
Exploration of the Gut–Brain Axis through Metabolomics Identifies Serum Propionic Acid Associated with Higher Cognitive Decline in Older Persons
 ,
,  , , , , , , , , , ,
, , , , , , , , , ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Nested Case–Control Samples
2.3. Metabolomics Analysis of Serum Samples
2.4. Other Variables
2.5. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fan, Y.; Pedersen, O. Gut Microbiota in Human Metabolic Health and Disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Tengeler, A.C.; Kozicz, T.; Kiliaan, A.J. Relationship between Diet, the Gut Microbiota, and Brain Function. Nutr. Rev. 2018, 76, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut Microbiota Functions: Metabolism of Nutrients and Other Food Components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Xu, R.; Wang, Q. Towards Understanding Brain-Gut-Microbiome Connections in Alzheimer’s Disease. BMC Syst. Biol. 2016, 10, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saji, N.; Murotani, K.; Hisada, T.; Kunihiro, T.; Tsuduki, T.; Sugimoto, T.; Kimura, A.; Niida, S.; Toba, K.; Sakurai, T. Relationship between Dementia and Gut Microbiome-Associated Metabolites: A Cross-Sectional Study in Japan. Sci. Rep. 2020, 10, 8088. [Google Scholar] [CrossRef] [PubMed]
- Szablewski, L. Human Gut Microbiota in Health and Alzheimer’s Disease. JAD 2018, 62, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, C.; Santangelo, R. Alzheimer’s Disease and Gut Microbiota Modifications: The Long Way between Preclinical Studies and Clinical Evidence. Pharmacol. Res. 2018, 129, 329–336. [Google Scholar] [CrossRef]
- Chouraki, V.; Preis, S.R.; Yang, Q.; Beiser, A.; Li, S.; Larson, M.G.; Weinstein, G.; Wang, T.J.; Gerszten, R.E.; Vasan, R.S.; et al. Association of Amine Biomarkers with Incident Dementia and Alzheimer’s Disease in the Framingham Study. Alzheimers Dement. 2017, 13, 1327–1336. [Google Scholar] [CrossRef]
- Gulaj, E.; Pawlak, K.; Bien, B.; Pawlak, D. Kynurenine and Its Metabolites in Alzheimer’s Disease Patients. Adv. Med. Sci. 2010, 55, 204–211. [Google Scholar] [CrossRef]
- Vogt, N.M.; Romano, K.A.; Darst, B.F.; Engelman, C.D.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Blennow, K.; Zetterberg, H.; Bendlin, B.B.; et al. The Gut Microbiota-Derived Metabolite Trimethylamine N-Oxide Is Elevated in Alzheimer’s Disease. Alz. Res. Ther. 2018, 10, 124. [Google Scholar] [CrossRef] [Green Version]
- MahmoudianDehkordi, S.; Arnold, M.; Nho, K.; Ahmad, S.; Jia, W.; Xie, G.; Louie, G.; Kueider-Paisley, A.; Moseley, M.A.; Thompson, J.W.; et al. Altered Bile Acid Profile Associates with Cognitive Impairment in Alzheimer’s Disease-An Emerging Role for Gut Microbiome. Alzheimers Dement. 2019, 15, 76–92. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, R.; Bouzari, B.; Hosseini-Fard, S.R.; Mazaheri, M.; Ahmadyousefi, Y.; Abdi, M.; Jalalifar, S.; Karimitabar, Z.; Teimoori, A.; Keyvani, H.; et al. Role of Microbiota-Derived Short-Chain Fatty Acids in Nervous System Disorders. Biomed. Pharmacother. 2021, 139, 111661. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Meng, L.; Shen, L. Multiple Roles of Short-Chain Fatty Acids in Alzheimer Disease. Nutrition 2022, 93, 111499. [Google Scholar] [CrossRef] [PubMed]
