
Bovine Colostrum Supplementation Improves Bone Metabolism in an Osteoporosis-Induced Animal Model

 , , , , ,
, , , , ,  ,
,  ,
,  ,
,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bovine Colostrum Preparation
2.2. Animal Care and Use
2.3. Study Design
2.4. Bone Biomarkers
2.5. Bone Microarchitecture (MicroCT)
2.6. Mechanical Properties
2.7. Gene Expression
2.8. Statistical Analyses
3. Results
3.1. Bone Microarchitecture
3.2. Mechanical Properties
3.3. Bone Biomarkers
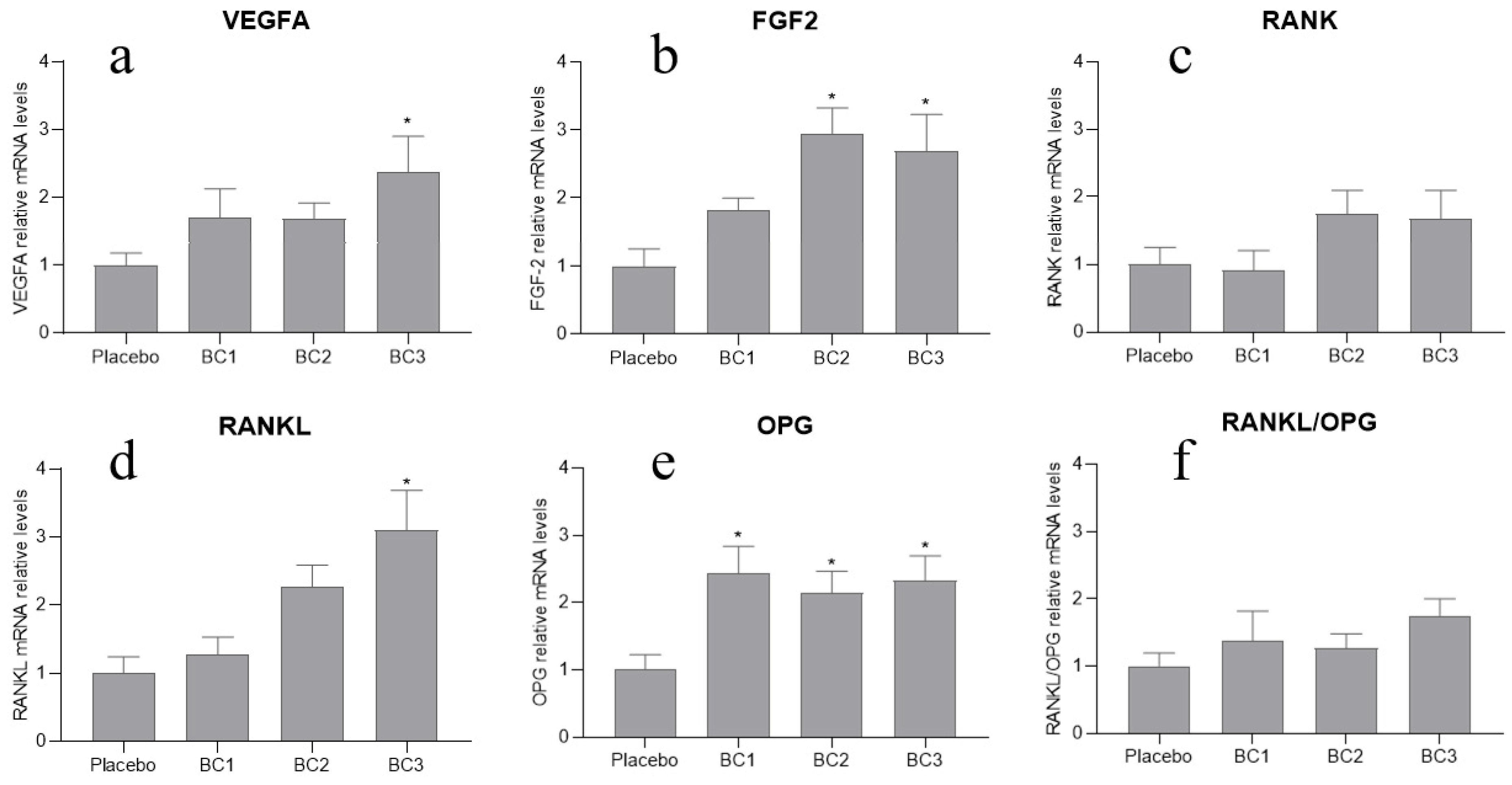
3.4. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peck, W.A. Consensus development conference: Diagnosis, prophylaxis, and treatment of osteoporosis. Am. J. Med. 1993, 94, 646–650. [Google Scholar] [CrossRef]
- On Osteoporosis, N.C.D.P.; Prevention, D. Osteoporosis prevention, diagnosis, and therapy. JAMA 2001, 285, 785–795. [Google Scholar] [CrossRef]
- Bliuc, D.; Nguyen, N.D.; Alarkawi, D.; Nguyen, T.V.; Eisman, J.A.; Center, J.R. Accelerated bone loss and increased post-fracture mortality in elderly women and men. Osteoporos. Int. 2015, 26, 1331–1339. [Google Scholar] [CrossRef]
- Cauley, J.A. Public health impact of osteoporosis. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1243–1251. [Google Scholar] [CrossRef] [Green Version]
- Adachi, J.D.; Ioannidis, G.; Pickard, L.; Berger, C.; Prior, J.C.; Joseph, L.; Hanley, D.A.; Olszynski, W.P.; Murray, T.M.; Anastassiades, T.; et al. The association between osteoporotic fractures and health-related quality of life as measured by the Health Utilities Index in the Canadian Multicentre Osteoporosis Study (CaMos). Osteoporos. Int. 2003, 14, 895–904. [Google Scholar] [CrossRef]
- Amorim, T.; Wyon, M.; Maia, J.; Machado, J.C.; Marques, F.; Metsios, G.S.; Flouris, A.D.; Koutedakis, Y. Prevalence of low bone mineral density in female dancers. Sports Med. 2015, 45, 257–268. [Google Scholar] [CrossRef]
- Amorim, T.; Koutedakis, Y.; Nevill, A.; Wyon, M.; Maia, J.; Machado, J.C.; Marques, F.; Metsios, G.S.; Flouris, A.D.; Adubeiro, N.; et al. Bone mineral density in vocational and professional ballet dancers. Osteoporos. Int. 2017, 28, 2903–2912. [Google Scholar] [CrossRef]
- Hernlund, E.; Svedbom, A.; Ivergård, M.; Compston, J.; Cooper, C.; Stenmark, J.; McCloskey, E.V.; Jönsson, B.; Kanis, J.A. Osteoporosis in the European Union: Medical management, epidemiology and economic burden. A report prepared in collaboration with the International Osteoporosis Foundation (IOF) and the European Federation of Pharmaceutical Industry Associations (EFPIA). Arch. Osteoporos. 2013, 8, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosla, S.; Shane, E. A Crisis in the Treatment of Osteoporosis. J. Bone Miner. Res. 2016, 31, 1485–1487. [Google Scholar] [CrossRef] [Green Version]
- Adler, R.A.; El-Hajj Fuleihan, G.; Bauer, D.C.; Camacho, P.M.; Clarke, B.L.; Clines, G.A.; Compston, J.E.; Drake, M.T.; Edwards, B.J.; Favus, M.J.; et al. Managing Osteoporosis in Patients on Long-Term Bisphosphonate Treatment: Report of a Task Force of the American Society for Bone and Mineral Research. J. Bone Miner. Res. 2016, 31, 16–35. [Google Scholar] [CrossRef] [PubMed]
- Watts, N.B.; Brown, J.P.; Papapoulos, S.; Lewiecki, E.M.; Kendler, D.L.; Dakin, P.; Wagman, R.B.; Wang, A.; Daizadeh, N.S.; Smith, S.; et al. Safety Observations With 3 Years of Denosumab Exposure: Comparison Between Subjects Who Received Denosumab During the Randomized FREEDOM Trial and Subjects Who Crossed Over to Denosumab During the FREEDOM Extension. J. Bone Miner. Res. 2017, 32, 1481–1485. [Google Scholar] [CrossRef] [Green Version]
