
Puerarin Reduces Oxidative Damage and Photoaging Caused by UVA Radiation in Human Fibroblasts by Regulating Nrf2 and MAPK Signaling Pathways

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Cell Culture
2.3. Measurement of Cytotoxicity
2.4. UVA Irradiation Procedure
2.5. Protective Effects of Puerarin on UVA-Induced Oxidative Stress
2.6. Intracellular ROS Content Measurement
2.7. Measurement of Intracellular Total Antioxidant Capacity (ABTS Method), MDA Content, and Antioxidant Enzyme Activity
2.8. ELISA
2.9. RT-PCR
2.10. Experiments on Animals
2.10.1. Experimental Animals and Groups
2.10.2. Moisturizing Experiment
2.10.3. Skin Tissue Total Antioxidant Capacity and CAT Activity Measurement
2.10.4. Hematoxylin-Eosin (HE) Dye
2.11. Statistical Analysis
3. Results
3.1. Effects of Puerarin on HSF
3.2. Effects of Puerarin on Intracellular Antioxidant Capacity
3.3. Effects of Puerarin on UVA-Induced Cell Senescence
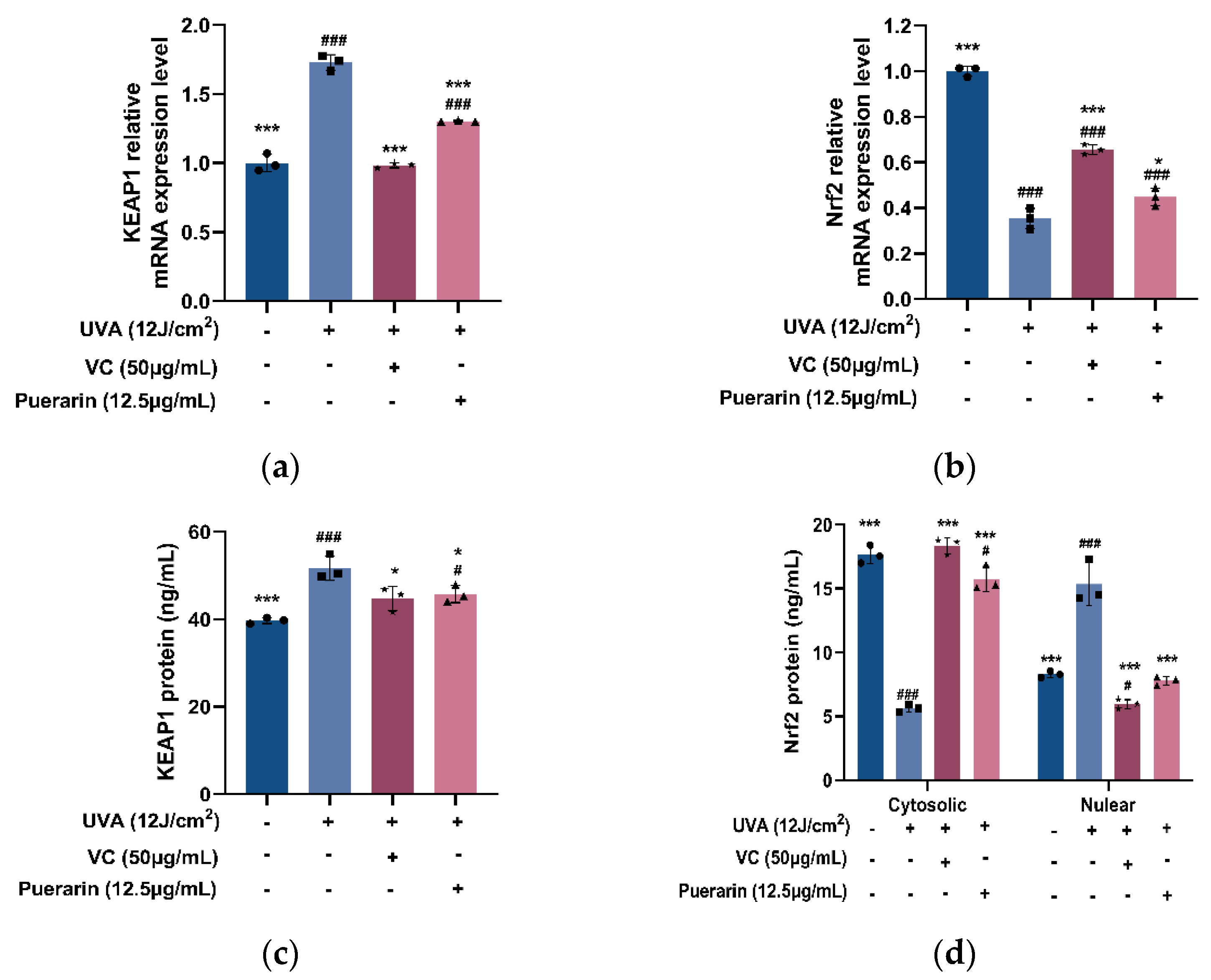
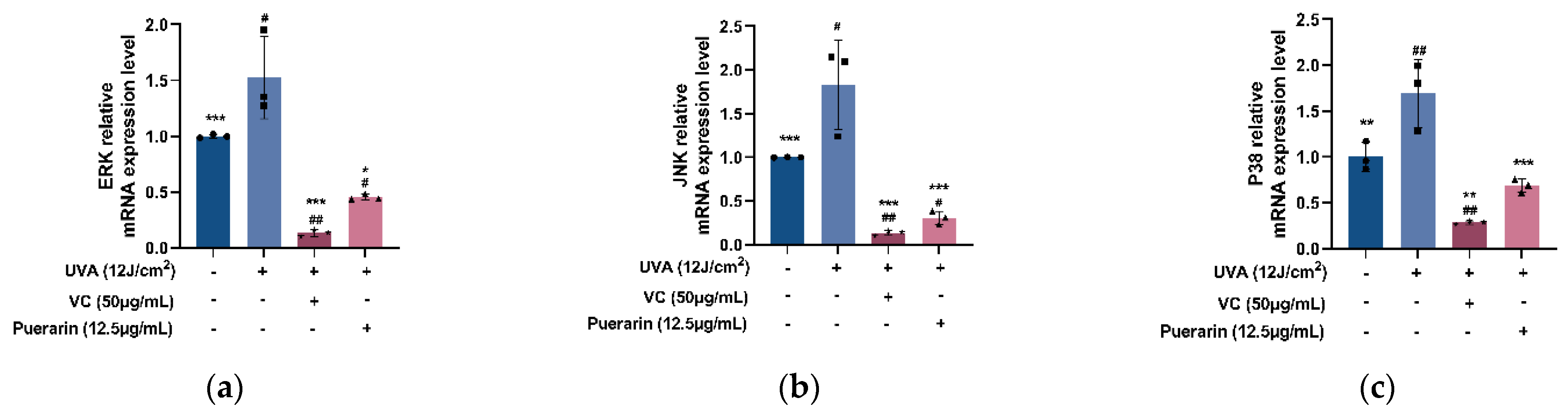
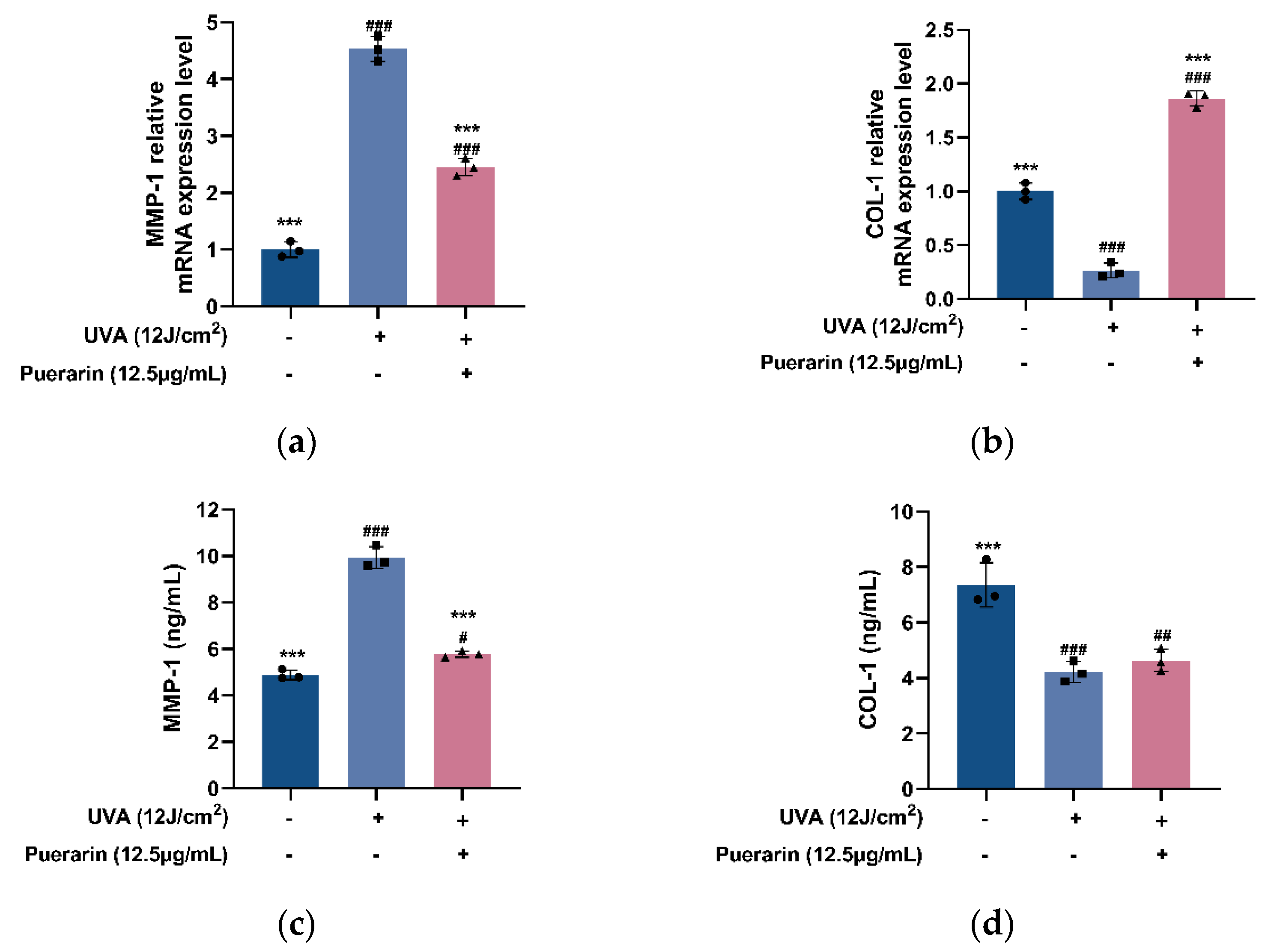
3.4. Protective Mechanism of Puerarin against Oxidative Stress in Mice
3.4.1. Blood Routine Test Results
3.4.2. Moisturizing Experiment
3.4.3. Total Antioxidant Capacity and CAT Enzyme Activity in Mice Tissue
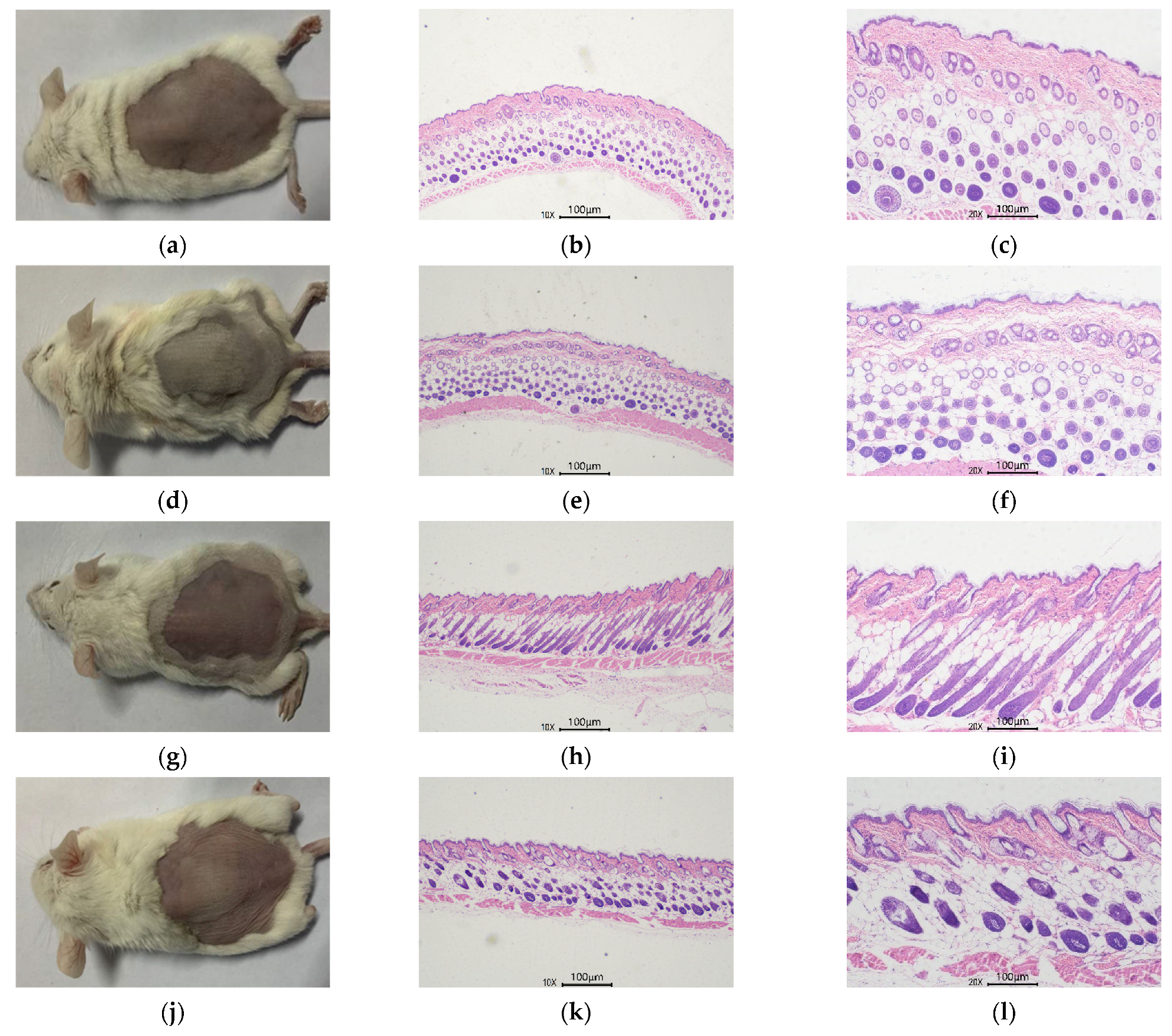
3.4.4. Skin Histological Observation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blank Group | Model Group | VC Group | Puerarin Group | |
|---|---|---|---|---|
| WBC (109/L) | 7.53 ± 1.11 | 7.55 ± 0.89 | 4.17 ± 0.42 | 7.5 ± 2.26 |
| RBC (1012/L) | 10.1 ± 1.18 | 9.85 ± 0.42 | 9.6 ± 0.26 | 10.93 ± 0.97 |
| Hb (g/L) | 162 ± 8.54 | 155.25 ± 9.91 | 158.67 ± 7.77 | 175.5 ± 16.11 |
| HCT | 0.50 ± 0.03 | 0.47 ± 0.04 | 0.48 ± 0.03 | 0.54 ± 0.05 |
| MCV (fL) | 52.33 ± 1.53 | 47.75 ± 2.36 | 51.5 ± 1.53 | 49.25 ± 1.71 |
| MCH (Pg) | 16.33 ± 1.15 | 15.75 ± 0.5 | 16.67 ± 0.58 | 16 ± 0.82 |
| MCHC (g/L) | 323.67 ± 2.08 | 328.25 ± 10.01 | 328 ± 8.72 | 326 ± 2.94 |
| PLT (109/L) | 1039.33 ± 218.48 | 744.25 ± 112 | 689.33 ± 165.03 | 835.75 ± 235.36 |
