
Trehalose Reduces Nerve Injury Induced Nociception in Mice but Negatively Affects Alertness

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and Trehalose Treatment
2.2. Spared Nerve Injury (SNI)
2.3. Analysis of Nociception
2.4. Primary Neuron Culture
2.5. Calcium Imaging
2.6. Phenomaster Ananlysis of Voluntary Wheel Running, Feeding, and Drinking
2.7. Motor Functions
2.8. Open Field Test
2.9. Elevated Plus Maze
2.10. Barnes Maze
2.11. Sociability and Social Memory
2.12. IntelliCage Tasks and Schedule
2.13. Statistics
3. Results
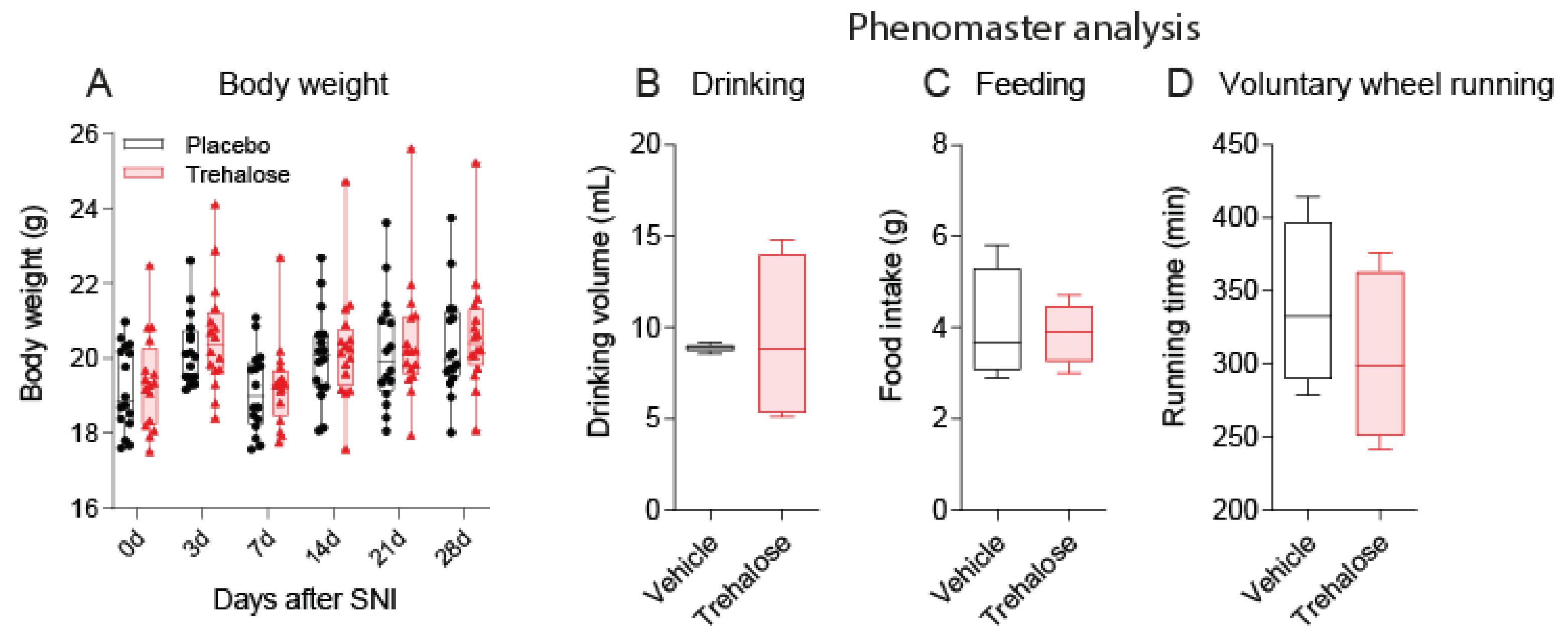
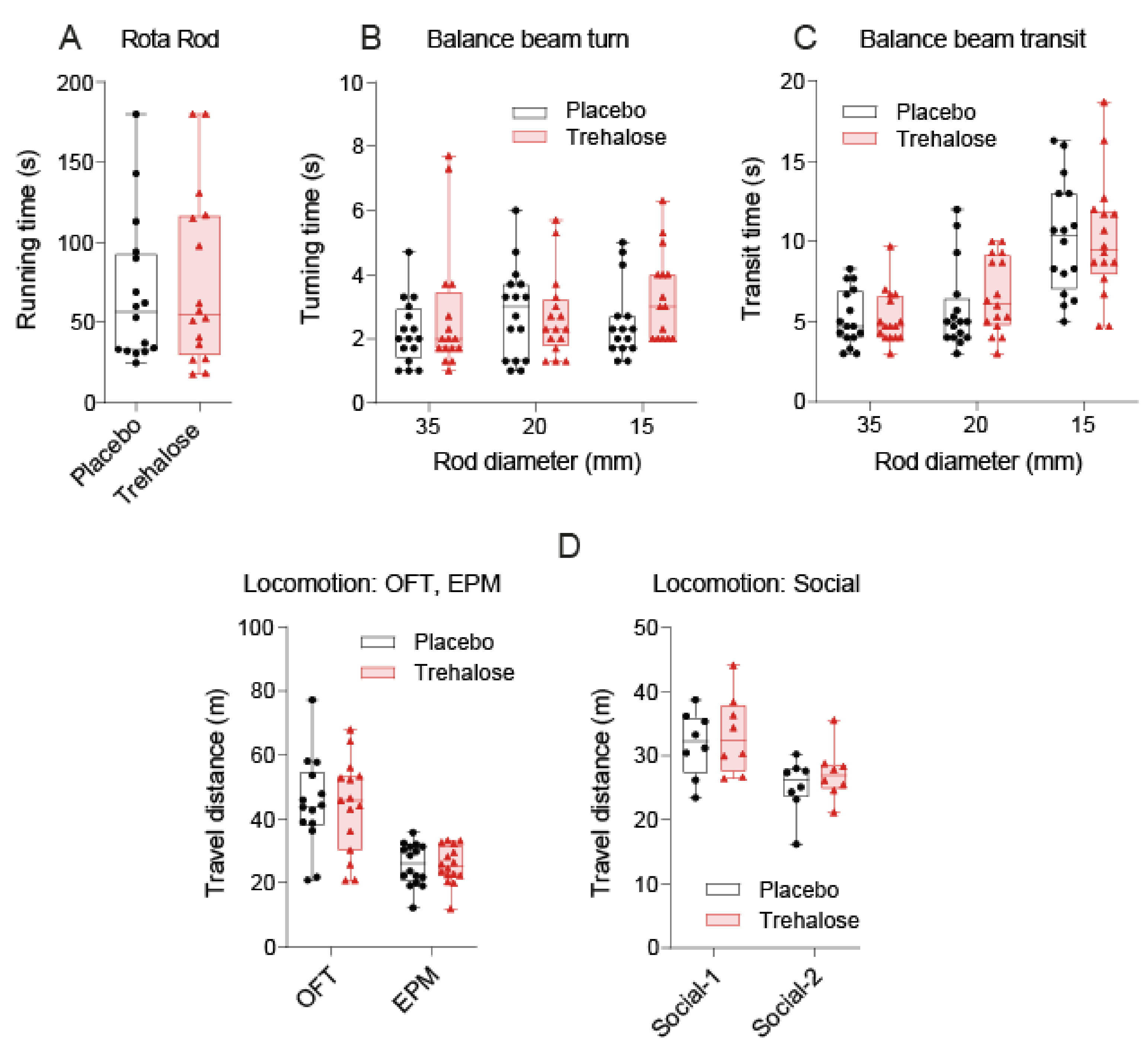
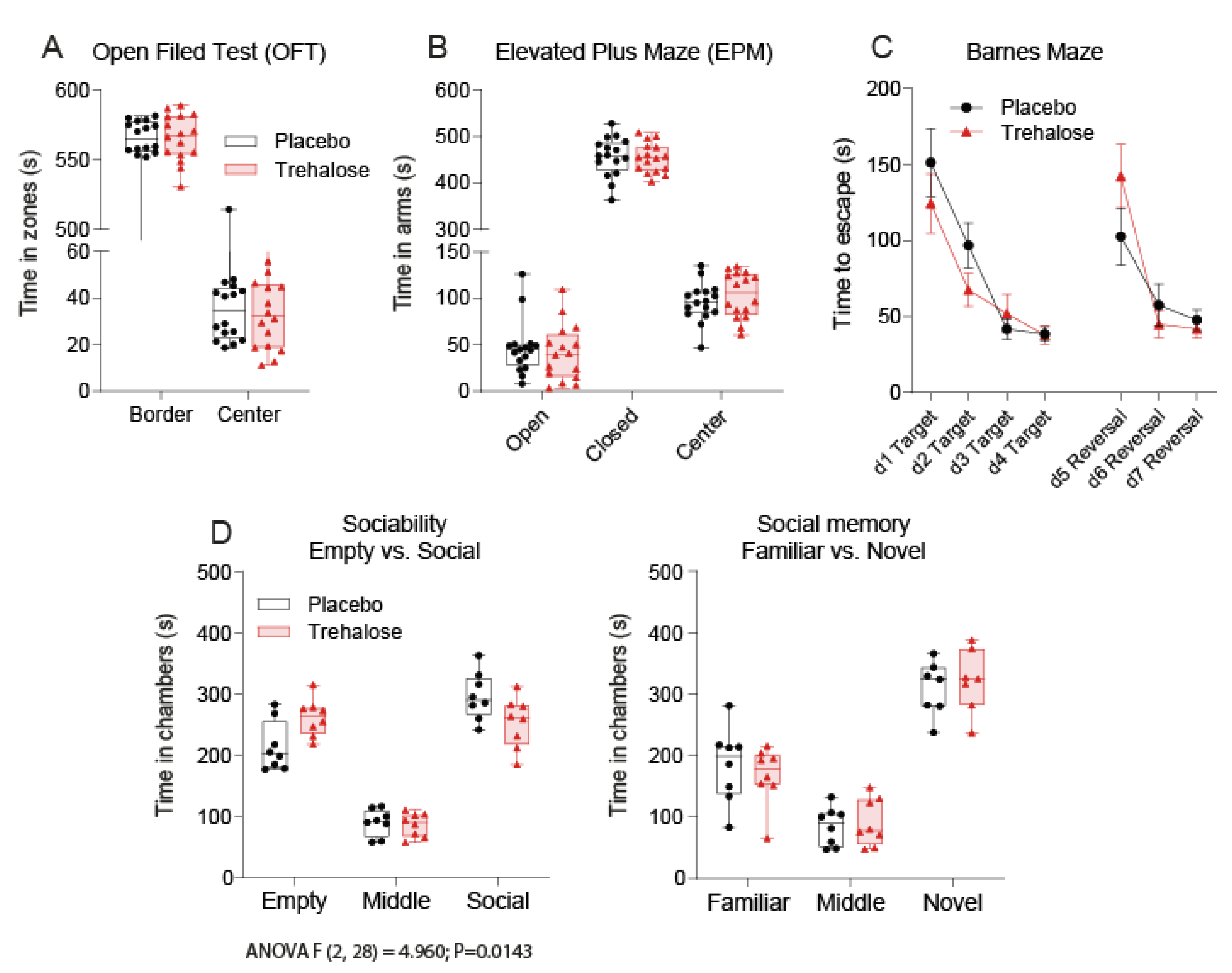
3.1. Trehalose Long-Term Treatment Is Well Tolerated
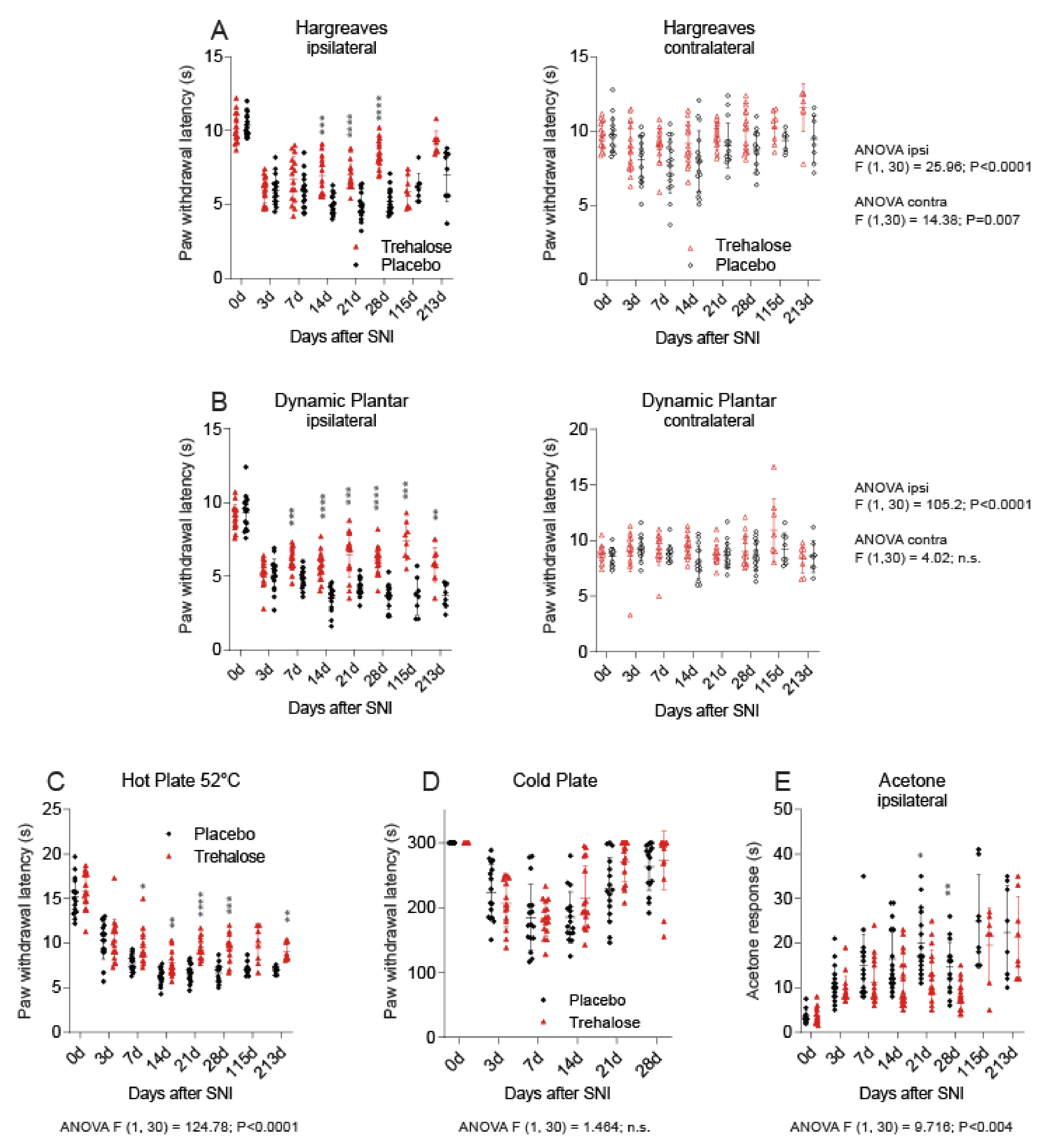
3.2. Trehalose Reduces Nociception in the SNI Model of Neuropathic Pain
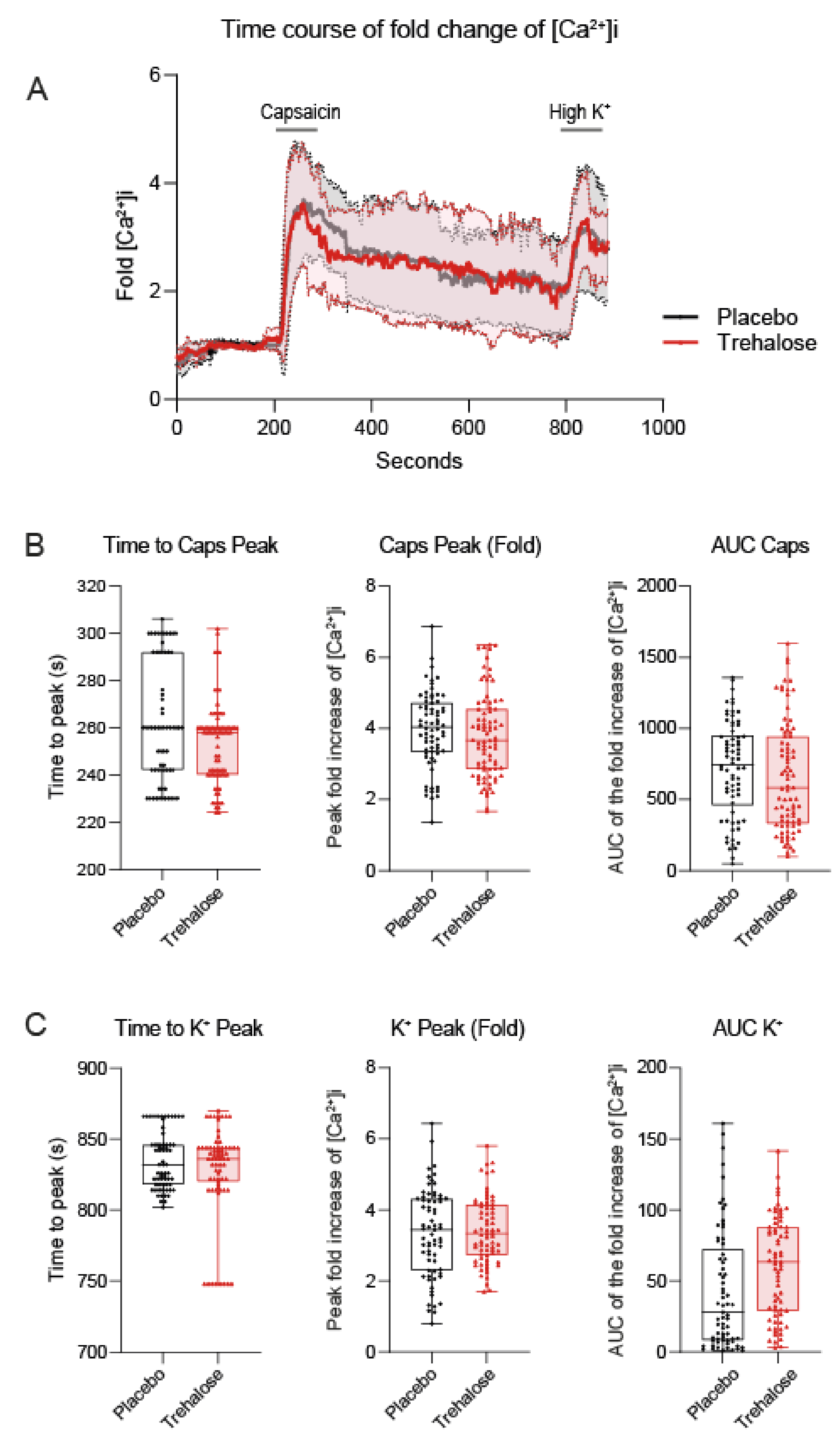
3.3. Trehalose Has No Effect on Calcium Fluxes in Primary Sensory Neurons
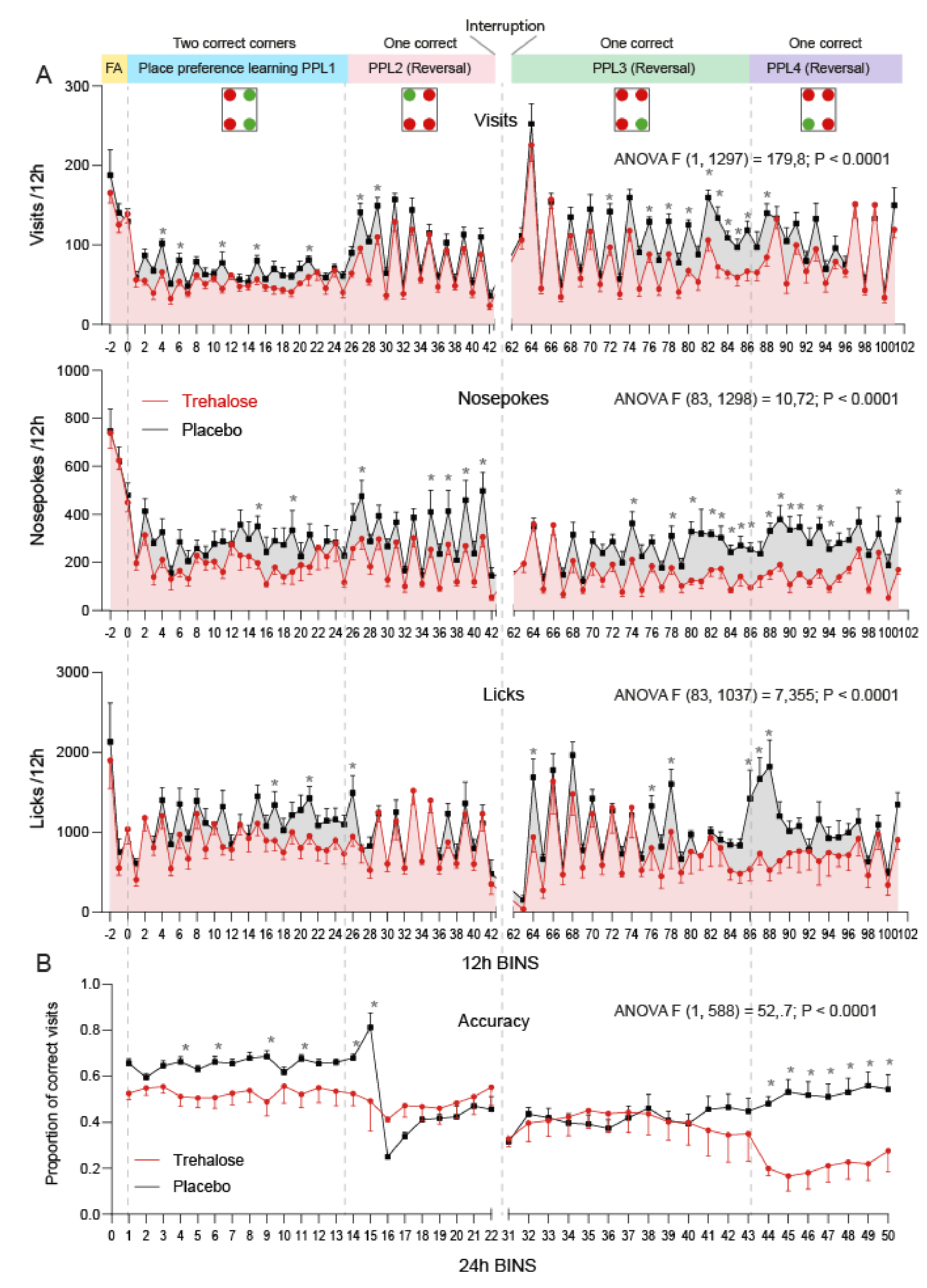
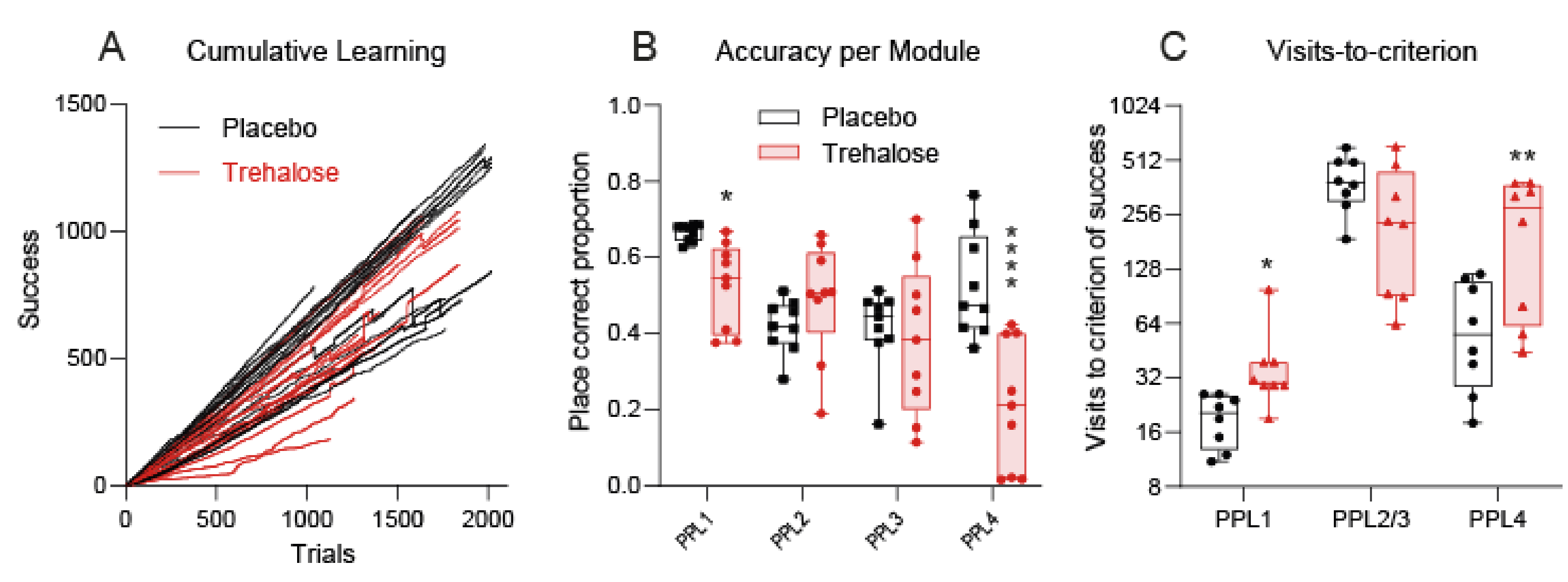
3.4. Trehalose Reduces Alertness and Accuracy in IntelliCage Learning Tasks
3.5. Trehalose Reduces Social Interest
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jain, N.K.; Roy, I. Effect of trehalose on protein structure. Protein Sci. 2008, 18, 24–36. [Google Scholar] [CrossRef]
- Halbe, L.; Rami, A. Trehalase localization in the cerebral cortex, hippocampus and cerebellum of mouse brains. J. Adv. Res. 2019, 18, 71–79. [Google Scholar] [CrossRef]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef]
- Arai, C.; Suyama, A.; Arai, S.; Arai, N.; Yoshizane, C.; Koya-Miyata, S.; Mizote, A.; Endo, S.; Ariyasu, T.; Mitsuzumi, H.; et al. Trehalose itself plays a critical role on lipid metabolism: Trehalose increases jejunum cytoplasmic lipid droplets which negatively correlated with mesenteric adipocyte size in both HFD-fed trehalase KO and WT mice. Nutr. Metab. 2020, 17, 12–22. [Google Scholar] [CrossRef]
- Van Ende, M.; Timmermans, B.; Vanreppelen, G.; Siscar-Lewin, S.; Fischer, D.; Wijnants, S.; Romero, C.L.; Yazdani, S.; Rogiers, O.; Demuyser, L.; et al. The involvement of the Candida glabrata trehalase enzymes in stress resistance and gut colonization. Virulence 2021, 12, 329–345. [Google Scholar] [CrossRef]