- Diker-Cohen, T.; Rosenberg, D.; Avni, T.; Shepshelovich, D.; Tsvetov, G.; Gafter-Gvili, A. Risk for Infections during Treatment with Denosumab for Osteoporosis: A Systematic Review and Meta-analysis. J. Clin. Endocrinol. Metab. 2020, 105, 1641–1658. [Google Scholar] [CrossRef]
- Boquete-Castro, A.; Gómez-Moreno, G.; Calvo-Guirado, J.L.; Aguilar-Salvatierra, A.; Delgado-Ruiz, R.A. Denosumab and osteonecrosis of the jaw. A systematic analysis of events reported in clinical trials. Clin. Oral Implant. Res. 2016, 27, 367–375. [Google Scholar] [CrossRef]
- Ismail, A.; Bekhet, A.; Ibrahim Abushouk, A.; Mirbaha, S.; Baratloo, A. Denosumab and Atypical Femoral Fractures: A Scoping Literature Review. Trauma Mon. 2017, 23. [Google Scholar] [CrossRef]
- Khorsand, I.; Kashef, R.; Ghazanfarpour, M.; Mansouri, E.; Dashti, S.; Khadivzadeh, T. The Beneficial and Adverse Effects of Raloxifene in Menopausal Women: A Mini Review. J. Menopausal Med. 2018, 24, 183–187. [Google Scholar] [CrossRef]
- Grady, D.; Ettinger, B.; Moscarelli, E.; Plouffe, L.; Sarkar, S.; Ciaccia, A.; Cummings, S. Safety and adverse effects associated with raloxifene: Multiple outcomes of raloxifene evaluation. Obstet. Gynecol. 2004, 104, 837–844. [Google Scholar] [CrossRef]
- Overman, R.A.; Borse, M.; Gourlay, M.L. Salmon calcitonin use and associated cancer risk. Ann. Pharmacother. 2013, 47, 1675–1684. [Google Scholar] [CrossRef]
- Wells, G.; Chernoff, J.; Gilligan, J.P.; Krause, D.S. Does salmon calcitonin cause cancer? A review and meta-analysis. Osteoporos. Int. 2016, 27, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacNabb, C.; Patton, D.; Hayes, J.S. Sclerostin Antibody Therapy for the Treatment of Osteoporosis: Clinical Prospects and Challenges. J. Osteoporos. 2016, 2016, 6217286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosman, F.; de Beur, S.J.; LeBoff, M.S.; Lewiecki, E.M.; Tanner, B.; Randall, S.; Lindsay, R. Clinician’s Guide to Prevention and Treatment of Osteoporosis. Osteoporos. Int. 2014, 25, 2359–2381. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Crockett, J.C. Osteoporosis—A current view of pharmacological prevention and treatment. Drug Des. Devel. Ther. 2013, 7, 435–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amrein, K.; Scherkl, M.; Hoffmann, M.; Neuwersch-Sommeregger, S.; Köstenberger, M.; Tmava Berisha, A.; Martucci, G.; Pilz, S.; Malle, O. Vitamin D deficiency 2.0: An update on the current status worldwide. Eur. J. Clin. Nutr. 2020, 74, 1498–1513. [Google Scholar] [CrossRef] [PubMed]
- Coronado-Zarco, R.; Olascoaga-Gómez de León, A.; García-Lara, A.; Quinzaños-Fresnedo, J.; Nava-Bringas, T.I.; Macías-Hernández, S.I. Nonpharmacological interventions for osteoporosis treatment: Systematic review of clinical practice guidelines. Osteoporos. Sarcopenia 2019, 5, 69–77. [Google Scholar] [CrossRef]
- Wolman, R.; Wyon, M.A.; Koutedakis, Y.; Nevill, A.M.; Eastell, R.; Allen, N. Vitamin D status in professional ballet dancers: Winter vs. summer. J. Sci. Med. Sport 2013, 16, 388–391. [Google Scholar] [CrossRef] [Green Version]
