
Prevention of Loperamide-Induced Constipation in Mice and Alteration of 5-Hydroxytryotamine Signaling by Ligilactobacillus salivarius Li01


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Cultivation
2.2. Establishment of Animal Models and Experimental Design
2.3. Measurement of Water Content in Mouse Feces
2.4. Determination of GI Transit Rate
2.5. Biochemical Analysis
2.6. Quantitative PCR Assay
2.7. Histological Analysis
2.8. 16S rRNA Gene Analysis
2.9. Data Analysis
3. Results and Discussion
3.1. Establishment of a Constipation Mouse Model and Effects of Treatment
3.2. Effects of Li01 on Gut Motility and Fluid Transit in Constipated Mice and Associated Parameters
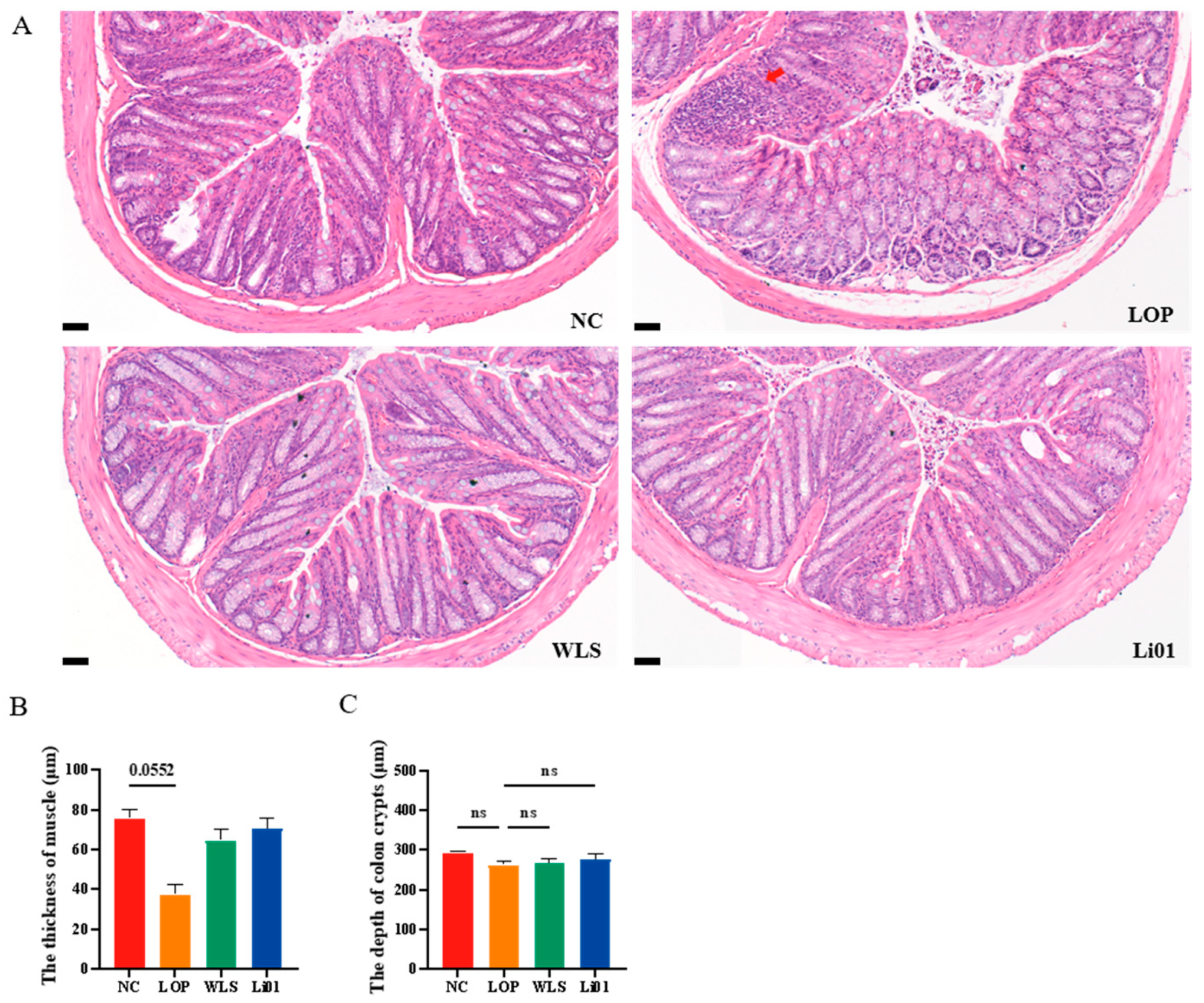
3.3. Effects of Li01 on the Intestinal Barrier Structure in Constipated Mice
3.4. Alteration of the 5-HT Signaling Pathway
3.5. Alteration of the Gut Microbiota Composition
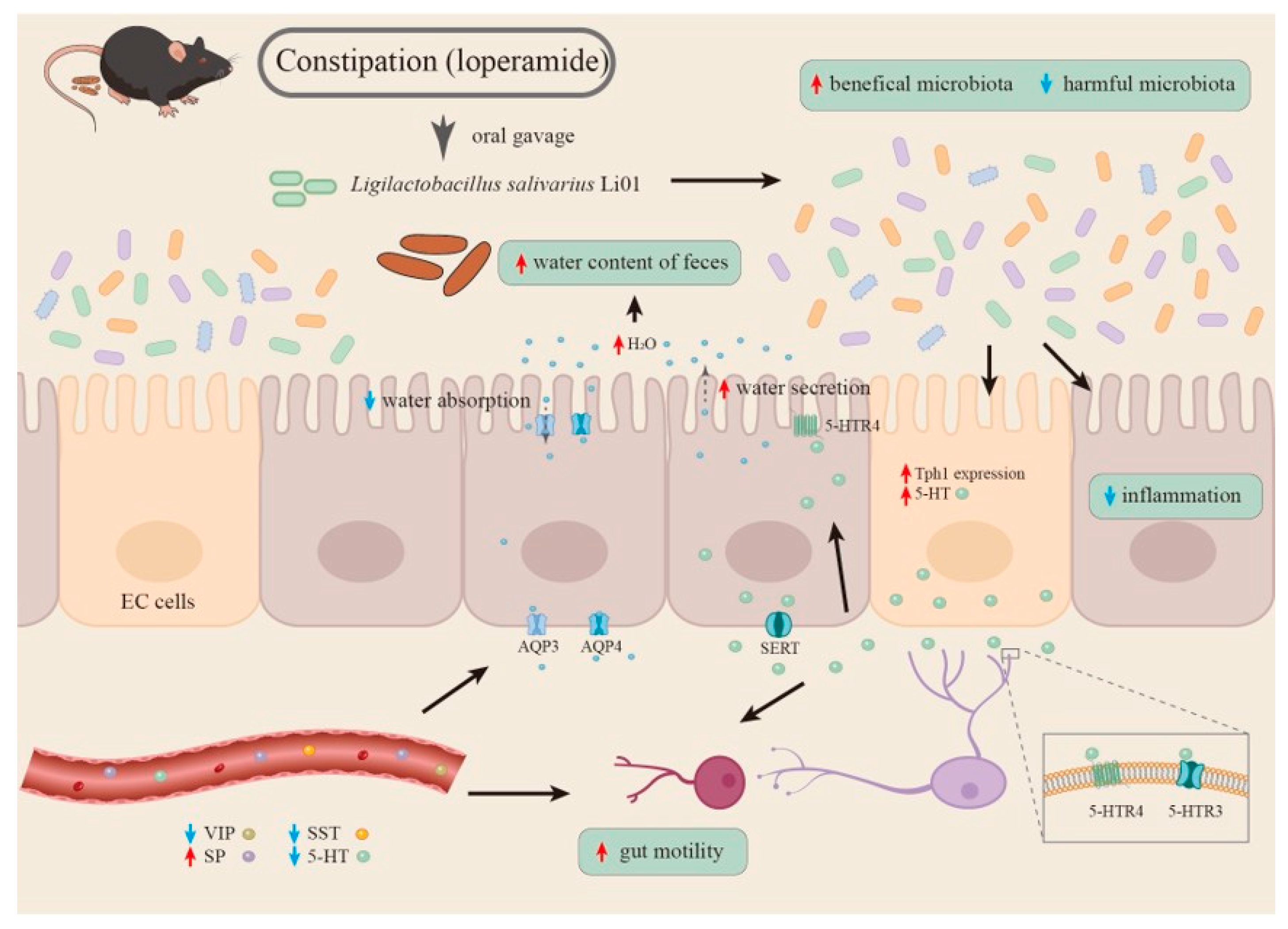
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rao, S.S.C.; Rattanakovit, K.; Patcharatrakul, T. Diagnosis and management of chronic constipation in adults. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Bharucha, A.E.; Lacy, B.E. Mechanisms, Evaluation, and Management of Chronic Constipation. Gastroenterology 2020, 158, 1232–1249.e1233. [Google Scholar] [CrossRef] [PubMed]
- Camilleri, M.; Ford, A.C.; Mawe, G.M.; Dinning, P.G.; Rao, S.S.; Chey, W.D.; Simren, M.; Lembo, A.; Young-Fadok, T.M.; Chang, L. Chronic constipation. Nat. Rev. Dis. Prim. 2017, 3, 17095. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Chen, Y.Y.; Xu, D.Q.; Yue, S.J.; Fu, R.J.; Yang, J.; Xing, L.M.; Tang, Y.P. Action Mode of Gut Motility, Fluid and Electrolyte Transport in Chronic Constipation. Front. Pharmacol. 2021, 12, 630249. [Google Scholar] [CrossRef] [PubMed]
- Vriesman, M.H.; Koppen, I.J.N.; Camilleri, M.; Di Lorenzo, C.; Benninga, M.A. Management of functional constipation in children and adults. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 21–39. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, P.C.; Marcobal, A.; Ursell, L.K.; Larauche, M.; Duboc, H.; Earle, K.A.; Sonnenburg, E.D.; Ferreyra, J.A.; Higginbottom, S.K.; Million, M.; et al. Complex interactions among diet, gastrointestinal transit, and gut microbiota in humanized mice. Gastroenterology 2013, 144, 967–977. [Google Scholar] [CrossRef] [Green Version]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.; Lu, T.; Wu, J.; Fan, D.; Liu, B.; Zhu, X.; Guo, H.; Du, Y.; Liu, F.; Tian, Y.; et al. Gut microbiota drives macrophage-dependent self-renewal of intestinal stem cells via niche enteric serotonergic neurons. Cell Res. 2022, 32, 555–569. [Google Scholar] [CrossRef]