- Buts, J.P.; Stilmant, C.; Bernasconi, P.; Neirinck, C.; De Keyser, N. Characterization of alpha, alpha-trehalase released in the intestinal lumen by the probiotic saccharomyces boulardii. Scand. J. Gastroenterol. 2008, 43, 1489–1496. [Google Scholar] [CrossRef]
- Jiao, H.; Zhang, L.; Xie, H.-W.; Simmons, N.B.; Liu, H.; Zhao, H. Trehalase Gene as a Molecular Signature of Dietary Diversification in Mammals. Mol. Biol. Evol. 2019, 36, 2171–2183. [Google Scholar] [CrossRef] [Green Version]
- Dhaene, S.; Van Der Eycken, J.; Beerens, K.; Franceus, J.; Desmet, T.; Caroen, J. Synthesis, trehalase hydrolytic resistance and inhibition properties of 4- and 6-substituted trehalose derivatives. J. Enzym. Inhib. Med. Chem. 2020, 35, 1964–1989. [Google Scholar] [CrossRef]
- Zhang, Y.; DeBosch, B.J. Microbial and metabolic impacts of trehalose and trehalose analogues. Gut Microbes 2020, 11, 1475–1482. [Google Scholar] [CrossRef]
- Assoni, G.; Frapporti, G.; Colombo, E.; Gornati, D.; Perez-Carrion, M.D.; Polito, L.; Seneci, P.; Piccoli, G.; Arosio, D. Trehalose-based neuroprotective autophagy inducers. Bioorganic Med. Chem. Lett. 2021, 40, 127929. [Google Scholar] [CrossRef]
- Zhang, Y.; Shaikh, N.; Ferey, J.L.; Wankhade, U.D.; Chintapalli, S.V.; Higgins, C.B.; Crowley, J.R.; Heitmeier, M.R.; Stothard, A.I.; Mihi, B.; et al. Lactotrehalose, an Analog of Trehalose, Increases Energy Metabolism Without Promoting Clostridioides difficile Infection in Mice. Gastroenterology 2020, 158, 1402–1416.e2. [Google Scholar] [CrossRef]
- Evans, T.D.; Jeong, S.-J.; Zhang, X.; Sergin, I.; Razani, B. TFEB and trehalose drive the macrophage autophagy-lysosome system to protect against atherosclerosis. Autophagy 2018, 14, 724–726. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Chigurupati, S.; Raymick, J.; Mann, D.; Bowyer, J.F.; Schmitt, T.; Beger, R.D.; Hanig, J.P.; Schmued, L.C.; Paule, M.G. Neuroprotective effect of the chemical chaperone, trehalose in a chronic mptp-induced parkinson’s disease mouse model. Neurotoxicology 2014, 44, 250–262. [Google Scholar] [CrossRef]
- Liu, Z.; Chen, D.; Chen, X.; Bian, F.; Qin, W.; Gao, N.; Xiao, Y.; Li, J.; Pflugfelder, S.C.; Li, D.-Q. Trehalose Induces Autophagy Against Inflammation by Activating TFEB Signaling Pathway in Human Corneal Epithelial Cells Exposed to Hyperosmotic Stress. Investig. Opthalmology Vis. Sci. 2020, 61, 26. [Google Scholar] [CrossRef] [PubMed]
