
The Roles of MTRR and MTHFR Gene Polymorphisms in Colorectal Cancer Survival


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Baseline Information
2.3. Study Outcomes
2.4. SNP Genotyping and Selection
2.5. Statistical Analysis
3. Results
3.1. Patient Baseline Characteristics
3.2. Gene-Level Association with CRC Survival
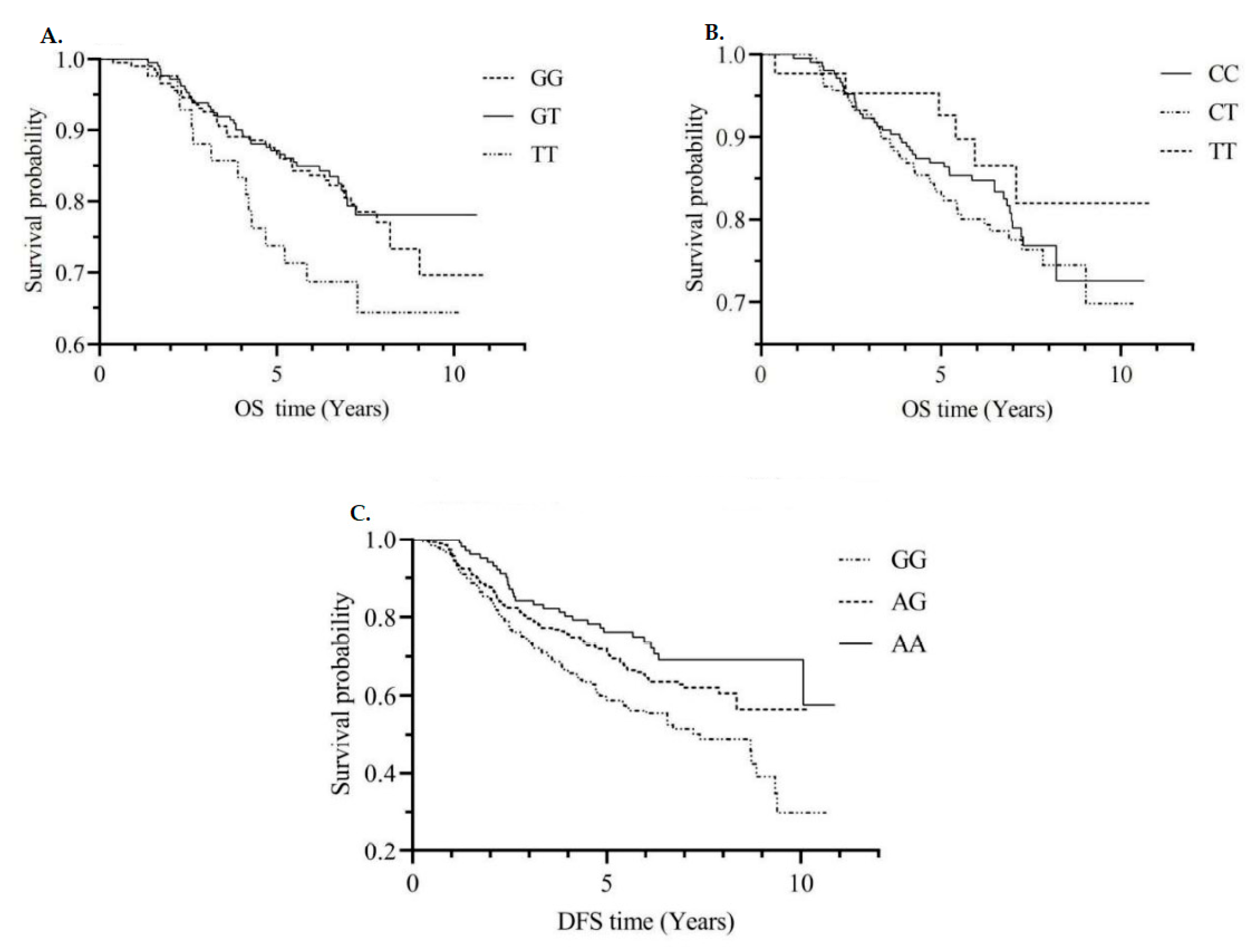
3.3. Single SNPs and CRC Survival
3.4. Associations between Haplotypes and CRC Survival
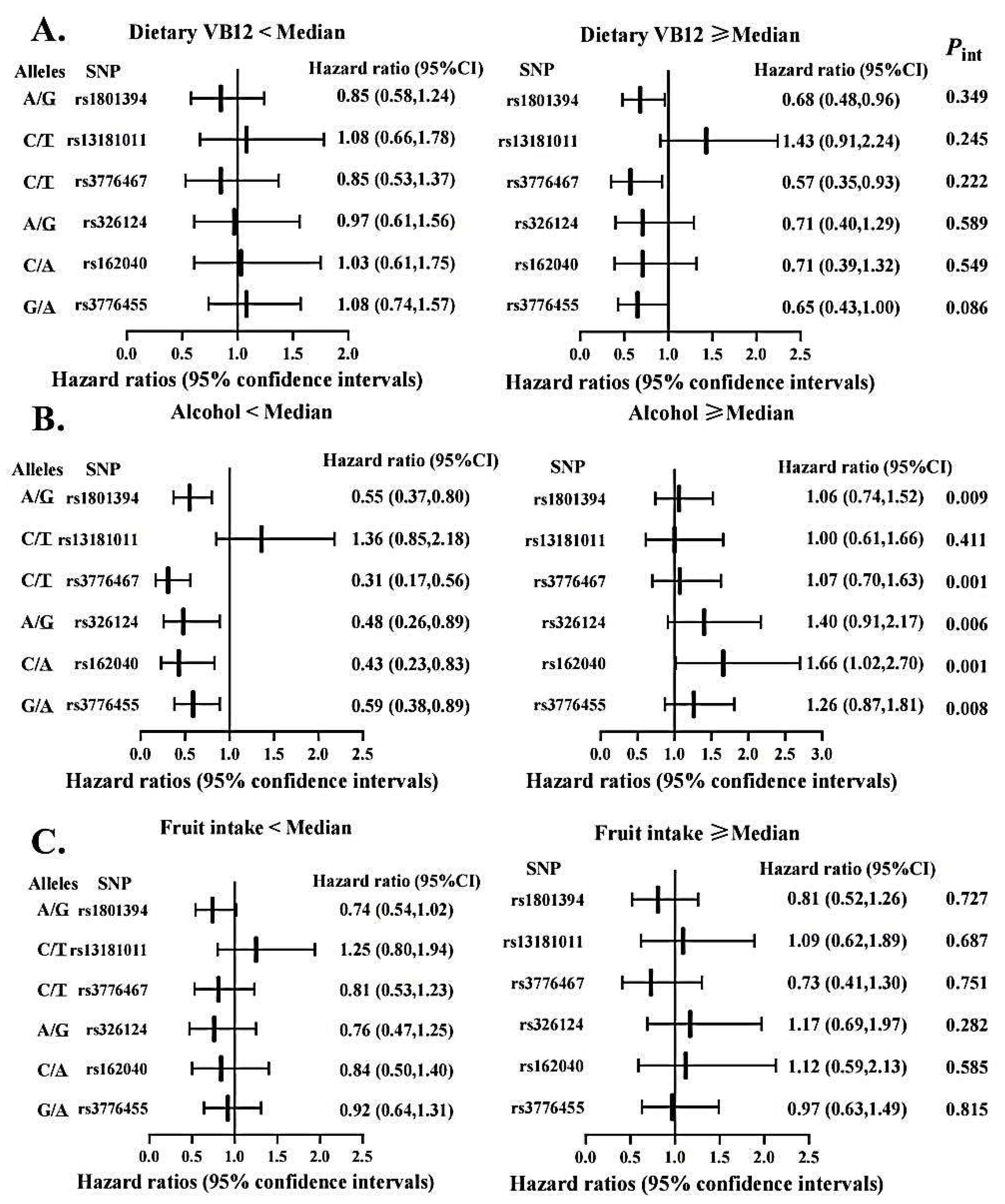
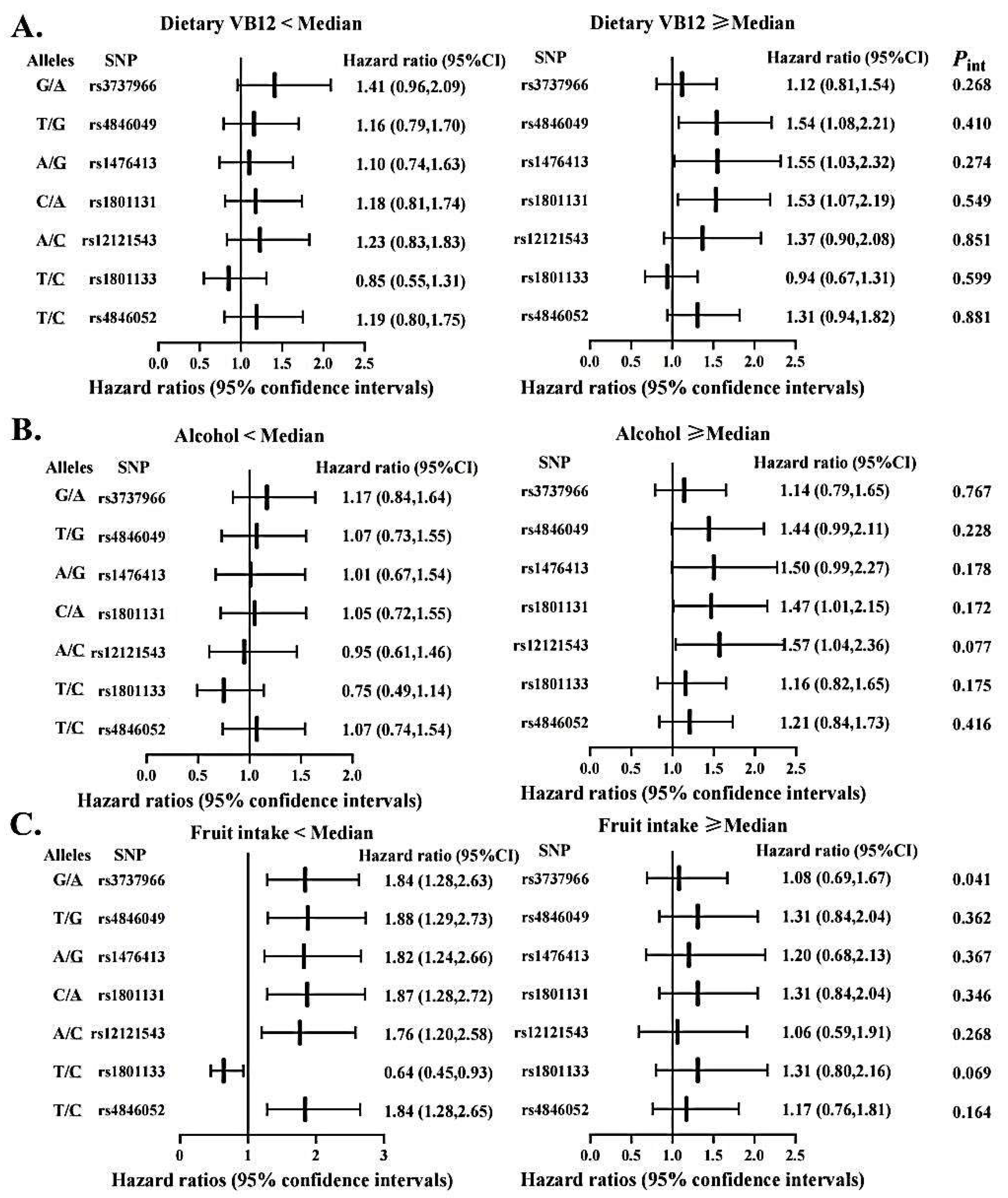
3.5. Gene-Diet Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xi, Y.; Xu, P. Global colorectal cancer burden in 2020 and projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef] [PubMed]
- Renee, P.; Stephanie, P.; Sharon, D.; Carolyn, L. Folate and Its Impact on Cancer Risk. Curr. Nutr. Rep. 2018, 7, 70–84. [Google Scholar]
- Williams, E.A. Folate, colorectal cancer and the involvement of DNA methylation. Proc. Nutr. Soc. 2012, 71, 592–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, D.H.; Lee, B.H.; Kang, S.H.; Lee, H.J.; Lim, S.Y.; Suh, Y.K.; Ahn, Y.O. Folate intake and the risk of colorectal cancer in a Korean population. Eur. J. Clin. Nutr. 2009, 63, 1057–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulrich, C.M. Nutrigenetics in cancer research—Folate metabolism and colorectal cancer. J. Nutr. 2005, 135, 2698–2702. [Google Scholar] [CrossRef] [Green Version]
- Eussen, S.J.; Vollset, S.E.; Igland, J.; Meyer, K.; Fredriksen, Å.; Ueland, P.M.; Jenab, M.; Slimani, N.; Boffetta, P.; Overvad, K.; et al. Plasma folate, related genetic variants, and colorectal cancer risk in EPIC. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1328–1340. [Google Scholar] [CrossRef] [Green Version]
- Hubner, R.A.; Houlston, R.S. Folate and colorectal cancer prevention. Br. J. Cancer 2009, 100, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Li, W.X.; Cheng, F.; Zhang, A.J.; Dai, S.X.; Li, G.H.; Lv, W.W.; Zhou, T.; Zhang, Q.; Zhang, H.; Zhang, T.; et al. Folate Deficiency and Gene Polymorphisms of MTHFR, MTR and MTRR Elevate the Hyperhomocysteinemia Risk. Clin. Lab. 2017, 63, 523–533. [Google Scholar] [CrossRef]
- Huang, Y.; Han, S.Z.; Li, Y.; Mao, Y.M.; Xie, Y. Different roles of MTHFR C677T and A1298C polymorphisms in colorectal adenoma and colorectal cancer: A meta-analysis. J. Hum. Genet. 2007, 52, 73–85. [Google Scholar] [CrossRef]