- Kendig, D.M.; Grider, J.R. Serotonin and colonic motility. Neurogastroenterol. Motil. 2015, 27, 899–905. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.K.; Gershon, M.D. CrossTalk proposal: 5-HT is necessary for peristalsis. J. Physiol. 2015, 593, 3225–3227. [Google Scholar] [CrossRef] [Green Version]
- Bhattarai, Y.; Williams, B.B.; Battaglioli, E.J.; Whitaker, W.R.; Till, L.; Grover, M.; Linden, D.R.; Akiba, Y.; Kandimalla, K.K.; Zachos, N.C.; et al. Gut Microbiota-Produced Tryptamine Activates an Epithelial G-Protein-Coupled Receptor to Increase Colonic Secretion. Cell Host Microbe 2018, 23, 775–785.e775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbara, G.; Stanghellini, V.; Brandi, G.; Cremon, C.; Di Nardo, G.; De Giorgio, R.; Corinaldesi, R. Interactions between commensal bacteria and gut sensorimotor function in health and disease. Am. J. Gastroenterol. 2005, 100, 2560–2568. [Google Scholar] [CrossRef] [PubMed]
- Dimidi, E.; Mark Scott, S.; Whelan, K. Probiotics and constipation: Mechanisms of action, evidence for effectiveness and utilisation by patients and healthcare professionals. Proc. Nutr. Soc. 2020, 79, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Tan, A.H.; Lim, S.Y.; Chong, K.K.; MAA, A.M.; Hor, J.W.; Lim, J.L.; Low, S.C.; Chong, C.W.; Mahadeva, S.; Lang, A.E. Probiotics for Constipation in Parkinson Disease: A Randomized Placebo-Controlled Study. Neurology 2021, 96, e772–e782. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Park, M.Y.; Brooks, I.; Lee, J.; Kim, S.H.; Kim, J.Y.; Oh, B.; Kim, J.W.; Kwon, O. Spore-forming Bacillus coagulans SNZ 1969 improved intestinal motility and constipation perception mediated by microbial alterations in healthy adults with mild intermittent constipation: A randomized controlled trial. Food Res. Int. 2021, 146, 110428. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Lu, Y.; Zhang, T.; Xie, J.; Han, S.; Zhang, S.; Fei, Y.; Ling, Z.; Wu, J.; Hu, Y.; et al. Improved functionality of Ligilactobacillus salivarius Li01 in alleviating colonic inflammation by layer-by-layer microencapsulation. NPJ Biofilms Microbiomes 2021, 7, 58. [Google Scholar] [CrossRef]