- Wyon, M.A.; Wolman, R.; Nevill, A.M.; Cloak, R.; Metsios, G.S.; Gould, D.; Ingham, A.; Koutedakis, Y. Acute Effects of Vitamin D3 Supplementation on Muscle Strength in Judoka Athletes: A Randomized Placebo-Controlled, Double-Blind Trial. Clin. J. Sport Med. 2016, 26, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Wang, X.-F.; Li, D.-Y.; Chen, Y.-C.; Zhao, L.-J.; Liu, X.-G.; Guo, Y.-F.; Shen, J.; Lin, X.; Deng, J.; et al. The good, the bad, and the ugly of calcium supplementation: A review of calcium intake on human health. Clin. Interv. Aging 2018, 13, 2443–2452. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.M.; Gallagher, J.C.; Suiter, C. Medium doses of daily vitamin D decrease falls and higher doses of daily vitamin D3 increase falls: A randomized clinical trial. J. Steroid Biochem. Mol. Biol. 2017, 173, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Burt, L.A.; Billington, E.O.; Rose, M.S.; Raymond, D.A.; Hanley, D.A.; Boyd, S.K. Effect of High-Dose Vitamin D Supplementation on Volumetric Bone Density and Bone Strength: A Randomized Clinical Trial. JAMA 2019, 322, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Letavernier, E.; Daudon, M.; Vitamin, D. Hypercalciuria and Kidney Stones. Nutrients 2018, 10, 366. [Google Scholar] [CrossRef] [Green Version]
- Bolland, M.J.; Avenell, A.; Baron, J.A.; Grey, A.; MacLennan, G.S.; Gamble, G.D.; Reid, I.R. Effect of calcium supplements on risk of myocardial infarction and cardiovascular events: Meta-analysis. BMJ 2010, 341, c3691. [Google Scholar] [CrossRef] [Green Version]
- Reid, I.R. Efficacy, effectiveness and side effects of medications used to prevent fractures. J. Intern. Med. 2015, 277, 690–706. [Google Scholar] [CrossRef]
- Tripathi, V.; Vashishtha, B. Bioactive Compounds of Colostrum and Its Application. Food Rev. Int. 2006, 22, 225–244. [Google Scholar] [CrossRef]
- Bagwe, S.; Tharappel, L.J.; Kaur, G.; Buttar, H.S. Bovine colostrum: An emerging nutraceutical. J. Complement. Integr. Med. 2015, 12, 175–185. [Google Scholar] [CrossRef]
- Godhia, M.; Patel, N. Colostrum—Its Composition, Benefits As A Nutraceutical: A Review. Curr. Res. Nutr. Food Sci. J. 2013, 1, 37–47. [Google Scholar] [CrossRef]
- Godden, S. Colostrum management for dairy calves. Vet. Clin. North Am. Food Anim. Pract. 2008, 24, 19–39. [Google Scholar] [CrossRef] [PubMed]
- Superti, F. Lactoferrin from Bovine Milk: A Protective Companion for Life. Nutrients 2020, 12, 2562. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-R.; Kim, H.-M.; Choi, H.-S.; Hong, J.H. Effects of Colostrum Basic Protein from Colostrum Whey Protein: Increases in Osteoblast Proliferation and Bone Metabolism. J. Food Sci. Nutr. 2007, 12, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Du, M.; Xu, W.; Yi, H.; Han, X.; Wang, C.; Zhang, L. Protective effects of bovine colostrum acid proteins on bone loss of ovariectomized rats and the ingredients identification. Mol. Nutr. Food Res. 2011, 55, 220–228. [Google Scholar] [CrossRef]
- Yun, B.; Maburutse, B.E.; Kang, M.; Park, M.R.; Park, D.J.; Kim, Y.; Oh, S. Short communication: Dietary bovine milk-derived exosomes improve bone health in an osteoporosis-induced mouse model. J. Dairy Sci. 2020, 103, 7752–7760. [Google Scholar] [CrossRef]
- Lee, J.; Kwon, S.H.; Kim, H.M.; Fahey, S.N.; Knighton, D.R.; Sansom, A. Effect of a Growth Protein-Colostrum Fraction on bone development in juvenile rats. Biosci. Biotechnol. Biochem. 2008, 72, 1–6. [Google Scholar] [CrossRef]
- Cornish, J.; Callon, K.E.; Naot, D.; Palmano, K.P.; Banovic, T.; Bava, U.; Watson, M.; Lin, J.M.; Tong, P.C.; Chen, Q.; et al. Lactoferrin is a potent regulator of bone cell activity and increases bone formation in vivo. Endocrinology 2004, 145, 4366–4374. [Google Scholar] [CrossRef] [Green Version]