- DeBosch, B.J.; Heitmeier, M.R.; Mayer, A.; Higgins, C.B.; Crowley, J.R.; Kraft, T.; Chi, M.; Newberry, E.P.; Chen, Z.; Finck, B.N.; et al. Trehalose inhibits solute carrier 2A (SLC2A) proteins to induce autophagy and prevent hepatic steatosis. Sci. Signal. 2016, 9, ra21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, S.; Davies, J.E.; Huang, Z.; Tunnacliffe, A.; Rubinsztein, D.C. Trehalose, a novel mtor-independent autophagy enhancer, accelerates the clearance of mutant huntingtin and alpha-synuclein. J. Biol. Chem. 2007, 282, 5641–5652. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Chen, S.; Song, L.; Tang, Y.; Shen, Y.; Jia, L.; Le, W. MTOR-independent, autophagic enhancer trehalose prolongs motor neuron survival and ameliorates the autophagic flux defect in a mouse model of amyotrophic lateral sclerosis. Autophagy 2014, 10, 588–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rusmini, P.; Cortese, K.; Crippa, V.; Cristofani, R.; Cicardi, M.E.; Ferrari, V.; Vezzoli, G.; Tedesco, B.; Meroni, M.; Messi, E.; et al. Trehalose induces autophagy via lysosomal-mediated TFEB activation in models of motoneuron degeneration. Autophagy 2019, 15, 631–651. [Google Scholar] [CrossRef]
- Abokyi, S.; Shan, S.W.; To, C.-H.; Chan, H.H.; Tse, D.Y.-Y. Autophagy Upregulation by the TFEB Inducer Trehalose Protects against Oxidative Damage and Cell Death Associated with NRF2 Inhibition in Human RPE Cells. Oxidative Med. Cell. Longev. 2020, 2020, 1–18. [Google Scholar] [CrossRef]
- Zhu, L.; Yuan, Y.; Yuan, L.; Li, L.; Liu, F.; Liu, J.; Chen, Y.; Lu, Y.; Cheng, J. Activation of TFEB-mediated autophagy by trehalose attenuates mitochondrial dysfunction in cisplatin-induced acute kidney injury. Theranostics 2020, 10, 5829–5844. [Google Scholar] [CrossRef]
- Jeong, S.J.; Stitham, J.; Evans, T.D.; Zhang, X.; Rodriguez-Velez, A.; Yeh, Y.S.; Tao, J.; Takabatake, K.; Epelman, S.; Lodhi, I.J.; et al. Trehalose causes low-grade lysosomal stress to activate tfeb and the autophagy-lysosome biogenesis response. Autophagy 2021, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.-J.; Ringeling, F.R.; Vissers, C.; Jacob, F.; Pokrass, M.; Jimenez-Cyrus, D.; Su, Y.; Kim, N.-S.; Zhu, Y.; Zheng, L.; et al. Temporal Control of Mammalian Cortical Neurogenesis by m6A Methylation. Cell 2017, 171, 877–889.e17. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Barkhordarian, H.; Emadi, S.; Park, C.B.; Sierks, M.R. Trehalose differentially inhibits aggregation and neurotoxicity of beta-amyloid 40 and 42. Neurobiol. Dis. 2005, 20, 74–81. [Google Scholar] [CrossRef]