- Collino, S.; Martin, F.-P.J.; Rezzi, S. Clinical Metabolomics Paves the Way towards Future Healthcare Strategies: Clinical Metabolomics. Br. J. Clin. Pharmacol. 2013, 75, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.A. The Tantalizing Links between Gut Microbes and the Brain. Nature 2015, 526, 312–314. [Google Scholar] [CrossRef] [Green Version]
- Ou, F.-S.; Michiels, S.; Shyr, Y.; Adjei, A.A.; Oberg, A.L. Biomarker Discovery and Validation: Statistical Considerations. JTO 2021, 16, 537–545. [Google Scholar] [CrossRef]
- Antoniak, M.; Pugliatti, M.; Hubbard, R.; Britton, J.; Sotgiu, S.; Sadovnick, A.D.; Yee, I.M.; Cumsille, M.A.; Bevilacqua, J.A.; Burdett, S.; et al. Vascular Factors and Risk of Dementia: Design of the Three-City Study and Baseline Characteristics of the Study Population. Neuroepidemiology 2003, 22, 316–325. [Google Scholar] [CrossRef]
- Féart, C. Adherence to a Mediterranean Diet, Cognitive Decline, and Risk of Dementia. JAMA 2009, 302, 638. [Google Scholar] [CrossRef] [Green Version]
- Féart, C.; Jutand, M.A.; Larrieu, S.; Letenneur, L.; Delcourt, C.; Combe, N.; Barberger-Gateau, P. Energy, Macronutrient and Fatty Acid Intake of French Elderly Community Dwellers and Association with Socio-Demographic Characteristics: Data from the Bordeaux Sample of the Three-City Study. Br. J. Nutr. 2007, 98, 1046–1057. [Google Scholar] [CrossRef]
- Low, D.Y.; Lefèvre-Arbogast, S.; González-Domínguez, R.; Urpi-Sarda, M.; Micheau, P.; Petera, M.; Centeno, D.; Durand, S.; Pujos-Guillot, E.; Korosi, A.; et al. Diet-Related Metabolites Associated with Cognitive Decline Revealed by Untargeted Metabolomics in a Prospective Cohort. Mol. Nutr. Food Res. 2019, 63, 1900177. [Google Scholar] [CrossRef] [Green Version]
- Folstein, M.F.; Folstein, S.E.; McHugh, P.R. “Mini-Mental State”. A Practical Method for Grading the Cognitive State of Patients for the Clinician. J. Psychiatr. Res. 1975, 12, 189–198. [Google Scholar] [CrossRef]
- Benton, A.L. Manuel du Test de Rétention Visuelle: Applications Cliniques et Expérimentales; Editions du Centre de Psychologie Appliquée; Centre de Psychologie Appliquée: Paris, France, 1953. [Google Scholar]
- Isaacs, B.; Kennie, A.T. The Set Test as an Aid to the Detection of Dementia in Old People. Br. J. Psychiatry 1973, 123, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Reitan, R.M. Validity of the Trail Making Test as an Indicator of Organic Brain Damage. Percept. Mot. Ski. 1958, 8, 271–276. [Google Scholar] [CrossRef]
- González-Domínguez, R.; Jáuregui, O.; Queipo-Ortuño, M.I.; Andrés-Lacueva, C. Characterization of the Human Exposome by a Comprehensive and Quantitative Large-Scale Multianalyte Metabolomics Platform. Anal. Chem. 2020, 92, 13767–13775. [Google Scholar] [CrossRef]
- Jafari, M.; Ansari-Pour, N. Why, When and How to Adjust Your p Values? Cell J. 2018, 20, 604–607. [Google Scholar] [CrossRef]
- Royston, P.; Ambler, G.; Sauerbrei, W. The Use of Fractional Polynomials to Model Continuous Risk Variables in Epidemiology. Int. J. Epidemiol. 1999, 28, 964–974. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.M.; Cologne, J.B.; Jang, E.; Lange, T.; Tatsukawa, Y.; Ohishi, W.; Utada, M.; Cullings, H.M. Causal Mediation Analysis in Nested Case-control Studies Using Conditional Logistic Regression. Biom. J. 2020, 62, 1939–1959. [Google Scholar] [CrossRef]