| N# (109/L) | 2.33 ± 0.97 | 2.83 ± 1.54 | 1.55 ± 0.36 | 2.8 ± 1.67 |
| L# (109/L) | 5.75 ± 0.85 | 5.65 ± 0.05 | 2.45 ± 0.4 | 4.77 ± 1.78 |
References
- Xuan, S.H.; Lee, N.H.; Park, S.N. Atractyligenin, a terpenoid isolated from coffee silverskin, inhibits cutaneous photoaging. J. Photochem. Photobiol. B Biol. 2019, 194, 166–173. [Google Scholar] [CrossRef]
- Kato, S.; Matsui, H.; Saitoh, Y.; Miwa, N. Fish collagen-containing drink is subcutaneously absorbed and attenuates the UVA-induced tissue-integrity destruction and DNA damages in 3D-human skin tissue model. J. Funct. Foods 2011, 3, 50–55. [Google Scholar] [CrossRef]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1–Nrf2 system. Free Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Sykiotis, G.P.; Habeos, I.G.; Samuelson, A.V.; Bohmann, D. The role of the antioxidant and longevity-promoting Nrf2 pathway in metabolic regulation. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.P.; Lian, W.Y.; Chen, B.Z.; Chen, S.B.; Wang, S.B. Investigation on the resources of Puerariae. J. Chin. Med. Mater. 1993, 08, 13–14. [Google Scholar] [CrossRef]
- Lan, L. Research progress of puerariae. Strait Pharm. J. 2013, 25, 10–14. [Google Scholar]
- Shi, C.X.; Du, J.R.; Wu, W.; Liu, L.X.; Yang, G.L.; Zhang, H.Y. Chemical Constituents and Pharmacological Action of Puerariae Lobatae Radix: A Review. Mod. Chin. Med. 2021, 23, 2177–2195. [Google Scholar] [CrossRef]
- Song, W.; Li, Y.J.; Qiao, X.; Qian, Y.; Ye, M. Chemistry of the Chinese herbal medicine Puerariae Radix (Ge-Gen): A review. J. Chin. Pharm. Sci. 2014, 23, 347–360. [Google Scholar] [CrossRef]
- Ma, X.; Yan, L.; Zhu, Q.; Shao, F. Puerarin attenuates cisplatin-induced rat nephrotoxicity: The involvement of TLR4/NF-kappa B signaling pathway. PLoS ONE 2017, 12, e0171612. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Zhang, X.; Xu, X.; Yu, Y. Effects of puerarin on the retina and STAT3 expression in diabetic rats. Exp. Ther. Med. 2017, 14, 5480–5484. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Zhu, X.; Zhang, K.; Yao, Y.; Zhuang, M.; Tan, C.; Zhou, F.; Zhu, L. Puerarin inhibits amyloid beta-induced NLRP3 inflammasome activation in retinal pigment epithelial cells via suppressing ROS-dependent oxidative and endoplasmic reticulum stresses. Exp. Cell Res. 2017, 357, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Leng, W.; Zhang, J. Protective Effect of Puerarin Against Oxidative Stress Injury of Neural Cells and Related Mechanisms. Med. Sci. Monit. 2016, 22, 1244–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.-Q.; Ding, J.; Xiao, Z.-H.; Liu, C.-M. Puerarin ameliorates carbon tetrachloride-induced oxidative DNA damage and inflammation in mouse kidney through ERK/Nrf2/ARE pathway. Food Chem. Toxicol. 2014, 71, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.S.; Chen, B.D.; Lin, B.Y.; Song, M.H.; Li, Z.X. Puerarin protects traumatic brain injury through Nrf2-ARE pathway to resist oxidative stress. Stroke Nerv. Dis. 2017, 24, 91–94. [Google Scholar]
- Luan, B.; Luan, M.; Qiu, Y.H. Effect of puerarin on activation of Nrf2 pathway in relieving oxidative stress injury in rats with heart failure. Mod. Prev. Med. 2019, 46, 1852–1856. [Google Scholar]
- Zhang, Y.; Wang, D.; Fu, H.; Zhao, D.; Zhang, J.; Li, M.; Wang, C. Protective effects of extracellular proteins of Saccharomycopsis fibuligera on UVA-damaged human skin fibroblasts. J. Funct. Foods 2022, 88, 104897. [Google Scholar] [CrossRef]
- Li, Z.L.; Ouyang, W.Q.; Zhang, L.; Liu, Y.M.; Yang, G.M. Protective effects of EMP on oxidative damage of mice skin irradiated by ultraviolet A. J. Northwest AF Univ. (Nat. Sci. Ed.) 2007, 35, 19–22. [Google Scholar] [CrossRef]
- Mo, Q.; Fu, H.; Zhao, D.; Zhang, J.; Wang, C.; Wang, D.; Li, M. Protective Effects of Mogroside V on Oxidative Stress Induced by H2O2 in Skin Fibroblasts. Drug Des. Dev. Ther. 2021, 15, 4901–4909. [Google Scholar] [CrossRef]