- Naot, D.; Chhana, A.; Matthews, B.G.; Callon, K.E.; Tong, P.C.; Lin, J.-M.; Costa, J.L.; Watson, M.; Grey, A.B.; Cornish, J. Molecular mechanisms involved in the mitogenic effect of lactoferrin in osteoblasts. Bone 2011, 49, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Wang, K.; Wu, C.; Zhang, L. Effects of bovine colostrum acid protein on bone loss and hemobiochemistry indexes in rats. Dairy Sci. Technol. 2009, 89, 449–461. [Google Scholar] [CrossRef] [Green Version]
- Lelovas, P.P.; Xanthos, T.T.; Thoma, S.E.; Lyritis, G.P.; Dontas, I.A. The laboratory rat as an animal model for osteoporosis research. Comp. Med. 2008, 58, 424–430. [Google Scholar] [PubMed]
- Yousefzadeh, N.; Kashfi, K.; Jeddi, S.; Ghasemi, A. Ovariectomized rat model of osteoporosis: A practical guide. EXCLI J. 2020, 19, 89–107. [Google Scholar] [CrossRef] [PubMed]
- Potikanond, S.; Rattanachote, P.; Pintana, H.; Suntornsaratoon, P.; Charoenphandhu, N.; Chattipakorn, N.; Chattipakorn, S. Obesity does not aggravate osteoporosis or osteoblastic insulin resistance in orchiectomized rats. J. Endocrinol. 2016, 228, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.-M.; Xue, Y.; Lin, Q.-M. Bovine lactoferrin improves bone mass and microstructure in ovariectomized rats via OPG/RANKL/RANK pathway. Acta Pharmacol. Sin. 2012, 33, 1277–1284. [Google Scholar] [CrossRef] [Green Version]
- Comelekoglu, U.; Bagis, S.; Yalin, S.; Ogenler, O.; Yildiz, A.; Sahin, N.O.; Oguz, I.; Hatungil, R. Biomechanical evaluation in osteoporosis: Ovariectomized rat model. Clin. Rheumatol. 2007, 26, 380–384. [Google Scholar] [CrossRef]
- Luu, A.N.; Anez-Bustillos, L.; Aran, S.; Araiza Arroyo, F.J.; Entezari, V.; Rosso, C.; Snyder, B.D.; Nazarian, A. Microstructural, densitometric and metabolic variations in bones from rats with normal or altered skeletal states. PLoS ONE 2013, 8, e82709. [Google Scholar] [CrossRef] [Green Version]
- Ekeland, A.; Engesæter, L.B.; Langeland, N. Mechanical Properties of Fractured and Intact Rat Femora Evaluated by Bending, Torsional and Tensile Tests. Acta Orthop. Scand. 1981, 52, 605–613. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.Y.; Jiang, L.; Ibrahim, S.A.; Zhang, L.; Zhang, H.; Zhang, M.; Ren, F.Z. Orally administered lactoferrin preserves bone mass and microarchitecture in ovariectomized rats. J. Nutr. 2009, 139, 958–964. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J. Statistical Power Analysis for the Behavioral Sciences; L. Erlbaum Associates: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Price, P.A.; Parthemore, J.G.; Deftos, L.J. New biochemical marker for bone metabolism. Measurement by radioimmunoassay of bone GLA protein in the plasma of normal subjects and patients with bone disease. J. Clin. Investig. 1980, 66, 878–883. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Guo, H.Y.; Zhang, W.; Wen, P.C.; Zhang, H.; Guo, Z.R.; Ren, F.Z. Effect of iron saturation level of lactoferrin on osteogenic activity in vitro and in vivo. J. Dairy Sci. 2013, 96, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grey, A.; Banovic, T.; Zhu, Q.; Watson, M.; Callon, K.; Palmano, K.; Ross, J.; Naot, D.; Reid, I.R.; Cornish, J. The low-density lipoprotein receptor-related protein 1 is a mitogenic receptor for lactoferrin in osteoblastic cells. Mol. Endocrinol. 2004, 18, 2268–2278. [Google Scholar] [CrossRef] [Green Version]