- VanderWeele, T.J. Mediation Analysis: A Practitioner’s Guide. Annu. Rev. Public Health 2016, 37, 17–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirosh, A.; Calay, E.S.; Tuncman, G.; Claiborn, K.C.; Inouye, K.E.; Eguchi, K.; Alcala, M.; Rathaus, M.; Hollander, K.S.; Ron, I.; et al. The Short-Chain Fatty Acid Propionate Increases Glucagon and FABP4 Production, Impairing Insulin Action in Mice and Humans. Sci. Transl. Med. 2019, 11, eaav0120. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Xu, H.; Tu, X.; Gao, Z. The Role of Short-Chain Fatty Acids of Gut Microbiota Origin in Hypertension. Front. Microbiol. 2021, 12, 730809. [Google Scholar] [CrossRef] [PubMed]
- Demigné, C.; Morand, C.; Levrat, M.-A.; Besson, C.; Moundras, C.; Rémésy, C. Effect of Propionate on Fatty Acid and Cholesterol Synthesis and on Acetate Metabolism in Isolated Rat Hepatocytes. Br. J. Nutr. 1995, 74, 209–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, S.; Rakugi, H.; Morishita, R. Roles of Vascular Risk Factors in the Pathogenesis of Dementia. Hypertens. Res. 2020, 43, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic Distribution of Three Pathways for Propionate Production within the Human Gut Microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, D.J.; Preston, T. Formation of Short Chain Fatty Acids by the Gut Microbiota and Their Impact on Human Metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani-López, E.; García, H.S.; López-Malo, A. Organic Acids as Antimicrobials to Control Salmonella in Meat and Poultry Products. Food Res. Int. 2012, 45, 713–721. [Google Scholar] [CrossRef]
- Reis, J.A.; Paula, A.T.; Casarotti, S.N.; Penna, A.L.B. Lactic Acid Bacteria Antimicrobial Compounds: Characteristics and Applications. Food Eng. Rev. 2012, 4, 124–140. [Google Scholar] [CrossRef]
- Schönfeld, P.; Wojtczak, L. Short- and Medium-Chain Fatty Acids in Energy Metabolism: The Cellular Perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef] [Green Version]
- Chambers, E.S.; Byrne, C.S.; Aspey, K.; Chen, Y.; Khan, S.; Morrison, D.J.; Frost, G. Acute Oral Sodium Propionate Supplementation Raises Resting Energy Expenditure and Lipid Oxidation in Fasted Humans. Diabetes Obes. Metab. 2018, 20, 1034–1039. [Google Scholar] [CrossRef]
- Topping, D.L.; Clifton, P.M. Short-Chain Fatty Acids and Human Colonic Function: Roles of Resistant Starch and Nonstarch Polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killingsworth, J.; Sawmiller, D.; Shytle, R.D. Propionate and Alzheimer’s Disease. Front. Aging Neurosci. 2021, 12, 580001. [Google Scholar] [CrossRef] [PubMed]
- Ticinesi, A.; Nouvenne, A.; Tana, C.; Prati, B.; Meschi, T. Gut Microbiota and Microbiota-Related Metabolites as Possible Biomarkers of Cognitive Aging. In Reviews on Biomarker Studies in Aging and Anti-Aging Research; Advances in Experimental Medicine and Biology; Guest, P.C., Ed.; Springer International Publishing: Cham, Denmark, 2019; Volume 1178, pp. 129–154. ISBN 978-3-030-25649-4. [Google Scholar]
- Mitsui, R.; Ono, S.; Karaki, S.; Kuwahara, A. Propionate Modulates Spontaneous Contractions via Enteric Nerves and Prostaglandin Release in the Rat Distal Colon. JJP 2005, 55, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Sunkara, L.T.; Jiang, W.; Zhang, G. Modulation of Antimicrobial Host Defense Peptide Gene Expression by Free Fatty Acids. PLoS ONE 2012, 7, e49558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ringer, A.I. The Chemistry of Gluconeogenesis. J. Biol. Chem. 1912, 12, 511–515. [Google Scholar] [CrossRef]