- Zhang, W.Y.; Zhang, F.Q. Enzymatic hydrolysis and antioxidant activity of polypeptide from eggshell membrane. Food Res. Dev. 2021, 42, 47–55. [Google Scholar]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuryana, C.T.; Haryana, S.M.; Wirohadidjojo, Y.W.; Arfian, N. Achatina fulica mucous improves cell viability and increases collagen deposition in UVB-irradiated human fibroblast culture. J. Stem Cells Regen. Med. 2020, 16, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.X.; Zhao, H. Efficacy evaluation of cosmetics (Ⅱ) Scientific support for moisturizing cosmetic efficacy claims. China Surfactant Deterg. Cosmet. 2018, 48, 67–72. [Google Scholar] [CrossRef]
- Tian, Y.; Liu, W.; Niu, T.; Dai, C.; Li, X.; Cui, C.; Zhao, X.; E, Y.; Lu, H. The Injury and Cumulative Effects on Human Skin by UV Exposure from Artificial Fluorescence Emission. Photochem. Photobiol. 2014, 90, 1433–1438. [Google Scholar] [CrossRef] [PubMed]
- Rabe, J.H.; Mamelak, A.J.; McElgunn, P.J.S.; Morison, W.L.; Sauder, D.N. Photoaging: Mechanisms and repair. J. Am. Acad. Dermatol. 2006, 55, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Kong, A.-N. Molecular mechanisms of Nrf2-mediated antioxidant response. Mol. Carcinog. 2009, 48, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Gillette, D.D.; Li, X.; Zhang, Z.; Wen, H. Nuclear Factor E2-related Factor-2 (Nrf2) Is Required for NLRP3 and AIM2 Inflammasome Activation. J. Biol. Chem. 2014, 289, 17020–17029. [Google Scholar] [CrossRef] [Green Version]
- Jaeeun, L.; Weon, L.J.; Hyeyoung, K. Lycopene Inhibits IL-6 Expression by Upregulating NQO1 and HO-1 via Activation of Nrf2 in Ethanol/Lipopolysaccharide-Stimulated Pancreatic Acinar Cells. Antioxidants 2022, 11, 519. [Google Scholar] [CrossRef]
- Tsai, B.C.-K.; Hsieh, D.J.-Y.; Lin, W.-T.; Tamilselvi, S.; Day, C.H.; Ho, T.-J.; Chang, R.-L.; Viswanadha, V.P.; Kuo, C.-H.; Huang, C.-Y. Functional potato bioactive peptide intensifies Nrf2-dependent antioxidant defense against renal damage in hypertensive rats. Food Res. Int. 2020, 129, 108862. [Google Scholar] [CrossRef]