- Lin, W.; Yuhong, L.; Zhengrong, Z.; Zuoli, S.; Yi, H.; Rena, L. Methylenetetrahydrofolate reductase and psychiatric diseases. Transl. Psychiatry 2018, 8, 242. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Stampfer, M.J.; Giovannucci, E.; Artigas, C.; Hunter, D.J.; Fuchs, C.; Willett, W.C.; Selhub, J.; Hennekens, C.H.; Rozen, R. Methylenetetrahydrofolate reductase polymorphism, dietary interactions, and risk of colorectal cancer. Cancer Res. 1997, 57, 1098–1102. [Google Scholar] [PubMed]
- Park, K.S.; Mok, J.W.; Kim, J.C. The 677C>T mutation in 5,10-methylenetetrahydrofolate reductase and colorectal cancer risk. Genet. Test. 1999, 3, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Slattery, M.L.; Potter, J.D.; Samowitz, W.; Schaffer, D.; Leppert, M. Methylenetetrahydrofolate reductase, diet, and risk of colon cancer. Cancer Epidemiol. Biomark. Prev. 1999, 8, 513–518. [Google Scholar]
- Ulrich, C.M.; Kampman, E.; Bigler, J.; Schwartz, S.M.; Chen, C.; Bostick, R.; Fosdick, L.; Beresford, S.A.; Yasui, Y.; Potter, J.D. Colorectal adenomas and the C677T MTHFR polymorphism: Evidence for gene-environment interaction? Cancer Epidemiol. Biomark. Prev. 1999, 8, 659–668. [Google Scholar]
- Le Marchand, L.; Donlon, T.; Hankin, J.H.; Kolonel, L.N.; Wilkens, L.R.; Seifried, A. B-vitamin intake, metabolic genes, and colorectal cancer risk (United States). Cancer Causes Control 2002, 13, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ma, J.; Stampfer, M.J.; Palomeque, C.; Selhub, J.; Hunter, D.J. Linkage disequilibrium between the 677C>T and 1298A>C polymorphisms in human methylenetetrahydrofolate reductase gene and their contributions to risk of colorectal cancer. Pharmacogenetics 2002, 12, 339–342. [Google Scholar] [CrossRef]
- Keku, T.; Millikan, R.; Worley, K.; Winkel, S.; Eaton, A.; Biscocho, L.; Martin, C.; Sandler, R. 5,10-Methylenetetrahydrofolate reductase codon 677 and 1298 polymorphisms and colon cancer in African Americans and whites. Cancer Epidemiol. Biomark. Prev. 2002, 11, 1611–1621. [Google Scholar]
- Mahla, G.; Marjan, A.; Reza, K.; Javad, B.; Amin, K.M. Association of MTHFR C677T variant genotype with serum folate and Vit B12 in Iranian patients with colorectal cancer or adenomatous polyps. BMC Med. Genom. 2021, 14, 246. [Google Scholar] [CrossRef]
- Levine, A.J.; Siegmund, K.D.; Ervin, C.M.; Diep, A.; Lee, E.R.; Frankl, H.D.; Haile, R.W. The methylenetetrahydrofolate reductase 677C-->T polymorphism and distal colorectal adenoma risk. Cancer Epidemiol. Biomark. Prev. 2000, 9, 657–663. [Google Scholar]