- Hoyles, L.; Snelling, T.; Umlai, U.-K.; Nicholson, J.K.; Carding, S.R.; Glen, R.C.; McArthur, S. Microbiome–Host Systems Interactions: Protective Effects of Propionate upon the Blood–Brain Barrier. Microbiome 2018, 6, 55. [Google Scholar] [CrossRef] [Green Version]
- Sadler, R.; Cramer, J.V.; Heindl, S.; Kostidis, S.; Betz, D.; Zuurbier, K.R.; Northoff, B.H.; Heijink, M.; Goldberg, M.P.; Plautz, E.J.; et al. Short-Chain Fatty Acids Improve Poststroke Recovery via Immunological Mechanisms. J. Neurosci. 2020, 40, 1162–1173. [Google Scholar] [CrossRef]
- Erny, D.; Hrabě de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host Microbiota Constantly Control Maturation and Function of Microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Ho, L.; Ono, K.; Tsuji, M.; Mazzola, P.; Singh, R.; Pasinetti, G.M. Protective Roles of Intestinal Microbiota Derived Short Chain Fatty Acids in Alzheimer’s Disease-Type Beta-Amyloid Neuropathological Mechanisms. Expert. Rev. Neurother. 2018, 18, 83–90. [Google Scholar] [CrossRef] [Green Version]
- Colombo, A.V.; Sadler, R.K.; Llovera, G.; Singh, V.; Roth, S.; Heindl, S.; Sebastian Monasor, L.; Verhoeven, A.; Peters, F.; Parhizkar, S.; et al. Microbiota-Derived Short Chain Fatty Acids Modulate Microglia and Promote Aβ Plaque Deposition. eLife 2021, 10, e59826. [Google Scholar] [CrossRef]
- Marizzoni, M.; Cattaneo, A.; Mirabelli, P.; Festari, C.; Lopizzo, N.; Nicolosi, V.; Mombelli, E.; Mazzelli, M.; Luongo, D.; Naviglio, D.; et al. Short-Chain Fatty Acids and Lipopolysaccharide as Mediators Between Gut Dysbiosis and Amyloid Pathology in Alzheimer’s Disease. JAD 2020, 78, 683–697. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, A.; Geddes, T.; Han, B.; Bahado-Singh, R.O.; Wilson, G.D.; Imam, K.; Maddens, M.; Graham, S.F. Diagnostic Biomarkers of Alzheimer’s Disease as Identified in Saliva Using 1H NMR-Based Metabolomics. JAD 2017, 58, 355–359. [Google Scholar] [CrossRef] [PubMed]
- Figueira, J.; Jonsson, P.; Nordin Adolfsson, A.; Adolfsson, R.; Nyberg, L.; Öhman, A. NMR Analysis of the Human Saliva Metabolome Distinguishes Dementia Patients from Matched Controls. Mol. BioSyst. 2016, 12, 2562–2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, N.M.; Kerby, R.L.; Dill-McFarland, K.A.; Harding, S.J.; Merluzzi, A.P.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Zetterberg, H.; Blennow, K.; et al. Gut Microbiome Alterations in Alzheimer’s Disease. Sci. Rep. 2017, 7, 13537. [Google Scholar] [CrossRef] [Green Version]
- Watson, G.S.; Craft, S. The Role of Insulin Resistance in the Pathogenesis of Alzheimer’s Disease: Implications for Treatment. CNS Drugs 2003, 17, 27–45. [Google Scholar] [CrossRef]
- Adler, G.K.; Hornik, E.S.; Murray, G.; Bhandari, S.; Yadav, Y.; Heydarpour, M.; Basu, R.; Garg, R.; Tirosh, A. Acute Effects of the Food Preservative Propionic Acid on Glucose Metabolism in Humans. BMJ Open Diab. Res. Care 2021, 9, e002336. [Google Scholar] [CrossRef]
- US Department of Agriculture Propionic Acid. Available online: https://www.ams.usda.gov/sites/default/files/media/Prop%20acid%20report.pdf (accessed on 4 October 2021).