- Hseu, Y.-C.; Chang, C.-T.; Gowrisankar, Y.V.; Chen, X.-Z.; Lin, H.-C.; Yen, H.-R.; Yang, H.-L. Zerumbone Exhibits Antiphotoaging and Dermatoprotective Properties in Ultraviolet A-Irradiated Human Skin Fibroblast Cells via the Activation of Nrf2/ARE Defensive Pathway. Oxidative Med. Cell. Longev. 2019, 2019, 4098674. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, T.; Ishitsuka, Y.; Nakamura, Y.; Okiyama, N.; Watanabe, R.; Fujisawa, Y.; Fujimoto, M. Honey and Chamomile Activate Keratinocyte Antioxidative Responses via the KEAP1/NRF2 System. Clin. Cosmet. Investig. Dermatol. 2020, 13, 657–660. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, J.; Zhang, X.; Dong, M. Puerarin suppresses MPP+/MPTP-induced oxidative stress through an Nrf2-dependent mechanism. Food Chem. Toxicol. 2020, 144, 111644. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.-D.; Lee, J.-H.; Lee, Y.-M.; Kim, D.-K. Puerarin inhibits inflammation and oxidative stress in dextran sulfate sodium-induced colitis mice model. Biomed. Pharmacother. 2020, 124, 109847. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.-M.; Chen, J.; Wang, M.; Zheng, C.; Zhou, X.-L. Puerarin attenuates cadmium-induced hepatic lipid metabolism disorder by inhibiting oxidative stress and inflammation in mice. J. Inorg. Biochem. 2021, 222, 111521. [Google Scholar] [CrossRef] [PubMed]
- Xia, D.-Z.; Zhang, P.-H.; Fu, Y.; Yu, W.-F.; Ju, M.-T. Hepatoprotective activity of puerarin against carbon tetrachloride-induced injuries in rats: A randomized controlled trial. Food Chem. Toxicol. 2013, 59, 90–95. [Google Scholar] [CrossRef]
- Ali, S.S.; Ahsan, H.; Zia, M.K.; Siddiqui, T.; Khan, F.H. Understanding oxidants and antioxidants: Classical team with new players. J. Food Biochem. 2020, 44, e13145. [Google Scholar] [CrossRef]
- Ayer, A.; Zarjou, A.; Agarwal, A.; Stocker, R. Heme Oxygenases in Cardiovascular Health and Disease. Physiol. Rev. 2016, 96, 1449–1508. [Google Scholar] [CrossRef] [Green Version]