- Grey, A.; Zhu, Q.; Watson, M.; Callon, K.; Cornish, J. Lactoferrin potently inhibits osteoblast apoptosis, via an LRP1-independent pathway. Mol. Cell Endocrinol. 2006, 251, 96–102. [Google Scholar] [CrossRef]
- Blais, A.; Malet, A.; Mikogami, T.; Martin-Rouas, C.; Tomé, D. Oral bovine lactoferrin improves bone status of ovariectomized mice. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E1281–E1288. [Google Scholar] [CrossRef] [PubMed]
- Lorget, F.; Clough, J.; Oliveira, M.; Daury, M.C.; Sabokbar, A.; Offord, E. Lactoferrin reduces in vitro osteoclast differentiation and resorbing activity. Biochem. Biophys. Res. Commun. 2002, 296, 261–266. [Google Scholar] [CrossRef]
- Cornish, J.; Naot, D. Lactoferrin as an effector molecule in the skeleton. Biometals 2010, 23, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Bienko, M.; Wolski, D.; Lis, A.; Radzki, R.; Filip, D.; Polak, P. Densitometric, tomographic and mechanical parameters of the female Wistar rat skeletal system after lactoferrin and colostrum treatment in the condition of gonadectomy-induced osteopenia. Med. Weter. 2016, 72, 580–586. [Google Scholar] [CrossRef] [Green Version]
- Fan, F.; Shi, P.; Liu, M.; Chen, H.; Tu, M.; Lu, W.; Du, M. Lactoferrin preserves bone homeostasis by regulating the RANKL/RANK/OPG pathway of osteoimmunology. Food Funct. 2018, 9, 2653–2660. [Google Scholar] [CrossRef]
- Bharadwaj, S.; Naidu, A.G.; Betageri, G.V.; Prasadarao, N.V.; Naidu, A.S. Milk ribonuclease-enriched lactoferrin induces positive effects on bone turnover markers in postmenopausal women. Osteoporos. Int. 2009, 20, 1603–1611. [Google Scholar] [CrossRef] [PubMed]
- Boyce, B.F.; Xing, L. Functions of RANKL/RANK/OPG in bone modeling and remodeling. Arch. Biochem. Biophys. 2008, 473, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Lacey, D.L.; Timms, E.; Tan, H.L.; Kelley, M.J.; Dunstan, C.R.; Burgess, T.; Elliott, R.; Colombero, A.; Elliott, G.; Scully, S.; et al. Osteoprotegerin Ligand is a Cytokine that Regulates Osteoclast Differentiation and Activation. Cell 1998, 93, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Kong, Y.Y.; Yoshida, H.; Sarosi, I.; Tan, H.L.; Timms, E.; Capparelli, C.; Morony, S.; Oliveira-dos-Santos, A.J.; Van, G.; Itie, A.; et al. OPGL is a key regulator of osteoclastogenesis, lymphocyte development and lymph-node organogenesis. Nature 1999, 397, 315–323. [Google Scholar] [CrossRef]
- Burgess, T.L.; Qian, Y.; Kaufman, S.; Ring, B.D.; Van, G.; Capparelli, C.; Kelley, M.; Hsu, H.; Boyle, W.J.; Dunstan, C.R.; et al. The ligand for osteoprotegerin (OPGL) directly activates mature osteoclasts. J. Cell Biol. 1999, 145, 527–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.; Lacey, D.L.; Dunstan, C.R.; Solovyev, I.; Colombero, A.; Timms, E.; Tan, H.L.; Elliott, G.; Kelley, M.J.; Sarosi, I.; et al. Tumor necrosis factor receptor family member RANK mediates osteoclast differentiation and activation induced by osteoprotegerin ligand. Proc. Natl. Acad. Sci. USA 1999, 96, 3540–3545. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, N.; Kinosaki, M.; Yamaguchi, K.; Shima, N.; Yasuda, H.; Yano, K.; Morinaga, T.; Higashio, K. RANK is the essential signaling receptor for osteoclast differentiation factor in osteoclastogenesis. Biochem. Biophys. Res. Commun. 1998, 253, 395–400. [Google Scholar] [CrossRef]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Guerrini, M.M.; Takayanagi, H. The immune system, bone and RANKL. Arch. Biochem. Biophys. 2014, 561, 118–123. [Google Scholar] [CrossRef]