- Więckowska-Gacek, A.; Mietelska-Porowska, A.; Wydrych, M.; Wojda, U. Western Diet as a Trigger of Alzheimer’s Disease: From Metabolic Syndrome and Systemic Inflammation to Neuroinflammation and Neurodegeneration. Ageing Res. Rev. 2021, 70, 101397. [Google Scholar] [CrossRef]
- Zinöcker, M.; Lindseth, I. The Western Diet–Microbiome-Host Interaction and Its Role in Metabolic Disease. Nutrients 2018, 10, 365. [Google Scholar] [CrossRef] [Green Version]
- Spagnuolo, M.S.; Pallottini, V.; Mazzoli, A.; Iannotta, L.; Tonini, C.; Morone, B.; Ståhlman, M.; Crescenzo, R.; Strazzullo, M.; Iossa, S.; et al. A Short-Term Western Diet Impairs Cholesterol Homeostasis and Key Players of Beta Amyloid Metabolism in Brain of Middle Aged Rats. Mol. Nutr. Food Res. 2020, 64, 2000541. [Google Scholar] [CrossRef]
- Hakvoort, T.B.M.; He, Y.; Kulik, W.; Vermeulen, J.L.M.; Duijst, S.; Ruijter, J.M.; Runge, J.H.; Deutz, N.E.P.; Koehler, S.E.; Lamers, W.H. Pivotal Role of Glutamine Synthetase in Ammonia Detoxification. Hepatology 2017, 65, 281–293. [Google Scholar] [CrossRef]
- El-Ansary, A.; Shaker, G.H.; El-Gezeery, A.R.; Al-Ayadhi, L. The Neurotoxic Effect of Clindamycin—Induced Gut Bacterial Imbalance and Orally Administered Propionic Acid on DNA Damage Assessed by the Comet Assay: Protective Potency of Carnosine and Carnitine. Gut Pathog. 2013, 5, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seiler, N. Ammonia and Alzheimer’s Disease. Neurochem. Int. 2002, 41, 189–207. [Google Scholar] [CrossRef]
- Wang, R.; Reddy, P.H. Role of Glutamate and NMDA Receptors in Alzheimer’s Disease. JAD 2017, 57, 1041–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martos, D.; Tuka, B.; Tanaka, M.; Vécsei, L.; Telegdy, G. Memory Enhancement with Kynurenic Acid and Its Mechanisms in Neurotransmission. Biomedicines 2022, 10, 849. [Google Scholar] [CrossRef]
- Hawkins, R.A. The Blood-Brain Barrier and Glutamate. AJCN 2009, 90, 867S–874S. [Google Scholar] [CrossRef]


{kind=link}
| Discovery (n = 418) | Validation (n = 420) | |||
|---|---|---|---|---|