- Parzonko, A.; Kiss, A.K. Caffeic acid derivatives isolated from Galinsoga parviflora herb protected human dermal fibroblasts from UVA-radiation. Phytomedicine 2019, 57, 215–222. [Google Scholar] [CrossRef]
- Jung, K.-A.; Kwak, M.-K. The Nrf2 System as a Potential Target for the Development of Indirect Antioxidants. Molecules 2010, 15, 7266–7291. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.C. Glutathione synthesis. Biochim. Biophys. Acta-Gen. Subj. 2013, 1830, 3143–3153. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yang, L. Molecular regulatory mechanism of Nrf2 antioxidant. China J. Bioinform. 2018, 16, 1–6. [Google Scholar]
- Son, Y.; Kim, S.; Chung, H.-T.; Pae, H.-O. Chapter Two—Reactive Oxygen Species in the Activation of MAP Kinases. In Methods in Enzymology; Cadenas, E., Packer, L., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 528, pp. 27–48. [Google Scholar]
- Zhao, M.; Zhu, P.; Fujino, M.; Nishio, Y.; Chen, J.; Ito, H.; Takahashi, K.; Nakajima, M.; Tanaka, T.; Zhao, L.; et al. 5-Aminolevulinic acid with sodium ferrous citrate induces autophagy and protects cardiomyocytes from hypoxia-induced cellular injury through MAPK-Nrf-2-HO-1 signaling cascade. Biochem. Biophys. Res. Commun. 2016, 479, 663–669. [Google Scholar] [CrossRef]
- Zhang, H.; Yuan, B.; Huang, H.; Qu, S.; Yang, S.; Zeng, Z. Gastrodin induced HO-1 and Nrf2 up-regulation to alleviate H2O2-induced oxidative stress in mouse liver sinusoidal endothelial cells through p38 MAPK phosphorylation. Braz. J. Med. Biol. Res. 2018, 51, e7439. [Google Scholar] [CrossRef]
- Wang, Z.; Ka, S.-O.; Lee, Y.; Park, B.-H.; Bae, E.J. Butein induction of HO-1 by p38 MAPK/Nrf2 pathway in adipocytes attenuates high-fat diet induced adipose hypertrophy in mice. Eur. J. Pharmacol. 2017, 799, 201–210. [Google Scholar] [CrossRef]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-Degrading Metalloproteinases in Photoaging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Kammeyer, A.; Luiten, R.M. Oxidation events and skin aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef]