- Jokic, M.; Brcic-Kostic, K.; Stefulj, J.; Ivkovic, T.C.; Bozo, L.; Gamulin, M.; Kapitanovic, S. Association of MTHFR, MTR, MTRR, RFC1, and DHFR Gene Polymorphisms with Susceptibility to Sporadic Colon Cancer. DNA Cell Biol. 2011, 30, 771–776. [Google Scholar] [CrossRef]
- Pardini, B.; Kumar, R.; Naccarati, A.; Prasad, R.B.; Forsti, A.; Polakova, V.; Vodickova, L.; Novotny, J.; Hemminki, K.; Vodicka, P. MTHFR and MTRR genotype and haplotype analysis and colorectal cancer susceptibility in a case-control study from the Czech Republic. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2011, 721, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Arve, U.; Emil, V.S.; Svein, H.; Randi, G.; Egil, J.; Magne, U.P. Colorectal cancer and the methylenetetrahydrofolate reductase 677C -> T and methionine synthase 2756A -> G polymorphisms: A study of 2168 case-control pairs from the JANUS cohort. J. Cancer Epidemiol. Biomark. Prev. 2004, 13, 2175–2180. [Google Scholar]
- Zhong, S.; Yang, J.H.; Liu, K.; Jiao, B.H.; Chang, Z.J. Quantitative assessment of the association between MTHFR C677T polymorphism and colorectal cancer risk in East Asians. Tumor Biol. 2012, 33, 2041–2051. [Google Scholar] [CrossRef]
- Keitaro, M.; Nobuyuki, H.; Takashi, H.; Tomoyuki, K.; Manami, I.; Toshiro, T.; Kazuo, T. Methionine Synthase Reductase Gene A66G Polymorphism is Associated with Risk of Colorectal Cancer. Asian Pac. J. Cancer Prev. APJCP 2002, 3, 353–359. [Google Scholar]
- Zhu, L.; Wang, F.; Hu, F.L.; Wang, Y.B.N.; Li, D.D.; Dong, X.S.; Cui, B.B.; Zhao, Y.S. Association between MTHFR polymorphisms and overall survival of colorectal cancer patients in Northeast China. Med. Oncol. 2013, 30, 6. [Google Scholar] [CrossRef] [PubMed]
- Xin-Lin, C.; Yu-Mei, W.; Fei, Z.; Zheng, C.; Xiaofei, Y.; Cong, S.; Yunpeng, G.; Tian-Ge, Y.; Guo, T.; Yi-Ming, C.; et al. Methylenetetrahydrofolate reductase polymorphisms and colorectal cancer prognosis: A meta-analysis. J. Gene Med. 2019, 21, e3114. [Google Scholar] [CrossRef] [Green Version]
- Negandhi, A.A.; Hyde, A.; Dicks, E.; Pollett, W.; Younghusband, B.H.; Parfrey, P.; Green, R.C.; Savas, S. MTHFR Glu429Ala and ERCC5 His46His Polymorphisms Are Associated with Prognosis in Colorectal Cancer Patients: Analysis of Two Independent Cohorts from Newfoundland. PLoS ONE 2013, 8, 14. [Google Scholar] [CrossRef]