| Cases | Controls | Cases | Controls | |
| Matching variables | ||||
| Age (years), mean (SD) | 75.9 (4.4) | 75.7 (4.2) | 76.5 (5.2) | 76.1 (4.7) |
| Women | 138 (66) | 138 (66) | 133 (63) | 133 (63) |
| Level of education above secondary level | 149 (71) | 149 (71) | 150 (71) | 150 (71) |
| Baseline characteristics | ||||
| BMI (kg/m2), mean (SD) | 26.8 (4.3) | 26.1 (3.6) | 25.7 (4.5) | 25.0 (3.6) |
| Alcohol consumption (g per day), mean (SD) | 13.0 (14.6) | 14.6 (17.2) | 12.3 (14.5) | 12.4 (12.7) |
| Smoking (pack-years), mean (SD) | 9.1 (19.7) | 7.3 (14.6) | 8.1 (19.2) | 6.0 (12.8) |
| High blood pressure | 164 (78) | 159 (76) | 176 (84) | 174 (83) |
| Hypercholesterolemia | 79 (38) | 93 (44) | 85 (40) | 80 (38) |
| Diabetes | 27 (13) * | 12 (6) * | 27 (13) * | 12 (6) * |
| Number of medications, mean (SD) | 4.9 (2.7) * | 4.1 (2.4) * | 5.5 (3.0) * | 4.0 (2.9) * |
| Discovery (n = 418) | Validation (n = 420) | |||||
|---|---|---|---|---|---|---|
| Metabolite | OR 2 | 95% CI | FDR-Adjusted p Value 3 | OR | 95% CI | p Value |
| Phenylalanine | 0.93 | 0.76; 1.14 | 0.83 | |||
| Tyrosine | 1.18 | 0.97; 1.44 | 0.37 | |||
| Tryptophan | 1.11 | 0.92; 1.35 | 0.70 | |||
| Phenyl-lactic acid | 1.26 | 1.02; 1.57 | 0.26 | |||
| p-HPLA | - | - | 0.33 | |||
| Phenylacetylglutamine | 1.34 | 1.08; 1.66 | 0.09 * | 1.14 | 0.94; 1.39 | 0.19 |
| Epinephrine | - | - | 0.84 | |||
| p-Cresol-G | 1.13 | 0.93; 1.36 | 0.64 | |||
| p-Cresol-S | 1.13 | 0.93; 1.37 | 0.68 | |||
| Indoxyl-S | - | - | 0.80 | |||
| Serotonin | 0.96 | 0.79; 1.15 | 0.87 | |||
| Indolelactic acid | 1.38 | 1.11; 1.72 | 0.07* | 0.93 | 0.77; 1.12 | 0.46 |
| Indoleacetic acid | 1.12 | 0.91; 1.37 | 0.70 | |||
| 5-HIAA | - | - | 0.84 | |||
| Indolepropionic acid | 1.00 | 0.84; 1.20 | 1.00 | |||
| Kynurenine | 1.17 | 0.95; 1.43 | 0.49 | |||
| Kynurenic acid | 1.34 | 1.07; 1.67 | 0.10 * | 1.07 | 0.88; 1.29 | 0.49 |
| Xanthurenic acid | 1.12 | 0.92; 1.37 | 0.68 | |||
| Anthranilic acid | 0.97 | 0.80; 1.18 | 0.94 | |||
| Picolinic acid | 0.93 | 0.77; 1.12 | 0.80 | |||