- Ruggiero, F.; Roulet, M.; Bonod-Bidaud, C. [Dermis collagens: Beyond their structural properties]. J. Soc. Biol. 2005, 199, 301–311. [Google Scholar] [CrossRef]
- Duan, X.; Wu, T.; Liu, T.; Yang, H.; Ding, X.; Chen, Y.; Mu, Y. Vicenin-2 ameliorates oxidative damage and photoaging via modulation of MAPKs and MMPs signaling in UVB radiation exposed human skin cells. J. Photochem. Photobiol. B Biol. 2019, 190, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hou, H.; Fan, Y.; Wang, S.; Chen, Q.; Si, L.; Li, B. Protective effect of gelatin peptides from pacific cod skin against photoaging by inhibiting the expression of MMPs via MAPK signaling pathway. J. Photochem. Photobiol. B Biol. 2016, 165, 34–41. [Google Scholar] [CrossRef]
- Ma, J.-Q.; Ding, J.; Zhao, H.; Liu, C.-M. Puerarin Attenuates Carbon Tetrachloride-Induced Liver Oxidative Stress and Hyperlipidaemia in Mouse by JNK/c-Jun/CYP7A1 Pathway. Basic Clin. Pharmacol. Toxicol. 2014, 115, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Pan, S.Q.; Shen, C.; Pan, S.F.; Zhang, X.M.; He, Q.Y. Puerarin inhibits angiotensin II-induced cardiac hypertrophy via the redox-sensitive ERK1/2, p38 and NF-kappa B pathways. Acta Pharmacol. Sin. 2014, 35, 463–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.; Zhao, Y.; Li, Q.; Han, X.; Duan, J. Puerarin protects against iron overload-induced retinal injury through regulation of iron-handling proteins. Biomed. Pharmacother. 2020, 122, 109690. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tang, Y.; Sun, Y.; Veeraraghavan, V.P.; Mohan, S.K.; Cui, C. 6-shogaol, a active constiuents of ginger prevents UVB radiation mediated inflammation and oxidative stress through modulating NrF2 signaling in human epidermal keratinocytes (HaCaT cells). J. Photochem. Photobiol. B Biol. 2019, 197, 111518. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Pan, Z.; Xu, T.; Zhang, C.; Wu, Q.; Niu, Y. Puerarin induces the upregulation of glutathione levels and nuclear translocation of Nrf2 through PI3K/Akt/GSK-3 beta signaling events in PC12 cells exposed to lead. Neurotoxicology Teratol. 2014, 46, 1–9. [Google Scholar] [CrossRef]