- Lv, L.-X.; Hu, X.-J.; Qian, G.-R.; Zhang, H.; Lu, H.-F.; Zheng, B.-W.; Jiang, L.; Li, L.-J. Administration of Lactobacillus salivarius LI01 or Pediococcus pentosaceus LI05 improves acute liver injury induced by d-galactosamine in rats. Appl. Microbiol. Biotechnol. 2014, 98, 5619–5632. [Google Scholar] [CrossRef]
- Fei, Y.; Zhang, S.; Han, S.; Qiu, B.; Lu, Y.; Huang, W.; Li, F.; Chen, D.; Berglund, B.; Xiao, H.; et al. The Role of Dihydroresveratrol in Enhancing the Synergistic Effect of Ligilactobacillus salivarius Li01 and Resveratrol in Ameliorating Colitis in Mice. Research 2022, 2022, 9863845. [Google Scholar] [CrossRef]
- Wang, L.; Chai, M.; Wang, J.; Yu, Q.; Wang, G.; Zhang, H.; Zhao, J.; Chen, W. Bifidobacterium longum relieves constipation by regulating the intestinal barrier of mice. Food Funct. 2022, 13, 5037–5049. [Google Scholar] [CrossRef]
- Bahrudin, M.F.; Abdul Rani, R.; Tamil, A.M.; Mokhtar, N.M.; Raja Ali, R.A. Effectiveness of Sterilized Symbiotic Drink Containing Lactobacillus helveticus Comparable to Probiotic Alone in Patients with Constipation-Predominant Irritable Bowel Syndrome. Dig. Dis. Sci. 2020, 65, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, C.; Cui, S.; Lee, Y.K.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Adhesive Bifidobacterium Induced Changes in Cecal Microbiome Alleviated Constipation in Mice. Front. Microbiol. 2019, 10, 1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, W.; Han, Y.; Yang, Y.; Hao, Z.; An, N.; Chen, J.; Zhang, Z.; Gao, X.; Storey, K.B.; Chang, H.; et al. Dynamic Changes in Colonic Structure and Protein Expression Suggest Regulatory Mechanisms of Colonic Barrier Function in Torpor-Arousal Cycles of the Daurian Ground Squirrel. Int. J. Mol. Sci. 2022, 23, 9026. [Google Scholar] [CrossRef] [PubMed]
- Wald, A. Constipation: Advances in Diagnosis and Treatment. JAMA 2016, 315, 185–191. [Google Scholar] [CrossRef]
- Basilisco, G.; Bozzani, A.; Camboni, G.; Recchia, M.; Quatrini, M.; Conte, D.; Penagini, R.; Bianchi, P.A. Effect of loperamide and naloxone on mouth-to-caecum transit time evaluated by lactulose hydrogen breath test. Gut 1985, 26, 700–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ollitrault, P.; Dolladille, C.; Chretien, B.; Milliez, P.; Alexandre, J. Cardiovascular Toxicities Associated With Loperamide: Analysis of the World Health Organization Pharmacovigilance Database. Circulation 2021, 143, 403–405. [Google Scholar] [CrossRef]