| Ergothioneine | 0.98 | 0.81; 1.18 | 0.94 | |||
| Lactic acid | 1.10 | 0.91; 1.34 | 0.76 | |||
| Choline | 1.19 | 0.97; 1.45 | 0.37 | |||
| TMAO | 1.03 | 0.84; 1.26 | 0.94 | |||
| Betaine | 0.73 | 0.60; 0.88 | 0.04 * | 0.97 | 0.8; 1.17 | 0.73 |
| Carnitine | 1.07 | 0.88; 1.31 | 0.83 | |||
| GDCA | 1.24 | 1.01; 1.52 | 0.26 | |||
| Thiamine | - | - | 0.92 | |||
| Riboflavin | 0.87 | 0.71; 1.07 | 0.59 | |||
| Niacinamide | 1.26 | 1.03; 1.55 | 0.21 | |||
| Pantothenic acid | 1.43 | 1.15; 1.77 | 0.04 * | 1.04 | 0.86; 1.24 | 0.70 |
| 4-pyridoxic acid | - | - | 0.94 | |||
| Biotin | 1.38 | 1.03; 1.86 | 0.26 | |||
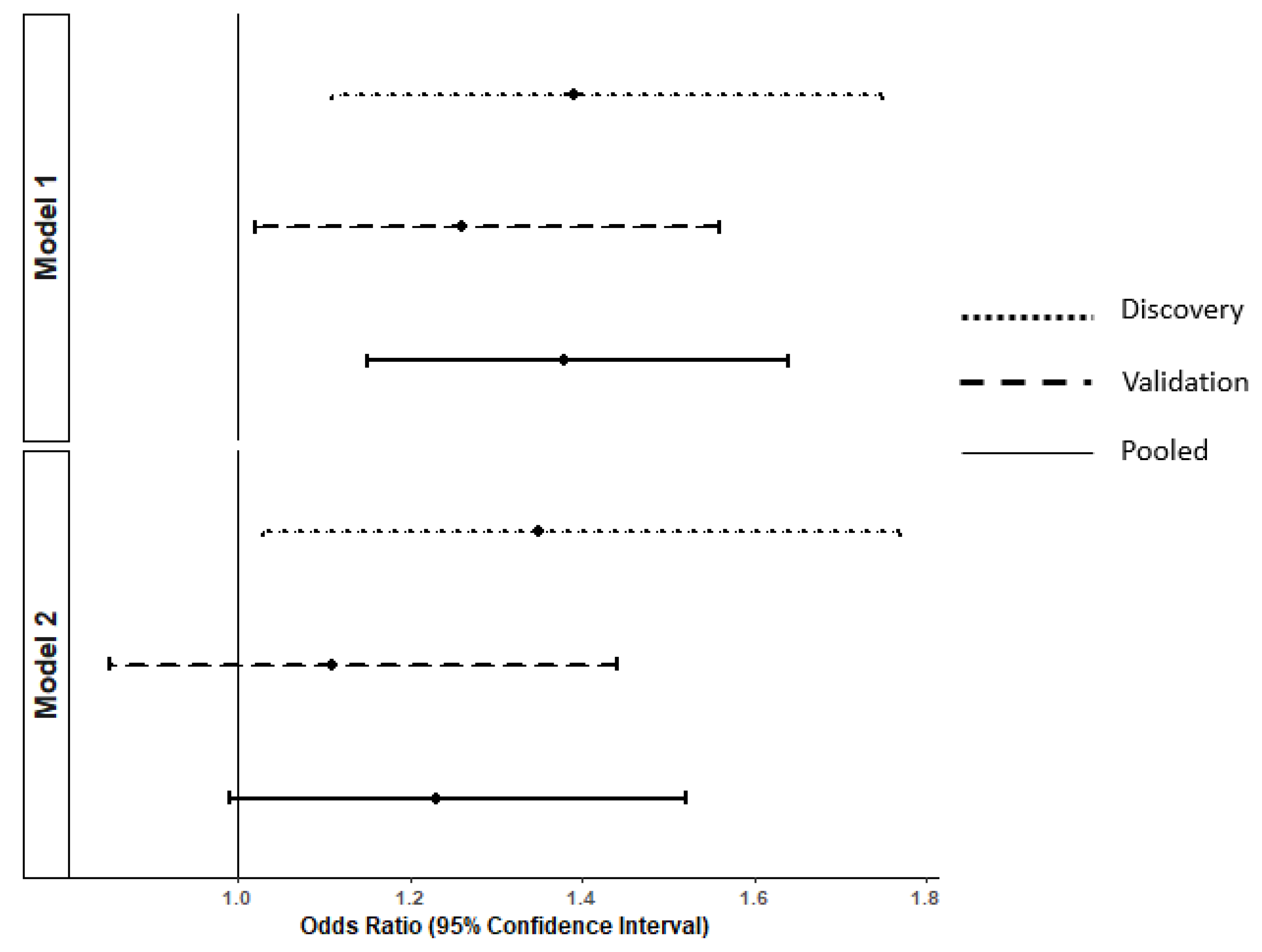
| Propionic acid | 1.40 | 1.11; 1.75 | 0.07 * | 1.26 | 1.02; 1.55 | 0.03 |
| Butyric acid | 1.08 | 0.90; 1.30 | 0.80 | |||
| Valeric acid | 0.93 | 0.76; 1.13 | 0.80 | |||
| 2-HBA | 1.10 | 0.87; 1.38 | 0.80 | |||
| 3-HBA-S | 0.99 | 0.82; 1.19 | 0.99 | |||
| 4-HBA-S | 1.06 | 0.87; 1.29 | 0.84 | |||
| 2,6-DHBA | 1.06 | 0.88; 1.29 | 0.84 | |||
| 3,4-DHBA | 0.95 | 0.77; 1.16 | 0.87 | |||
| HA | 0.82 | 0.67; 1.02 | 0.33 | |||
| 4-HHA | 0.95 | 0.78; 1.16 | 0.87 | |||
| 3-HHA | 0.98 | 0.79; 1.20 | 0.94 | |||
| iVA | 1.01 | 0.83; 1.22 | 0.99 | |||
| 2-HPAA | 1.08 | 0.89; 1.31 | 0.80 | |||
| 4-HPAA-G | 1.18 | 0.94; 1.47 | 0.50 | |||
| 3-HPAA-S | 1.00 | 0.82; 1.20 | 0.99 | |||
| 3,4-DHPAA-S | 1.00 | 0.82; 1.21 | 0.99 | |||
| FA-S | 0.95 | 0.78; 1.16 | 0.87 | |||
| 3-HPPA | 0.96 | 0.78; 1.17 | 0.89 | |||
| HPPA-S | 0.91 | 0.74; 1.13 | 0.80 | |||
| 3,5-DHPPA-S | 1.02 | 0.85; 1.23 | 0.94 | |||
| DHCA-3S | 1.03 | 0.84; 1.25 | 0.94 | |||
| DHFA | 1.11 | 0.91; 1.35 | 0.76 | |||
| DHFA-S | 0.89 | 0.73; 1.08 | 0.68 | |||
| DHiFA-S | 1.00 | 0.83; 1.22 | 0.99 | |||
| 3-HPHPA | 1.01 | 0.83; 1.23 | 0.99 | |||
| PYR-S | 0.99 | 0.82; 1.20 | 0.99 | |||
| MePYR-S | 0.97 | 0.80; 1.18 | 0.94 | |||
| CAT-S | - | - | 0.80 | |||
| 4-MeCAT-S | 0.84 | 0.66; 1.05 | 0.47 | |||
| VAN | 0.82 | 0.66; 1.02 | 0.33 | |||
| 3′,4′-DHPV-S | 0.70 | 0.54; 0.91 | 0.09 * | 1.04 | 0.86; 1.25 | 0.69 |
