; Mussig, K. Influence of Acute and Chronic Exercise on Glucose Uptake. J. Diabetes Res. 2016, 2016, 2868652. [Google Scholar] [CrossRef] [Green Version]
- Yaspelkis, B.B., 3rd; Singh, M.K.; Trevino, B.; Krisan, A.D.; Collins, D.E. Resistance training increases glucose uptake and transport in rat skeletal muscle. Acta Physiol. Scand. 2002, 175, 315–323. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [Green Version]
- Lihn, A.S.; Pedersen, S.B.; Richelsen, B. Adiponectin: Action, regulation and association to insulin sensitivity. Obes. Rev. 2005, 6, 13–21. [Google Scholar] [CrossRef]
- McLaughlin, T.; Lamendola, C.; Coglan, N.; Liu, T.C.; Lerner, K.; Sherman, A.; Cushman, S.W. Subcutaneous adipose cell size and distribution: Relationship to insulin resistance and body fat. Obesity 2014, 22, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.A.; Jackman, M.R.; Brown, I.L.; Johnson, G.C.; Steig, A.; Wyatt, H.R.; Hill, J.O.; Maclean, P.S. Resistant starch and exercise independently attenuate weight regain on a high fat diet in a rat model of obesity. Nutr. Metab. 2011, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Ibanez, C.A.; Vazquez-Martinez, M.; Leon-Contreras, J.C.; Reyes-Castro, L.A.; Rodriguez-Gonzalez, G.L.; Bautista, C.J.; Nathanielsz, P.W.; Zambrano, E. Different Statistical Approaches to Characterization of Adipocyte Size in Offspring of Obese Rats: Effects of Maternal or Offspring Exercise Intervention. Front. Physiol. 2018, 9, 1571. [Google Scholar] [CrossRef] [Green Version]
- Skurk, T.; Alberti-Huber, C.; Herder, C.; Hauner, H. Relationship between adipocyte size and adipokine expression and secretion. J. Clin. Endocrinol. Metab. 2007, 92, 1023–1033. [Google Scholar] [CrossRef]
- Stenkula, K.G.; Erlanson-Albertsson, C. Adipose cell size: Importance in health and disease. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R284–R295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lira, F.S.; Rosa, J.C.; Pimentel, G.D.; Tarini, V.A.; Arida, R.M.; Faloppa, F.; Alves, E.S.; do Nascimento, C.O.; Oyama, L.M.; Seelaender, M.; et al. Inflammation and adipose tissue: Effects of progressive load training in rats. Lipids Health Dis. 2010, 9, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.S.; White, A.; Perry, R.J.; Camporez, J.P.; Hidalgo, J.; Shulman, G.I.; Davis, R.J. Regulation of adipose tissue inflammation by interleukin 6. Proc. Natl. Acad. Sci. USA 2020, 117, 2751–2760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jankovic, A.; Korac, A.; Buzadzic, B.; Otasevic, V.; Stancic, A.; Daiber, A.; Korac, B. Redox implications in adipose tissue (dys)function--A new look at old acquaintances. Redox Biol. 2015, 6, 19–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.S.; Lira, F.S.; Rosa, J.C.; Paulino, E.C.; Brum, P.C.; Negrao, C.E.; dos Santos, R.V.; Batista, M.L., Jr.; do Nascimento, C.O.; Oyama, L.M.; et al. Depot-specific modulation of adipokine levels in rat adipose tissue by diet-induced obesity: The effect of aerobic training and energy restriction. Cytokine 2010, 52, 168–174. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet Outcomes | NPD-C | HPD-C | NPD-RT | HPD-RT |
|---|---|---|---|---|
| Daily food intake (g) | 20.1 ±0.4 | 20.7 ±0.1 | 19.5 ±0.5 b | 18.6 ±1.0 a,b |
| Dietary metabolizable energy (kcal/day) | 65.5 ±1.4 | 68.6 ±0.4 a | 63.3 ±1.3 b | 61.2 ±3.2 a,b |
| Feed efficiency (%) | 16.5 | 16.3 | 14.6 | 14.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medeiros, C.S.; de Sousa Neto, I.V.; Silva, K.K.S.; Cantuária, A.P.C.; Rezende, T.M.B.; Franco, O.L.; de Cassia Marqueti, R.; Freitas-Lima, L.C.; Araujo, R.C.; Yildirim, A.; et al. The Effects of High-Protein Diet and Resistance Training on Glucose Control and Inflammatory Profile of Visceral Adipose Tissue in Rats. Nutrients 2021, 13, 1969. https://doi.org/10.3390/nu13061969
Medeiros CS, de Sousa Neto IV, Silva KKS, Cantuária APC, Rezende TMB, Franco OL, de Cassia Marqueti R, Freitas-Lima LC, Araujo RC, Yildirim A, et al. The Effects of High-Protein Diet and Resistance Training on Glucose Control and Inflammatory Profile of Visceral Adipose Tissue in Rats. Nutrients. 2021; 13(6):1969. https://doi.org/10.3390/nu13061969
Chicago/Turabian StyleMedeiros, Claudia Stela, Ivo Vieira de Sousa Neto, Keemilyn Karla Santos Silva, Ana Paula Castro Cantuária, Taia Maria Berto Rezende, Octávio Luiz Franco, Rita de Cassia Marqueti, Leandro Ceotto Freitas-Lima, Ronaldo Carvalho Araujo, Azize Yildirim, and et al. 2021. "The Effects of High-Protein Diet and Resistance Training on Glucose Control and Inflammatory Profile of Visceral Adipose Tissue in Rats" Nutrients 13, no. 6: 1969. https://doi.org/10.3390/nu13061969
APA StyleMedeiros, C. S., de Sousa Neto, I. V., Silva, K. K. S., Cantuária, A. P. C., Rezende, T. M. B., Franco, O. L., de Cassia Marqueti, R., Freitas-Lima, L. C., Araujo, R. C., Yildirim, A., Mackenzie, R., & Alves Almeida, J. (2021). The Effects of High-Protein Diet and Resistance Training on Glucose Control and Inflammatory Profile of Visceral Adipose Tissue in Rats. Nutrients, 13(6), 1969. https://doi.org/10.3390/nu13061969