- Bai, J.; Cai, Y.; Huang, Z.; Gu, Y.; Huang, N.; Sun, R.; Zhang, G.; Liu, R. Shouhui Tongbian Capsule ameliorates constipation via gut microbiota-5-HT-intestinal motility axis. Biomed. Pharm. 2022, 154, 113627. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Lin, B.; Zhang, Y.; Xie, Z.; Zheng, Y.; Wang, Q.; Xiao, H. Bamboo shavings derived O-acetylated xylan alleviates loperamide-induced constipation in mice. Carbohydr. Polym. 2022, 276, 118761. [Google Scholar] [CrossRef]
- Wang, L.; Hu, L.; Xu, Q.; Yin, B.; Fang, D.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacterium adolescentis Exerts Strain-Specific Effects on Constipation Induced by Loperamide in BALB/c Mice. Int. J. Mol. Sci. 2017, 18, 318. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.J.; Park, J.W.; Kim, J.E.; Lee, S.J.; Gong, J.E.; Jung, Y.S.; Seo, S.; Hwang, D.Y. Novel Characterization of Constipation Phenotypes in ICR Mice Orally Administrated with Polystyrene Microplastics. Int. J. Mol. Sci. 2021, 22, 5845. [Google Scholar] [CrossRef]
- Cao, P.Q.; Li, X.P.; Ou-Yang, J.; Jiang, R.G.; Huang, F.F.; Wen, B.B.; Zhang, X.N.; Huang, J.A.; Liu, Z.H. The protective effects of yellow tea extract against loperamide-induced constipation in mice. Food Funct. 2021, 12, 5621–5636. [Google Scholar] [CrossRef]
- Hu, T.G.; Wen, P.; Fu, H.Z.; Lin, G.Y.; Liao, S.T.; Zou, Y.X. Protective effect of mulberry (Morus atropurpurea) fruit against diphenoxylate-induced constipation in mice through the modulation of gut microbiota. Food Funct. 2019, 10, 1513–1528. [Google Scholar] [CrossRef]
- Verbeure, W.; van Goor, H.; Mori, H.; van Beek, A.P.; Tack, J.; van Dijk, P.R. The Role of Gasotransmitters in Gut Peptide Actions. Front. Pharmacol. 2021, 12, 720703. [Google Scholar] [CrossRef] [PubMed]
- King, S.K.; Sutcliffe, J.R.; Ong, S.Y.; Lee, M.; Koh, T.L.; Wong, S.Q.; Farmer, P.J.; Peck, C.J.; Stanton, M.P.; Keck, J.; et al. Substance P and vasoactive intestinal peptide are reduced in right transverse colon in pediatric slow-transit constipation. Neurogastroenterol. Motil. 2010, 22, 883–892.e234. [Google Scholar] [CrossRef] [PubMed]
- El-Salhy, M.; Norrgard, O. Colonic neuroendocrine peptide levels in patients with chronic idiopathic slow transit constipation. Ups. J. Med. Sci. 1998, 103, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Sisto, M.; Ribatti, D.; Lisi, S. Aquaporin water channels: New perspectives on the potential role in inflammation. Adv. Protein Chem. Struct. Biol. 2019, 116, 311–345. [Google Scholar] [CrossRef]