| MHPV-S | 0.83 | 0.68; 1.02 | 0.33 | |||
| UroA-G | 0.92 | 0.75; 1.14 | 0.80 | |||
| UroA-S | 0.89 | 0.71; 1.11 | 0.71 | |||
| UroB-G | 1.04 | 0.86; 1.27 | 0.89 | |||
| UroB-S | 1.05 | 0.86; 1.28 | 0.87 | |||
| DHRSV-S | 0.98 | 0.80; 1.19 | 0.94 | |||
| EL | 0.87 | 0.71; 1.06 | 0.51 | |||
| EL-S | 0.83 | 0.69; 1.01 | 0.33 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neuffer, J.; González-Domínguez, R.; Lefèvre-Arbogast, S.; Low, D.Y.; Driollet, B.; Helmer, C.; Du Preez, A.; de Lucia, C.; Ruigrok, S.R.; Altendorfer, B.; et al. Exploration of the Gut–Brain Axis through Metabolomics Identifies Serum Propionic Acid Associated with Higher Cognitive Decline in Older Persons. Nutrients 2022, 14, 4688. https://doi.org/10.3390/nu14214688
Neuffer J, González-Domínguez R, Lefèvre-Arbogast S, Low DY, Driollet B, Helmer C, Du Preez A, de Lucia C, Ruigrok SR, Altendorfer B, et al. Exploration of the Gut–Brain Axis through Metabolomics Identifies Serum Propionic Acid Associated with Higher Cognitive Decline in Older Persons. Nutrients. 2022; 14(21):4688. https://doi.org/10.3390/nu14214688
Chicago/Turabian StyleNeuffer, Jeanne, Raúl González-Domínguez, Sophie Lefèvre-Arbogast, Dorrain Y. Low, Bénédicte Driollet, Catherine Helmer, Andrea Du Preez, Chiara de Lucia, Silvie R. Ruigrok, Barbara Altendorfer, and et al. 2022. "Exploration of the Gut–Brain Axis through Metabolomics Identifies Serum Propionic Acid Associated with Higher Cognitive Decline in Older Persons" Nutrients 14, no. 21: 4688. https://doi.org/10.3390/nu14214688
APA StyleNeuffer, J., González-Domínguez, R., Lefèvre-Arbogast, S., Low, D. Y., Driollet, B., Helmer, C., Du Preez, A., de Lucia, C., Ruigrok, S. R., Altendorfer, B., Aigner, L., Lucassen, P. J., Korosi, A., Thuret, S., Manach, C., Pallàs, M., Urpi-Sardà, M., Sánchez-Pla, A., Andres-Lacueva, C., & Samieri, C. (2022). Exploration of the Gut–Brain Axis through Metabolomics Identifies Serum Propionic Acid Associated with Higher Cognitive Decline in Older Persons. Nutrients, 14(21), 4688. https://doi.org/10.3390/nu14214688