- Osian, G.; Procopciuc, L.; Vlad, L.; Iancu, C.; Mocan, T.; Mocan, L. C677T and A1298C mutations in the MTHFR gene and survival in colorectal cancer. J. Gastrointest. Liver Dis. JGLD 2009, 18, 455–460. [Google Scholar]
- Green, R.C.; Green, J.S.; Buehler, S.K.; Robb, J.D.; Daftary, D.; Gallinger, S.; McLaughlin, J.R.; Parfrey, P.S.; Younghusband, H.B. Very high incidence of familial colorectal cancer in Newfoundland: A comparison with Ontario and 13 other population-based studies. Fam. Cancer 2007, 6, 53–62. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, S.R.; Wang, P.P.; Savas, S.; Wish, T.; Zhao, J.; Green, R.; Woods, M.; Sun, Z.; Roebothan, B.; et al. Influence of pre-diagnostic cigarette smoking on colorectal cancer survival: Overall and by tumour molecular phenotype. Br. J. Cancer 2014, 110, 1359–1366. [Google Scholar] [CrossRef] [Green Version]
- Woods, M.O.; Younghusband, H.B.; Parfrey, P.S.; Gallinger, S.; McLaughlin, J.; Dicks, E.; Stuckless, S.; Pollett, A.; Bapat, B.; Mrkonjic, M.; et al. The genetic basis of colorectal cancer in a population-based incident cohort with a high rate of familial disease. Gut 2010, 59, 1369–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Wang, P.P.; Roebothan, B.; Ryan, A.; Tucker, C.S.; Colbourne, J.; Baker, N.; Cotterchio, M.; Yi, Y.Q.; Sun, G. Assessing the validity of a self-administered food-frequency questionnaire (FFQ) in the adult population of Newfoundland and Labrador, Canada. Nutr. J. 2013, 12, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bakker, P.I.; Yelensky, R.; Pe’er, I.; Gabriel, S.B.; Daly, M.J.; Altshuler, D. Efficiency and power in genetic association studies. Nat. Genet. 2005, 37, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Gauderman, W.J.; Murcray, C.; Gilliland, F.; Conti, D.V. Testing association between disease and multiple SNPs in a candidate gene. Genet. Epidemiol. 2007, 31, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Conneely, K.N.; Boehnke, M. So Many Correlated Tests, So Little Time! Rapid Adjustment of P Values for Multiple Correlated Tests. Am. J. Hum. Genet. 2007, 81, 1158–1168. [Google Scholar] [CrossRef] [Green Version]
- Stephens, M.; Donnelly, P. A comparison of bayesian methods for haplotype reconstruction from population genotype data. Am. J. Hum. Genet. 2003, 73, 1162–1169. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Li, S.Q.; Wang, M.L.; He, J.; Xi, S.M. Association of MTRR A66G polymorphism with cancer susceptibility: Evidence from 85 studies. J. Cancer 2017, 8, 266–277. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.P.; Tang, R.N.; An, L. A meta-analysis of MTRR A66G polymorphism and colorectal cancer susceptibility. J. Buon 2015, 20, 918–922. [Google Scholar]