- Itoh, A.; Tsujikawa, T.; Fujiyama, Y.; Bamba, T. Enhancement of aquaporin-3 by vasoactive intestinal polypeptide in a human colonic epithelial cell line. J. Gastroenterol. Hepatol. 2003, 18, 203–210. [Google Scholar] [CrossRef]
- Zhu, S.; Ran, J.; Yang, B.; Mei, Z. Aquaporins in Digestive System. Adv. Exp. Med. Biol. 2017, 969, 123–130. [Google Scholar] [CrossRef]
- Cao, Y.; He, Y.; Wei, C.; Li, J.; Qu, L.; Zhang, H.; Cheng, Y.; Qiao, B. Aquaporins Alteration Profiles Revealed Different Actions of Senna, Sennosides, and Sennoside A in Diarrhea-Rats. Int. J. Mol. Sci. 2018, 19, 3210. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Yu, Z.; Zhang, Z.; Liang, X.; Gong, P.; Yi, H.; Yang, L.; Liu, T.; Shi, H.; Zhang, L. Bifidobacterium animalis F1-7 in combination with konjac glucomannan improves constipation in mice via humoral transport. Food Funct. 2021, 12, 791–801. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, J.; Zhang, Z.; Liang, X.; Liu, T.; Yi, H.; Gong, P.; Wang, L.; Yang, W.; Zhang, X.; et al. Konjac glucomannan with probiotics acts as a combination laxative to relieve constipation in mice by increasing short-chain fatty acid metabolism and 5-hydroxytryptamine hormone release. Nutrition 2021, 84, 111112. [Google Scholar] [CrossRef]
- Lincoln, J.; Crowe, R.; Kamm, M.A.; Burnstock, G.; Lennard-Jones, J.E. Serotonin and 5-Hydroxyindoleacetic Acid Are Increased in the Sigmoid Colon in Severe Idiopathic Constipation. Gastroenterology 1990, 98, 1219–1225. [Google Scholar] [CrossRef]
- Costedio, M.M.; Coates, M.D.; Brooks, E.M.; Glass, L.M.; Ganguly, E.K.; Blaszyk, H.; Ciolino, A.L.; Wood, M.J.; Strader, D.; Hyman, N.H.; et al. Mucosal serotonin signaling is altered in chronic constipation but not in opiate-induced constipation. Am. J. Gastroenterol. 2010, 105, 1173–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.; Singh, R.; Ha, S.E.; Martin, A.M.; Jones, L.A.; Jin, B.; Jorgensen, B.G.; Zogg, H.; Chervo, T.; Gottfried-Blackmore, A.; et al. Serotonin Deficiency Is Associated With Delayed Gastric Emptying. Gastroenterology 2021, 160, 2451–2466.e2419. [Google Scholar] [CrossRef] [PubMed]