- Izmitli, A.; Schebor, C.; McGovern, M.P.; Reddy, A.S.; Abbott, N.L.; de Pablo, J.J. Effect of trehalose on the interaction of alzheimer’s aβ-peptide and anionic lipid monolayers. Biochim. Biophys. Acta 2011, 1808, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Liang, Y.; Xu, F.; Sun, B.; Wang, Z. Trehalose rescues alzheimer’s disease phenotypes in app/ps1 transgenic mice. J. Pharm. Pharmacol. 2013, 65, 1753–1756. [Google Scholar] [CrossRef]
- Portbury, S.D.; Hare, D.J.; Sgambelloni, C.; Perronnes, K.; Portbury, A.J.; Finkelstein, D.I.; Adlard, P.A. Trehalose Improves Cognition in the Transgenic Tg2576 Mouse Model of Alzheimer’s Disease. J. Alzheimer Dis. 2017, 60, 549–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, J.; Hsiung, G.-Y.R.; Song, W. Trehalose Inhibits Aβ Generation and Plaque Formation in Alzheimer’s Disease. Mol. Neurobiol. 2020, 57, 3150–3157. [Google Scholar] [CrossRef]
- Tanaka, M.; Machida, Y.; Niu, S.; Ikeda, T.; Jana, N.; Doi, H.; Kurosawa, M.; Nekooki, M.; Nukina, N. Trehalose alleviates polyglutamine-mediated pathology in a mouse model of Huntington disease. Nat. Med. 2004, 10, 148–154. [Google Scholar] [CrossRef]
- Castillo, K.; Nassif, M.; Valenzuela, V.; Rojas, F.; Matus, S.; Mercado, G.; Court, F.A.; Van Zundert, B.; Hetz, C. Trehalose delays the progression of amyotrophic lateral sclerosis by enhancing autophagy in motoneurons. Autophagy 2013, 9, 1308–1320. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, F.-T.; Wang, Y.-X.; Guan, R.-Y.; Chen, C.; Li, D.-K.; Bu, L.-L.; Song, J.; Yang, Y.-J.; Dong, Y.; et al. Autophagic Modulation by Trehalose Reduces Accumulation of TDP-43 in a Cell Model of Amyotrophic Lateral Sclerosis via TFEB Activation. Neurotox. Res. 2018, 34, 109–120. [Google Scholar] [CrossRef]
- Portbury, S.D.; Hare, D.; Finkelstein, D.; Adlard, P.A. Trehalose improves traumatic brain injury-induced cognitive impairment. PLoS ONE 2017, 12, e0183683. [Google Scholar] [CrossRef] [PubMed]
- Portbury, S.D.; Hare, D.J.; Bishop, D.P.; Finkelstein, D.; Doble, P.A.; Adlard, P.A. Trehalose elevates brain zinc levels following controlled cortical impact in a mouse model of traumatic brain injury. Metallomics 2018, 10, 846–853. [Google Scholar] [CrossRef] [Green Version]