- Sharp, L.; Little, J. Polymorphisms in genes involved in folate metabolism and colorectal neoplasia: A HuGE review. Am. J. Epidemiol. 2004, 159, 423–443. [Google Scholar] [CrossRef] [Green Version]
- Mahasweta, C.; Tanusree, S.; Subhamita, M.; Swagata, S.; Kanchan, M. Folate System Gene Variant rs1801394 66AG may have a Causal Role in Down Syndrome in the Eastern Indian Population. Int. J. Mol. Cell. Med. 2020, 9, 215–224. [Google Scholar] [CrossRef]
- Manchun, S.; Manyi, S.; Li, Z.; Songli, S. Colorectal polyp risk is linked to an elevated level of homocysteine. Biosci. Rep. 2018, 38, BSR20171699. [Google Scholar] [CrossRef] [Green Version]
- Haerian, M.S.; Haerian, B.S.; Molanaei, S.; Kosari, F.; Sabeti, S.; Bidari-Zerehpoosh, F.; Abdolali, E. MTRR rs1801394 and its interaction with MTHFR rs1801133 in colorectal cancer: A case-control study and meta-analysis. Pharmacogenomics 2017, 18, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Shen, C.; Meng, X.; Bai, J.; Chen, F.; Yu, Y.; Jin, Y.; Fu, S. Methionine synthase reductase A66G polymorphism contributes to tumor susceptibility: Evidence from 35 case-control studies. Mol. Biol. Rep. 2012, 39, 805–816. [Google Scholar] [CrossRef]
- Zhou, D.; Mei, Q.; Luo, H.; Tang, B.; Yu, P. The polymorphisms in methylenetetrahydrofolate reductase, methionine synthase, methionine synthase reductase, and the risk of colorectal cancer. Int. J. Biol. Sci. 2012, 8, 819–830. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, J.C.; Levine, A.J.; Crott, J.W.; Baurley, J.; Haile, R.W. Folate-genetics and colorectal neoplasia: What we know and need to know next. Mol. Nutr. Food Res. 2013, 57, 607–627. [Google Scholar] [CrossRef] [PubMed]
- Pabalan, N.; Singian, E.; Tabangay, L.; Jarjanazi, H.; Singh, N. Associations of the A66G Methionine Synthase Reductase Polymorphism in Colorectal Cancer: A Systematic Review and Meta-Analysis. Biomark. Cancer 2015, 7, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Marugame, T.; Tsuji, E.; Inoue, H.; Shinomiya, S.; Kiyohara, C.; Onuma, K.; Hamada, H.; Koga, H.; Handa, K.; Hayabuchi, H.; et al. Methylenetetrahydrofolate reductase polymorphism and risk of colorectal adenomas. Cancer Lett. 2000, 151, 181–186. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, X.F.; Liu, H.X.; Hao, Y.S.; Zhao, C.L. MTHFR C677T Polymorphism and Colorectal Cancer Risk in Asians, a Meta- analysis of 21 Studies. Asian Pac. J. Cancer Prev. 2012, 13, 1203–1208. [Google Scholar] [CrossRef] [Green Version]