- Gershon, M.D.; Tack, J. The serotonin signaling system: From basic understanding to drug development for functional GI disorders. Gastroenterology 2007, 132, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Luo, J.; Li, J.; Kim, G.; Stewart, A.; Urban, J.F., Jr.; Huang, Y.; Chen, S.; Wu, L.G.; Chesler, A.; et al. Interleukin-33 Promotes Serotonin Release from Enterochromaffin Cells for Intestinal Homeostasis. Immunity 2021, 54, 151–163.e156. [Google Scholar] [CrossRef]
- Bharucha, A.E.; Wouters, M.M.; Tack, J. Existing and emerging therapies for managing constipation and diarrhea. Curr. Opin. Pharmacol. 2017, 37, 158–166. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Bortolanza, M.; Zhai, G.; Shang, A.; Ling, Z.; Jiang, B.; Shen, X.; Yao, Y.; Yu, J.; Li, L.; et al. Gut microbiota dysbiosis associated with plasma levels of Interferon-gamma and viral load in patients with acute hepatitis E infection. J. Med. Virol. 2022, 94, 692–702. [Google Scholar] [CrossRef]
- Yu, T.; Zheng, Y.P.; Tan, J.C.; Xiong, W.J.; Wang, Y.; Lin, L. Effects of Prebiotics and Synbiotics on Functional Constipation. Am. J. Med. Sci. 2017, 353, 282–292. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Yang, S.; Sun, S.; Si, Q.; Wang, L.; Zhang, Q.; Wu, G.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus rhamnosus Strains Relieve Loperamide-Induced Constipation via Different Pathways Independent of Short-Chain Fatty Acids. Front. Cell Infect. Microbiol. 2020, 10, 423. [Google Scholar] [CrossRef]
- Ezzine, C.; Loison, L.; Montbrion, N.; Bole-Feysot, C.; Dechelotte, P.; Coeffier, M.; Ribet, D. Fatty acids produced by the gut microbiota dampen host inflammatory responses by modulating intestinal SUMOylation. Gut Microbes 2022, 14, 2108280. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, G.; Chen, J.; Chen, X.; Chia, N.; O’Connor, H.M.; Wolf, P.G.; Gaskins, H.R.; Bharucha, A.E. Relationship Between Microbiota of the Colonic Mucosa vs Feces and Symptoms, Colonic Transit, and Methane Production in Female Patients With Chronic Constipation. Gastroenterology 2016, 150, 367–379.e361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Hu, L.; Yan, S.; Jiang, T.; Fang, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Effects of different oligosaccharides at various dosages on the composition of gut microbiota and short-chain fatty acids in mice with constipation. Food Funct. 2017, 8, 1966–1978. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.-B.; Kang, Z.-P.; Wang, M.-X.; Long, J.; Wang, H.-Y.; Huang, J.-Q.; Wei, S.-Y.; Zhou, W.; Zhao, H.-M.; Liu, D.-Y. Curcumin ameliorated dextran sulfate sodium-induced colitis via regulating the homeostasis of DCs and Treg and improving the composition of the gut microbiota. J. Funct. Foods 2021, 86, 104716. [Google Scholar] [CrossRef]