| Gene | Direction | Primer Pair Sequence (5′→3′) |
|---|---|---|
| β-actin | F | TGGCACCCAGCACAATGAA |
| R | CTAAGTCATAGTCCGCCTAGAAGCA | |
| Nrf2 | F | CAACTCAGCACCTTGTATC |
| R | TTCTTAGTATCTGGCTTCTT | |
| KEAP1 | F | GGAGGCGGAGCCCGA |
| R | GATGCCCTCAATGGACACCA | |
| HO-1 | F | CAAGCGCTATGTTCAGCGAC |
| R | GCTTGAACTTGGTGGCACTG | |
| NQO1 | F | CAGCCAATCAGCGTTCGGTA |
| R | CTTCATGGCGTAGTTGAATGATGTC | |
| GCLC | F | CAGTCAAGGACCGGCACAAG |
| R | CAAGAACATCGCCTCCATTCAG | |
| ERK | F | TGTTCCCAAATGCTGACTCCAA |
| R | TCGGGTCGTAATACTGCTCCAGATA | |
| JNK | F | CTGTGTGGAATCAAGCACCTTCA |
| R | CTGGCCAGACCGAAGTCAAGA | |
| P38 | F | TTAACAGGATGCCAAGCCATGA |
| R | GGCACCAATAAATACATTCGCAAAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, Q.; Li, S.; You, S.; Wang, D.; Zhang, J.; Li, M.; Wang, C. Puerarin Reduces Oxidative Damage and Photoaging Caused by UVA Radiation in Human Fibroblasts by Regulating Nrf2 and MAPK Signaling Pathways. Nutrients 2022, 14, 4724. https://doi.org/10.3390/nu14224724
Mo Q, Li S, You S, Wang D, Zhang J, Li M, Wang C. Puerarin Reduces Oxidative Damage and Photoaging Caused by UVA Radiation in Human Fibroblasts by Regulating Nrf2 and MAPK Signaling Pathways. Nutrients. 2022; 14(22):4724. https://doi.org/10.3390/nu14224724
Chicago/Turabian StyleMo, Qiuting, Shuping Li, Shiquan You, Dongdong Wang, Jiachan Zhang, Meng Li, and Changtao Wang. 2022. "Puerarin Reduces Oxidative Damage and Photoaging Caused by UVA Radiation in Human Fibroblasts by Regulating Nrf2 and MAPK Signaling Pathways" Nutrients 14, no. 22: 4724. https://doi.org/10.3390/nu14224724
APA StyleMo, Q., Li, S., You, S., Wang, D., Zhang, J., Li, M., & Wang, C. (2022). Puerarin Reduces Oxidative Damage and Photoaging Caused by UVA Radiation in Human Fibroblasts by Regulating Nrf2 and MAPK Signaling Pathways. Nutrients, 14(22), 4724. https://doi.org/10.3390/nu14224724