- Xiaolei, L.; Shunguo, Z.; Feng, Y. Rs4846049 Polymorphism at the 3’-UTR of MTHFR Gene: Association with Susceptibility to Childhood Acute Lymphoblastic Leukemia. BioMed Res. Int. 2019, 2019, 4631091. [Google Scholar] [CrossRef]
- Abbas, M.-G.; Batool, T.; Mehrnaz, N.-N.; Mehrnaz, M.; Ramin, S.; Saeedeh, S. The association of the placental MTHFR 3’-UTR polymorphisms, promoter methylation, and MTHFR expression with preeclampsia. J. Cell. Biochem. 2018, 119, 1346–1354. [Google Scholar]
- Jeon, Y.J.; Kim, J.W.; Park, H.M.; Kim, J.O.; Jang, H.G.; Oh, J.; Hwang, S.G.; Kwon, S.W.; Oh, D.; Kim, N.K. Genetic variants in 3’-UTRs of methylenetetrahydrofolate reductase (MTHFR) predict colorectal cancer susceptibility in Koreans. Sci. Rep. 2015, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu-Mei, W.; Wei-Xin, Z.; Wan-Shui, W.; Lu-Lu, S.; Dan, Y. Association between MTHFR microRNA binding site polymorphisms and methotrexate concentrations in Chinese pediatric patients with acute lymphoblastic leukemia. J. Gene Med. 2017, 19, 353–359. [Google Scholar]
- Lin, H.; Hargreaves, K.A.; Li, R.; Reiter, J.L.; Wang, Y.; Mort, M.; Cooper, D.N.; Zhou, Y.; Zhang, C.; Eadon, M.T.; et al. RegSNPs-intron: A computational framework for predicting pathogenic impact of intronic single nucleotide variants. Genome Biol. 2019, 20, 254. [Google Scholar] [CrossRef] [PubMed]
- Giovannucci, E.; Rimm, E.B.; Ascherio, A.; Stampfer, M.J.; Colditz, G.A.; Willett, W.C. Alcohol, low-methionine--low-folate diets, and risk of colon cancer in men. J. Natl. Cancer Inst. 1995, 87, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Yoshimitsu, S.; Morita, M.; Hamachi, T.; Tabata, S.; Abe, H.; Tajima, O.; Uezono, K.; Ohnaka, K.; Kono, S. Methionine synthase and thymidylate synthase gene polymorphisms and colorectal adenoma risk: The self defense forces study. Mol. Carcinog. 2012, 51, E151–E157. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | No. Patients (%) a | No. Deaths (%) a | MST (Years) | plog-rank |
|---|---|---|---|---|
| Age at diagnosis (years) b | 60.06 (±9.23) | 60.66 (±9.76) | - | - |
| Sex | 0.005 | |||
| Male | 330 (62.03) | 127 (69.40) | 6.28 | |