- Mitchner, N.A.; Harris, S.T. Current and emerging therapies for osteoporosis. J. Fam. Pract. 2009, 58, S45–S49. [Google Scholar]
- Simonet, W.S.; Lacey, D.L.; Dunstan, C.R.; Kelley, M.; Chang, M.S.; Lüthy, R.; Nguyen, H.Q.; Wooden, S.; Bennett, L.; Boone, T.; et al. Osteoprotegerin: A novel secreted protein involved in the regulation of bone density. Cell 1997, 89, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Deckers, M.M.; Karperien, M.; van der Bent, C.; Yamashita, T.; Papapoulos, S.E.; Löwik, C.W. Expression of vascular endothelial growth factors and their receptors during osteoblast differentiation. Endocrinology 2000, 141, 1667–1674. [Google Scholar] [CrossRef] [PubMed]
- Mayr-Wohlfart, U.; Waltenberger, J.; Hausser, H.; Kessler, S.; Günther, K.P.; Dehio, C.; Puhl, W.; Brenner, R.E. Vascular endothelial growth factor stimulates chemotactic migration of primary human osteoblasts. Bone 2002, 30, 472–477. [Google Scholar] [CrossRef]
- Midy, V.; Plouët, J. Vasculotropin/vascular endothelial growth factor induces differentiation in cultured osteoblasts. Biochem. Biophys. Res. Commun. 1994, 199, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Olsen, B.R. Osteoblast-derived VEGF regulates osteoblast differentiation and bone formation during bone repair. J. Clin. Invest. 2016, 126, 509–526. [Google Scholar] [CrossRef] [Green Version]
- Gerber, H.P.; Vu, T.H.; Ryan, A.M.; Kowalski, J.; Werb, Z.; Ferrara, N. VEGF couples hypertrophic cartilage remodeling, ossification and angiogenesis during endochondral bone formation. Nat. Med. 1999, 5, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Zelzer, E.; McLean, W.; Ng, Y.S.; Fukai, N.; Reginato, A.M.; Lovejoy, S.; D’Amore, P.A.; Olsen, B.R. Skeletal defects in VEGF(120/120) mice reveal multiple roles for VEGF in skeletogenesis. Development 2002, 129, 1893–1904. [Google Scholar] [CrossRef]
- Mayahara, H.; Ito, T.; Nagai, H.; Miyajima, H.; Tsukuda, R.; Taketomi, S.; Mizoguchi, J.; Kato, K. In vivo stimulation of endosteal bone formation by basic fibroblast growth factor in rats. Growth Factors 1993, 9, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Kurokawa, T.; Aoyama, I.; Hanada, K.; Tamura, M.; Kawaguchi, H. Stimulation of bone formation by intraosseous injection of basic fibroblast growth factor in ovariectomised rats. Int. Orthop. 1998, 22, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, H.; Oka, H.; Jingushi, S.; Izumi, T.; Fukunaga, M.; Sato, K.; Matsushita, T.; Nakamura, K. A local application of recombinant human fibroblast growth factor 2 for tibial shaft fractures: A randomized, placebo-controlled trial. J. Bone Miner. Res. 2010, 25, 2735–2743. [Google Scholar] [CrossRef] [PubMed]
- Debiais, F.; Hott, M.; Graulet, A.M.; Marie, P.J. The effects of fibroblast growth factor-2 on human neonatal calvaria osteoblastic cells are differentiation stage specific. J. Bone Miner. Res. 1998, 13, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Marie, P.J. Fibroblast growth factor signaling controlling osteoblast differentiation. Gene 2003, 316, 23–32. [Google Scholar] [CrossRef]