- Settembre, C.; Zoncu, R.; Medina, D.L.; Vetrini, F.; Erdin, S.; Erdin, S.; Huynh, T.; Ferron, M.; Karsenty, G.; Vellard, M.C.; et al. A lysosome-to-nucleus signalling mechanism senses and regulates the lysosome via mTOR and TFEB. EMBO J. 2012, 31, 1095–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Settembre, C.; Medina, D.L. TFEB and the CLEAR network. Methods Cell Biol. 2015, 126, 45–62. [Google Scholar] [CrossRef]
- Holler, C.J.; Taylor, G.; McEachin, Z.T.; Deng, Q.; Watkins, W.J.; Hudson, K.; Easley, C.A.; Hu, W.T.; Hales, C.M.; Rossoll, W.; et al. Trehalose upregulates progranulin expression in human and mouse models of GRN haploinsufficiency: A novel therapeutic lead to treat frontotemporal dementia. Mol. Neurodegener. 2016, 11, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, Y.; Chambers, J.K.; Matsuwaki, T.; Yamanouchi, K.; Nishihara, M. Possible involvement of lysosomal dysfunction in pathological changes of the brain in aged progranulin-deficient mice. Acta Neuropathol. Commun. 2014, 2, 78. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Srinivasan, K.; Friedman, B.A.; Suto, E.; Modrusan, Z.; Lee, W.P.; Kaminker, J.S.; Hansen, D.V.; Sheng, M. Progranulin deficiency causes impairment of autophagy and TDP-43 accumulation. J. Exp. Med. 2017, 214, 2611–2628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elia, L.P.; Mason, A.R.; Alijagic, A.; Finkbeiner, S. Genetic Regulation of Neuronal Progranulin Reveals a Critical Role for the Autophagy-Lysosome Pathway. J. Neurosci. 2019, 39, 3332–3344. [Google Scholar] [CrossRef] [Green Version]
- Altmann, C.; Hardt, S.; Fischer, C.; Heidler, J.; Lim, H.-Y.; Häussler, A.; Albuquerque, B.; Zimmer, B.; Möser, C.; Behrends, C.; et al. Progranulin overexpression in sensory neurons attenuates neuropathic pain in mice: Role of autophagy. Neurobiol. Dis. 2016, 96, 294–311. [Google Scholar] [CrossRef]
- Altmann, C.; Vasic, V.; Hardt, S.; Heidler, J.; Häussler, A.; Wittig, I.; Schmidt, M.H.H.; Tegeder, I. Progranulin promotes peripheral nerve regeneration and reinnervation: Role of notch signaling. Mol. Neurodegener. 2016, 11, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Kanngiesser, M.; Mair, N.; Lim, H.-Y.; Zschiebsch, K.; Blees, J.; Häussler, A.; Brüne, B.; Ferreiròs, N.; Kress, M.; Tegeder, I. Hypoxia-Inducible Factor 1 Regulates Heat and Cold Pain Sensitivity and Persistence. Antioxid. Redox Signal. 