- Qiao, Y.; Qiu, Z.; Tian, F.; Yu, L.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Pediococcus acidilactici Strains Improve Constipation Symptoms and Regulate Intestinal Flora in Mice. Front. Cell. Infect. Microbiol. 2021, 11, 655258. [Google Scholar] [CrossRef]
- Dubin, K.; Callahan, M.K.; Ren, B.; Khanin, R.; Viale, A.; Ling, L.; No, D.; Gobourne, A.; Littmann, E.; Huttenhower, C.; et al. Intestinal microbiome analyses identify melanoma patients at risk for checkpoint-blockade-induced colitis. Nat. Commun. 2016, 7, 10391. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| GAPDH | CAGTGGCAAAGTGGAGATTGTTG | TCGCTCCTGGAAGATGGTGAT |
| AQP3 | GCCAAGGTAGGATAGCAAATAA | TTGAAAACTTGGTCCCTTGC |
| AQP4 | CACCATAAACTGGGGTGGCT | AGACGGACTTAGCGATGCTG |
| Tph1 | CCATCTTCCGAGAGCTAAACAAA | TCTTCCCGATAGCCACAGTATT |
| SERT | GCGACGTGAAGGAAATGCTG | GGAGTTGGGGTGGACTCATC |
| 5-HTR3 | CTGTGGCGATCACCGGAAG | GGCTGACTGCGTAGAATAAAGG |
| 5-HTR4 | TCGATCTTTCACCTGTGCTGTAT | TGTTGTTCCAGCCTTGCATTATG |
| Muc2 | AAGCTGCACGGACACCTCTATAT | TCGCTCTTGGTCAGGACATAGTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, B.; Zhu, L.; Zhang, S.; Han, S.; Fei, Y.; Ba, F.; Berglund, B.; Li, L.; Yao, M. Prevention of Loperamide-Induced Constipation in Mice and Alteration of 5-Hydroxytryotamine Signaling by Ligilactobacillus salivarius Li01. Nutrients 2022, 14, 4083. https://doi.org/10.3390/nu14194083
Qiu B, Zhu L, Zhang S, Han S, Fei Y, Ba F, Berglund B, Li L, Yao M. Prevention of Loperamide-Induced Constipation in Mice and Alteration of 5-Hydroxytryotamine Signaling by Ligilactobacillus salivarius Li01. Nutrients. 2022; 14(19):4083. https://doi.org/10.3390/nu14194083
Chicago/Turabian StyleQiu, Bo, Lian Zhu, Shuobo Zhang, Shengyi Han, Yiqiu Fei, Furong Ba, Björn Berglund, Lanjuan Li, and Mingfei Yao. 2022. "Prevention of Loperamide-Induced Constipation in Mice and Alteration of 5-Hydroxytryotamine Signaling by Ligilactobacillus salivarius Li01" Nutrients 14, no. 19: 4083. https://doi.org/10.3390/nu14194083
APA StyleQiu, B., Zhu, L., Zhang, S., Han, S., Fei, Y., Ba, F., Berglund, B., Li, L., & Yao, M. (2022). Prevention of Loperamide-Induced Constipation in Mice and Alteration of 5-Hydroxytryotamine Signaling by Ligilactobacillus salivarius Li01. Nutrients, 14(19), 4083. https://doi.org/10.3390/nu14194083