| Female | 202 (37.97) | 56 (30.60) | 6.53 | |
| Race | 0.009 | |||
| White | 440 (96.92) | 133 (94.33) | 6.44 | |
| Other | 14 (3.08) | 8 (5.67) | 4.66 | |
| Stage at diagnosis | <0.001 | |||
| Ⅰ | 94 (17.67) | 18 (9.84) | 6.41 | |
| Ⅱ | 209 (39.29) | 58 (31.69) | 6.64 | |
| Ⅲ | 178 (33.46) | 65 (35.52) | 6.42 | |
| Ⅳ | 51 (9.59) | 42 (22.95) | 3.90 | |
| Tumour location | 0.444 | |||
| Colon | 341 (65.83) | 110 (62.86) | 6.40 | |
| Rectum | 177 (34.17) | 65 (37.14) | 6.33 | |
| MSI status | <0.001 | |||
| MSS/MSI-L | 446 (88.49) | 168 (96.55) | 6.33 | |
| MSI-H | 58 (11.51) | 6 (3.45) | 6.67 | |
| Reported screening procedure | 0.059 | |||
| Yes | 52 (11.45) | 10 (7.09) | 6.57 | |
| No | 402 (88.55) | 131 (92.91) | 6.40 | |
| BMI (kg/m2) | 8 (1.60) | 0.097 | ||
| <18.5 | 138 (27.60) | 6 (3.57) | 4.69 | |
| 18.5–24.9 | 205 (41.00) | 41 (24.40) | 6.40 | |
| 25.0–29.9 | 149 (29.80) | 74 (44.05) | 6.40 | |
| ≥30.0 | 47 (27.98) | 6.33 | ||
| Average drinks per week | 0.062 | |||
| 0.0 | 170 (39.44) | 46 (34.59) | 6.53 | |
| 0.0–6.9 | 138 (32.02) | 43 (32.33) | 6.42 | |
| 7.0–14.0 | 74 (17.17) | 23 (17.29) | 6.44 | |
| >14.0 | 49 (11.37) | 21 (15.79) | 5.86 | |
| Smoking status | 0.133 | |||
| Yes | 375 (72.39) | 136 (77.71) | 6.36 | |
| No | 143 (27.61) | 39 (22.29) | 6.41 | |
| Red meat intake (servings/week) | 0.048 | |||
| ≤2.0 | 84 (16.47) | 25 (14.71) | 6.72 | |
| 2.1–3.0 | 257 (50.39) | 86 (50.59) | 6.42 | |
| 3.1–5.0 | 83 (16.27) | 34 (20.00) | 6.15 | |
| >5.0 | 86 (16.86) | 25 (14.71) | 6.27 | |
| Ever taken folate regularly | 0.236 | |||
| Yes | 422 (93.99) | 128 (92.09) | 6.42 | |
| No | 27 (6.01) | 11 (7.91) | 6.32 | |
| Vitamin B12 (µg/day) | 9.03 (±6.06) | 9.58 (±5.50) | - | - |
| Overall Survival HR (95% CI) a | Disease-Free Survival HR (95% CI) a | CRC-Specific Survival HR (95% CI) b | |||||||
|---|---|---|---|---|---|---|---|---|---|
| All | Colon | Rectum | All | Colon | Rectum | All | Colon | Rectum | |
| MTRR | |||||||||
| PC1 | 0.94 (0.77, 1.14) | 0.82 (0.64, 1.07) | 1.11 (0.79, 1.55) | 0.83 (0.69, 1.00) | 0.72 (0.56, 0.92) | 1.02 (0.74, 1.39) | 0.85 (0.63, 1.15) | 0.62 (0.37, 1.02) | 1.03 (0.66, 1.59) |
| PC2 | 1.00 (0.83, 1.19) | 0.91 (0.72, 1.14) | 1.20 (0.86, 1.66) | 0.96 (0.82, 1.14) | 0.93 (0.75, 1.15) | 1.07 (0.80, 1.44) | 1.01 (0.78, 1.31) | 1.19 (0.81, 1.73) | 1.05 (0.62, 1.80) |
| PC3 | 0.85 (0.70, 1.04) | 0.78 (0.61, 1.00) | 1.06 (0.72, 1.55) | 0.90 (0.75, 1.08) | 0.82 (0.66, 1.03) | 1.19 (0.85, 1.66) | 1.06 (0.84, 1.36) | 0.93 (0.67, 1.31) | 1.29 (0.81, 2.04) |