2014, 20, 2555–2571. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.; Endle, H.; Schumann, L.; Wilken-Schmitz, A.; Kaiser, J.; Gerber, S.; Vogelaar, C.F.; Schmidt, M.H.H.; Nitsch, R.; Snodgrass, I.; et al. Prevention of age-associated neuronal hyperexcitability with improved learning and attention upon knockout or antagonism of LPAR2. Cell. Mol. Life Sci. 2021, 78, 1029–1050. [Google Scholar] [CrossRef] [PubMed]
- Vogel, A.; Wilken-Schmitz, A.; Hummel, R.; Lang, M.; Gurke, R.; Schreiber, Y.; Schäfer, M.K.E.; Tegeder, I. Low brain endocannabinoids associated with persistent non-goal directed nighttime hyperactivity after traumatic brain injury in mice. Sci. Rep. 2020, 10, 1–22. [Google Scholar] [CrossRef] [PubMed]
- De Bruin, N.; Schmitz, K.; Schiffmann, S.; Tafferner, N.; Schmidt, M.; Jordan, H.; Häußler, A.; Tegeder, I.; Geisslinger, G.; Parnham, M. Multiple rodent models and behavioral measures reveal unexpected responses to FTY720 and DMF in experimental autoimmune encephalomyelitis. Behav. Brain Res. 2016, 300, 160–174. [Google Scholar] [CrossRef]
- Hempenstall, K.; Nurmikko, T.J.; Johnson, R.W.; A’Hern, R.P.; Rice, A.S. Analgesic Therapy in Postherpetic Neuralgia: A Quantitative Systematic Review. PLoS Med. 2005, 2, e164. [Google Scholar] [CrossRef] [Green Version]
- Hardt, S.; Heidler, J.; Albuquerque, B.; Valek, L.; Altmann, C.; Wilken-Schmitz, A.; Schäfer, M.K.; Wittig, I.; Tegeder, I. Loss of synaptic zinc transport in progranulin deficient mice may contribute to progranulin-associated psychopathology and chronic pain. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2017, 1863, 2727–2745. [Google Scholar] [CrossRef]
- Albuquerque, B.; Häussler, A.; Vannoni, E.; Wolfer, D.P.; Tegeder, I. Learning and memory with neuropathic pain: Impact of old age and progranulin deficiency. Front. Behav. Neurosci. 2013, 7, 174. [Google Scholar] [CrossRef] [Green Version]
- Tran, B.N.; Valek, L.; Wilken-Schmitz, A.; Fuhrmann, D.C.; Namgaladze, D.; Wittig, I.; Tegeder, I. Reduced exploratory behavior in neuronal nucleoredoxin knockout mice. Redox Biol. 2021, 45, 102054. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Koprich, J.B.; Wang, Y.; Yu, W.-B.; Xiao, B.-G.; Brotchie, J.M.; Wang, J. Treatment with Trehalose Prevents Behavioral and Neurochemical Deficits Produced in an AAV α-Synuclein Rat Model of Parkinson’s Disease. Mol. Neurobiol. 2015, 53, 2258–2268. [Google Scholar] [CrossRef]