- Montero, A.; Okada, Y.; Tomita, M.; Ito, M.; Tsurukami, H.; Nakamura, T.; Doetschman, T.; Coffin, J.D.; Hurley, M.M. Disruption of the fibroblast growth factor-2 gene results in decreased bone mass and bone formation. J. Clin. Investig. 2000, 105, 1085–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, M.; Kellow, N.J.; Dordevic, A.L.; Evans, S.; Caissutti, J.; McCaffrey, T.A. Health Benefits of Whey or Colostrum Supplementation in Adults ≥35 Years; a Systematic Review. Nutrients 2020, 12, 299. [Google Scholar] [CrossRef] [Green Version]
- Duff, W.R.D.; Chilibeck, P.D.; Rooke, J.J.; Kaviani, M.; Krentz, J.R.; Haines, D.M. The Effect of Bovine Colostrum Supplementation in Older Adults During Resistance Training. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 276–285. [Google Scholar] [CrossRef]


{kind=link}
| Pre-Intervention | 4 Months BC Supplementation | Post-Intervention |
|---|---|---|
| Surgery: Ovariectomy and orchidectomy | Placebo: 0.5 g/day/OVX/ORX rats | Euthanasia: Blood collection: bone formation and resorption markers. Right tibia removal: micro-CT, mechanical testing. Left tibia removal: gene expression. |
| BC dose 1: 0.5 g/day/OVX rats 1.0 g/day/ORX rats | ||
| BC dose 2: 1.0 g/day/OVX rats 1.5 g/day/ORX rats | ||
| BC dose 3: 1.5 g/day/OVX rats 2.0 g/day/ORX rats |
| Gene | Primers (5′-3′) | Conditions |
|---|---|---|
| FGF2 | CAAAACCTGACCCGATCCCT | 95 °C, 3 s |
| AGAATCTGTCCCGTTCGGC | 62 °C, 20 s | |
| 72 °C, 15 s | ||
| VEGF-A | GCAGCGACAAGGCAGACTA | 95 °C, 3 s |
| GAGTGAAGGAGCAACCTCTCC | 64 °C, 20 s | |
| 72 °C, 15 s | ||
| OPG | AGGGCATACTTCCTGTTGCC | 95 °C, 3 s |
| CACAGCACAGCCACTTGTTC | 62 °C, 20 s | |
| 72 °C, 15 s | ||
| RANKL | ATTGTCCAGTCGCACTTCGT | 95 °C, 3 s |
| AGTCGAGTCCTGCAAACCTG | 62 °C, 20 s | |
| 72 °C, 15 s | ||
| RANK | TGGCCCGGATGAATACTTGG | 95 °C, 3 s |
| GCACACTGTGTCCTTGTTGAG | 63 °C, 20 s | |
| 72 °C, 15 s | ||
| TATA | AAGGTTCCCTCCTCTGCACT | 95 °C, 3 s |
| TGTACAGGTGGCTTGAACACT | 62 °C, 20 s | |
| 72 °C, 15 s | ||
| GAPDH | CTATAAATTGAGCCCGCAGCC | 95 °C, 3 s |
| CCTTCCCCATGGTGTCTGAG | 55 °C, 20 s | |
| 72 °C, 15 s | ||
| B-actin | TTTCTGCGCAAGTTAGGTTTT | 95 °C, 3 s |
| TTTCTGCGCAAGTTAGGTTTT | 60 °C, 20 s | |
| 72 °C, 15 s |
| Post-Intervention | ||||
|---|---|---|---|---|
| Analyzed Parameter | Placebo | BC1 | BC2 | BC3 |
| Cortical bone | ||||
| Porosity (%) | ||||
| ORX rats | 29.48 ± 4.24 | 39.56 ± 15.47 * | 25.51 ± 13.64 | 25.91 ± 7.39 |
| OVX rats | 26.56 ± 11.14 | 68.03 ± 14.32 ** | 25.16 ± 8.83 | 25.22 ± 8.54 |
| Volume (% BV/TV) | ||||
| ORX rats | 70.43 ± 4.13 | 60.44 ± 15.47 * | 74.49 ± 13.64 | 74.09 ± 7.39 |
| OVX rats | 73.36 ± 11.15 | 31.97 ± 14.32 ** | 74.84 ± 8.83 | 74.78 ± 8.54 |
| BMD (g/cm3) | ||||
| ORX rats | 2.84 ± 0.33 | 2.56 ± 0.91 | 3.17 ± 0.44 | 2.93 ± 0.37 |
| OVX rats | 2.93 ± 0.29 | 1.29 ± 0.63 ** | 2.33 ± 0.73 | 2.83 ± 0.31 |
| BMC (g) | ||||
| ORX rats | 71.97 ± 12.94 | 71.43 ± 11.41 | 74.53 ± 23.13 | 78.93 ± 10.89 |
| OVX rats | 71.21 ± 9.65 | 88.01 ± 7.50 ** | 74.99 ± 17.55 | 71.78 ± 14.76 |
| Trabecular bone | ||||
| Porosity (%) | ||||
| ORX rats | 87.17 ± 4.14 | 89.56 ± 3.13 | 84.23 ± 4.26 | 84.92 ± 4.22 |
| OVX rats | 87.21 ± 1.97 | 92.47 ± 3.85 * | 86.62 ± 4.81 | 85.26 ± 2.24 |
| Separation (µm) | ||||
| ORX rats | 113.02 ± 96.58 | 150.04 ± 51.30 | 86.44 ± 38.52 | 77.91 ± 22.43 |
| OVX rats | 163.12 ± 86.59 | 179.55 ± 50.98 | 145.79 ± 91.25 | 104.57 ± 47.40 |
| Thickness (µm) | ||||
| ORX rats | 16.44 ± 1.40 | 15.45 ± 2.42 | 17.91 ± 3.66 | 16.78 ± 1.55 |
| OVX rats | 25.73 ± 20.54 | 12.65 ± 3.64 ** | 21.11 ± 3.28 | 18.39 ± 2.45 |
| Volume (% BV/TV) | ||||