| PC4 | 0.92 (0.75, 1.11) | 0.84 (0.64, 1.09) | 1.01 (0.71, 1.44) | 0.91 (0.77, 1.09) | 0.84 (0.66, 1.07) | 0.94 (0.70, 1.27) | 0.83 (0.63, 1.10) | 0.57 (0.37, 0.87) | 1.01 (0.63, 1.62) |
| Global p | 0.414 | 0.082 | 0.794 | 0.149 | 0.015 | 0.843 | 0.533 | 0.025 | 0.863 |
| MTHFR | |||||||||
| PC1 | 1.22 (1.02, 1.46) | 1.23 (0.98, 1.55) | 1.17 (0.86, 1.59) | 1.13 (0.96, 1.33) | 1.16 (0.94, 1.43) | 1.05 (0.79, 1.40) | 1.78 (1.37, 2.31) | 2.08 (1.40, 3.08) | 1.64 (1.09, 2.48) |
| PC2 | 0.92 (0.76, 1.11) | 0.86 (0.64, 1.15) | 0.84 (0.62, 1.15) | 0.93 (0.78, 1.11) | 0.92 (0.71, 1.18) | 0.96 (0.73, 1.27) | 1.01 (0.79, 1.29) | 0.98 (0.61, 1.59) | 0.88 (0.60, 1.27) |
| PC3 | 1.11 (0.92, 1.33) | 1.08 (0.86, 1.35) | 1.32 (0.92, 1.90) | 1.09 (0.92, 1.29) | 1.08 (0.89, 1.32) | 1.13 (0.81, 1.58) | 1.21 (0.96, 1.52) | 1.41 (1.05, 1.89) | 1.08 (0.70, 1.66) |
| PC4 | 1.03 (0.88, 1.20) | 1.14 (0.93, 1.39) | 0.85 (0.62, 1.16) | 1.04 (0.90, 1.20) | 1.11 (0.91, 1.35) | 0.96 (0.74, 1.24) | 0.93 (0.75, 1.15) | 1.19 (0.89, 1.59) | 0.55 (0.31, 0.96) |
| PC5 | 0.96 (0.79, 1.16) | 0.99 (0.78, 1.26) | 0.77 (0.50, 1.19) | 1.04 (0.88, 1.24) | 1.05 (0.85, 1.30) | 0.94 (0.66, 1.35) | 1.32 (0.96, 1.81) | 1.60 (1.04, 2.47) | 0.93 (0.49, 1.78) |
| Global p | 0.226 | 0.233 | 0.291 | 0.511 | 0.493 | 0.976 | 0.0005 | 0.0004 | 0.051 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Du, M.; Vallis, J.; Shariati, M.; Parfrey, P.S.; Mclaughlin, J.R.; Wang, P.P.; Zhu, Y. The Roles of MTRR and MTHFR Gene Polymorphisms in Colorectal Cancer Survival. Nutrients 2022, 14, 4594. https://doi.org/10.3390/nu14214594
Wang Y, Du M, Vallis J, Shariati M, Parfrey PS, Mclaughlin JR, Wang PP, Zhu Y. The Roles of MTRR and MTHFR Gene Polymorphisms in Colorectal Cancer Survival. Nutrients. 2022; 14(21):4594. https://doi.org/10.3390/nu14214594
Chicago/Turabian StyleWang, Yu, Meizhi Du, Jillian Vallis, Matin Shariati, Patrick S. Parfrey, John R. Mclaughlin, Peizhong Peter Wang, and Yun Zhu. 2022. "The Roles of MTRR and MTHFR Gene Polymorphisms in Colorectal Cancer Survival" Nutrients 14, no. 21: 4594. https://doi.org/10.3390/nu14214594
APA StyleWang, Y., Du, M., Vallis, J., Shariati, M., Parfrey, P. S., Mclaughlin, J. R., Wang, P. P., & Zhu, Y. (2022). The Roles of MTRR and MTHFR Gene Polymorphisms in Colorectal Cancer Survival. Nutrients, 14(21), 4594. https://doi.org/10.3390/nu14214594