- Schaeffer, V.; Lavenir, I.; Ozcelik, S.; Tolnay, M.; Winkler, D.T.; Goedert, M. Stimulation of autophagy reduces neurodegeneration in a mouse model of human tauopathy. Brain 2012, 135, 2169–2177. [Google Scholar] [CrossRef] [Green Version]
- Tien, N.T.; Karaca, I.; Tamboli, I.Y.; Walter, J. Trehalose Alters Subcellular Trafficking and the Metabolism of the Alzheimer-associated Amyloid Precursor Protein. J. Biol. Chem. 2016, 291, 10528–10540. [Google Scholar] [CrossRef] [Green Version]
- Tanji, K.; Miki, Y.; Maruyama, A.; Mimura, J.; Matsumiya, T.; Mori, F.; Imaizumi, T.; Itoh, K.; Wakabayashi, K. Trehalose intake induces chaperone molecules along with autophagy in a mouse model of Lewy body disease. Biochem. Biophys. Res. Commun. 2015, 465, 746–752. [Google Scholar] [CrossRef]
- Maday, S.; Holzbaur, E.L. Autophagosome assembly and cargo capture in the distal axon. Autophagy 2012, 8, 858–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maday, S.; Holzbaur, E.L. Autophagosome Biogenesis in Primary Neurons Follows an Ordered and Spatially Regulated Pathway. Dev. Cell 2014, 30, 71–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tegeder, I.; Kögel, D. When lipid homeostasis runs havoc: Lipotoxicity links lysosomal dysfunction to autophagy. Matrix Biol. 2021, 100-101, 99–117. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.; Higgins, C.B.; Heitmeier, M.R.; Kraft, T.; Qian, X.; Crowley, J.R.; Hyrc, K.L.; Beatty, W.L.; Yarasheski, K.; Hruz, P.; et al. SLC2A8 (GLUT8) is a mammalian trehalose transporter required for trehalose-induced autophagy. Sci. Rep. 2016, 6, 38586. [Google Scholar] [CrossRef]
- De Toledo, F.W.; Grundler, F.; Sirtori, C.R.; Ruscica, M. Unravelling the health effects of fasting: A long road from obesity treatment to healthy life span increase and improved cognition. Ann. Med. 2020, 52, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Javaheri, A.; Godar, R.J.; Murphy, J.; Ma, X.; Rohatgi, N.; Mahadevan, J.; Hyrc, K.; Saftig, P.; Marshall, C.; et al. Intermittent fasting preserves beta-cell mass in obesity-induced diabetes via the autophagy-lysosome pathway. Autophagy 2017, 13, 1952–1968. [Google Scholar] [CrossRef] [PubMed]