| ORX rats | 12.82 ± 4.14 | 10.44 ± 3.13 | 15.77 ± 4.26 | 15.08 ± 4.22 |
| OVX rats | 12.78 ± 1.97 | 7.53 ± 3.85 * | 13.75 ± 4.93 | 14.74 ± 2.24 |
| BMD (g/cm3) | ||||
| ORX rats | 1.23 ± 0.20 | 1.15 ± 0.19 | 1.29 ± 0.22 | 1.27 ± 0.25 |
| OVX rats | 1.19 ± 0.12 | 1.07 ± 0.09 ** | 0.97 ± 0.28 | 1.22 ± 0.12 |
| BMC (g) | ||||
| ORX rats | 64.81 ± 12.86 | 66.25 ± 17.12 | 78.61 ± 10.60 * | 74.19 ± 10.19 |
| OVX rats | 61.23 ± 10.18 | 94.98 ± 12.48 ** | 71.59 ± 19.31 | 72.71 ± 13.40 * |
| Post-Intervention | ||||
|---|---|---|---|---|
| Analyzed Parameter | Placebo | BC1 | BC2 | BC3 |
| Max. tensile strength (σ, MPa) | ||||
| ORX rats | 3.84 ± 0.63 | 4.36 ± 0.90 | 5.00 ± 0.64 | 6.00 ± 0.45 |
| OVX rats | 8.00 ± 0.75 | 5.04 ± 0.76 | 4.86 ± 1.02 | 6.22 ± 1.74 |
| Elastic modulus (E, MPa) | ||||
| ORX rats | 151.77 ± 35.31 | 147.79 ± 9.30 | 192.33 ± 36.19 | 239.05 ± 21.42 |
| OVX rats | 385.06 ± 54.14 | 254.25 ± 53.54 | 202.02 ± 5.58 | 277.45 ± 74.13 |
| Stress at yield (σy, MPa) | ||||
| ORX rats | 1.88 ± 0.34 | 1.98 ± 0.78 | 2.39 ± 0.36 | 2.50 ± 1.03 |
| OVX rats | 3.53 ± 0.46 | 2.17 ± 0.44 | 2.83 ± 1.86 | 3.11 ± 0.46 |
| Strain at yield (εy, %) | ||||
| ORX rats | 1.37 ± 0.22 | 1.47 ± 0.45 | 1.39 ± 0.15 | 1.21 ± 0.43 |
| OVX rats | 1.10 ± 0.02 | 1.09 ± 0.31 | 1.45 ± 0.50 | 1.32 ± 0.26 |
| Post-Intervention | ||||
|---|---|---|---|---|
| Analyzed Parameter | Placebo | BC1 | BC2 | BC3 |
| Alkaline phosphatase (U/L) | ||||
| ORX rats | 114.50 ± 10.74 | 103.14 ± 14.01 | 97.03 ± 17.19 * | 119.73 ± 21.86 |
| OVX rats | 92.08 ± 26.29 | 72.83 ± 19.93 | 75.76 ± 26.08 | 70.8 ± 19.87 |
| Osteocalcin (µg/L) | ||||
| ORX rats | 10.71 ± 0.58 | 12.47 ± 1.44 * | 13.74 ± 1.51 ** | 16.58 ± 1.54 ** |
| OVX rats | 13.35 ± 2.47 | 12.24 ± 1.14 | 11.09 ± 1.58 | 15.59 ± 2.24 |
| Deoxypyridinoline (µg/L) | ||||
| ORX rats | 0.44 ± 0.04 | 0.49 ± 0.05 * | 0.45 ± 0.13 | 0.37 ± 0.11 |
| OVX rats | 0.43 ± 0.16 | 0.37 ± 0.09 | 0.44 ± 0.10 | 0.34 ± 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kydonaki, E.K.; Freitas, L.; Fonseca, B.M.; Reguengo, H.; Raposo Simón, C.; Bastos, A.R.; Fernandes, E.M.; Canadas, R.F.; Oliveira, J.M.; Correlo, V.M.; et al. Bovine Colostrum Supplementation Improves Bone Metabolism in an Osteoporosis-Induced Animal Model. Nutrients 2021, 13, 2981. https://doi.org/10.3390/nu13092981
Kydonaki EK, Freitas L, Fonseca BM, Reguengo H, Raposo Simón C, Bastos AR, Fernandes EM, Canadas RF, Oliveira JM, Correlo VM, et al. Bovine Colostrum Supplementation Improves Bone Metabolism in an Osteoporosis-Induced Animal Model. Nutrients. 2021; 13(9):2981. https://doi.org/10.3390/nu13092981
Chicago/Turabian StyleKydonaki, Eirini K., Laura Freitas, Bruno M. Fonseca, Henrique Reguengo, Carlos Raposo Simón, Ana R. Bastos, Emanuel M. Fernandes, Raphaël F. Canadas, Joaquim Miguel Oliveira, Vitor M. Correlo, and et al. 2021. "Bovine Colostrum Supplementation Improves Bone Metabolism in an Osteoporosis-Induced Animal Model" Nutrients 13, no. 9: 2981. https://doi.org/10.3390/nu13092981
APA StyleKydonaki, E. K., Freitas, L., Fonseca, B. M., Reguengo, H., Raposo Simón, C., Bastos, A. R., Fernandes, E. M., Canadas, R. F., Oliveira, J. M., Correlo, V. M., Reis, R. L., Vliora, M., Gkiata, P., Koutedakis, Y., Ntina, G., Pinto, R., Carrillo, A. E., Marques, F., & Amorim, T. (2021). Bovine Colostrum Supplementation Improves Bone Metabolism in an Osteoporosis-Induced Animal Model. Nutrients, 13(9), 2981. https://doi.org/10.3390/nu13092981