- Martin, B.; Mattson, M.P.; Maudsley, S. Caloric restriction and intermittent fasting: Two potential diets for successful brain aging. Ageing Res. Rev. 2006, 5, 332–353. [Google Scholar] [CrossRef] [Green Version]
- Pratscher, S.; Mickle, A.; Marks, J.; Rocha, H.; Bartsch, F.; Schmidt, J.; Tejera, L.; Garcia, S.; Custodero, C.; Jean, F.; et al. Optimizing Chronic Pain Treatment with Enhanced Neuroplastic Responsiveness: A Pilot Randomized Controlled Trial. Nutrients 2021, 13, 1556. [Google Scholar] [CrossRef]
- Smith, K.M.; Fagel, D.M.; Stevens, H.E.; Rabenstein, R.L.; Maragnoli, M.E.; Ohkubo, Y.; Picciotto, M.R.; Schwartz, M.L.; Vaccarino, F.M. Deficiency in Inhibitory Cortical Interneurons Associates with Hyperactivity in Fibroblast Growth Factor Receptor 1 Mutant Mice. Biol. Psychiatry 2008, 63, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Le Merre, P.; Esmaeili, V.; Charrière, E.; Galan, K.; Salin, P.-A.; Petersen, C.C.; Crochet, S. Reward-Based Learning Drives Rapid Sensory Signals in Medial Prefrontal Cortex and Dorsal Hippocampus Necessary for Goal-Directed Behavior. Neuron 2018, 97, 83–91.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouaichi, C.G.; Vincis, R. Cortical processing of chemosensory and hedonic features of taste in active licking mice. J. Neurophysiol. 2020, 123, 1995–2009. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kraft, V.; Schmitz, K.; Wilken-Schmitz, A.; Geisslinger, G.; Sisignano, M.; Tegeder, I. Trehalose Reduces Nerve Injury Induced Nociception in Mice but Negatively Affects Alertness. Nutrients 2021, 13, 2953. https://doi.org/10.3390/nu13092953
Kraft V, Schmitz K, Wilken-Schmitz A, Geisslinger G, Sisignano M, Tegeder I. Trehalose Reduces Nerve Injury Induced Nociception in Mice but Negatively Affects Alertness. Nutrients. 2021; 13(9):2953. https://doi.org/10.3390/nu13092953
Chicago/Turabian StyleKraft, Vanessa, Katja Schmitz, Annett Wilken-Schmitz, Gerd Geisslinger, Marco Sisignano, and Irmgard Tegeder. 2021. "Trehalose Reduces Nerve Injury Induced Nociception in Mice but Negatively Affects Alertness" Nutrients 13, no. 9: 2953. https://doi.org/10.3390/nu13092953
APA StyleKraft, V., Schmitz, K., Wilken-Schmitz, A., Geisslinger, G., Sisignano, M., & Tegeder, I. (2021). Trehalose Reduces Nerve Injury Induced Nociception in Mice but Negatively Affects Alertness. Nutrients, 13(9), 2953. https://doi.org/10.3390/nu13092953