
Immune Impairment Associated with Vitamin A Deficiency: Insights from Clinical Studies and Animal Model Research

 ,
,  , and
, and
Abstract
:1. Introduction
2. Vitamin A Deficiency and the Associated Pathology
2.1. How Vitamin A Deficiency Occurs
2.2. Pathologies Associated with VAD
3. Vitamin A Deficiency Dysregulates Immune Function
3.1. Vitamin A Deficiency Increases Disease Severity
3.1.1. VAD Influences the Immune Response to Parasitic Infections While the Latter Modulate VA Metabolism and Bioavailability
3.1.2. VAD Impacts Immune Response to Viral Infections
3.1.3. VAD Impairs Immune Response to Bacterial Infections
3.2. Vitamin A Deficiency Influences Immune Responses to Vaccines
4. Gut Microbiota Adaptation to Vitamin A Deficiency Impacts Host Immunity

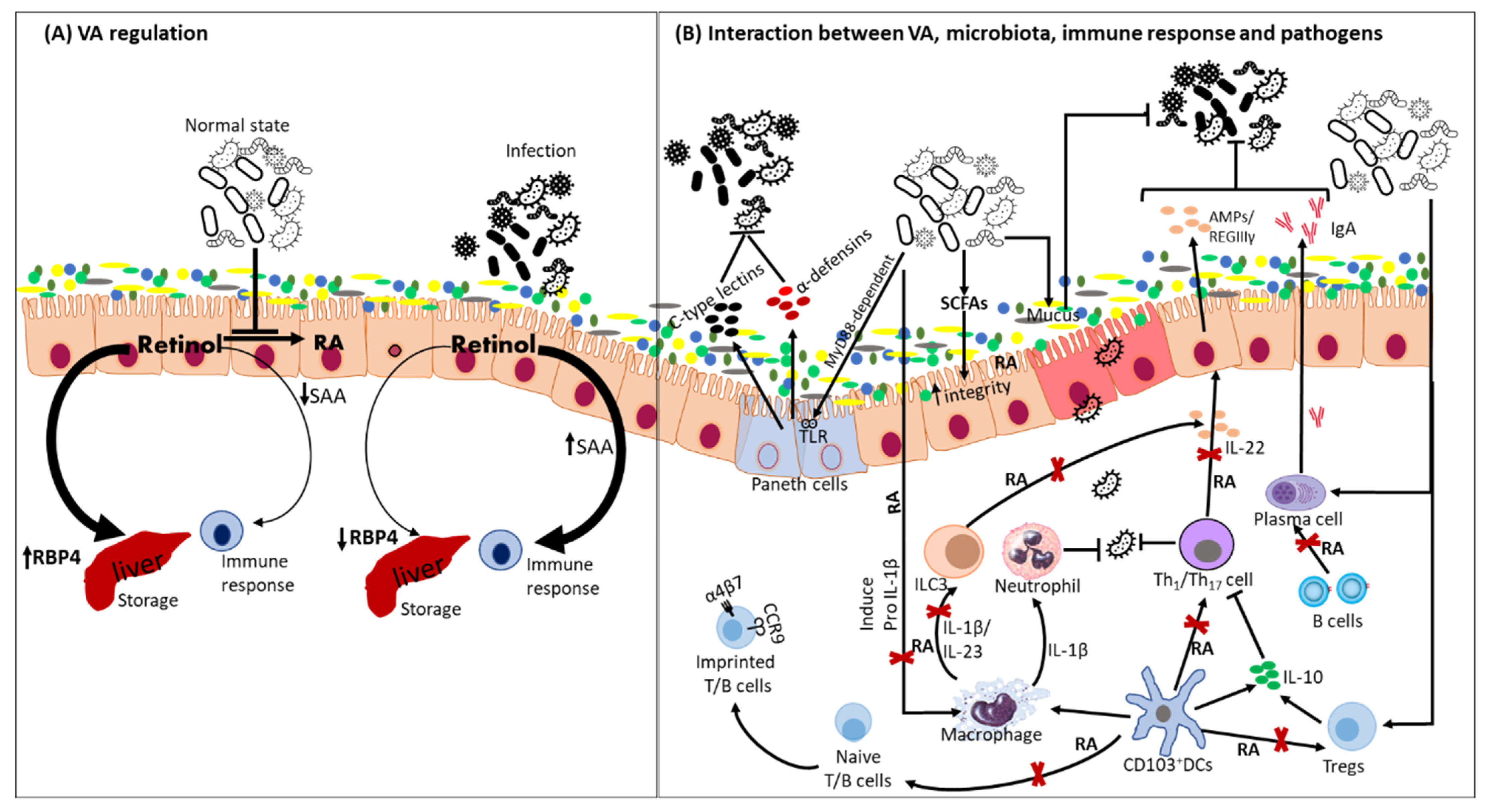{kind=link}
{kind=link}
| Host Model | Pathogen/Study | Key Findings | Reference |
|---|---|---|---|
| Rat | Gut microbiome | A decrease in proportion of Lactobacillus spp. in VAD rats led to decreased MUC2 mRNA expression and increased MUC3 mRNA expression in intestine. | [43] |
| Rat | Gut microbiome | Faecalibacterium abundance was positively correlated with, while Staphylococcus abundance was negatively correlated with, serum retinol levels. | [207] |
| Mouse | Gut microbiome | VAD causes a significant change in the microbiome structure and its meta-transcriptome (increase in Bacteroides vulgatus) in human gut microbiota–associated mice | [200] |
| Mouse | Rotavirus/Influenza virus | Lactobacillus spp. have been shown to exhibit antiviral effects against RVand influenza infections. | [205] |
| Mouse | Gut microbiome | VAD reduced gut microbiome diversity by increasing expression of MUC2 gene by goblet cell hyperplasia, resulting in low IL-17, IRF4, IL-21, IL-22 and IL-23. | [148] |
| Mouse | Alzheimer’s disease Gut microbiome | VAD aggravates gut microbiota dysbiosis and cognitive deficits, reduces the expression of GABA receptors and downregulates BDNF in the brain through intestinal microbiota disruption. | [8,209] |
| Mouse | Norovirus Gut microbiome | The abundance of Lactobacillus sp inhibited murine NoV proliferation via the upregulation of IFN-β; RA administration increased their abundance levels. | [150] |
| Human | Gut microbiome polio virus, tetanus toxoid, hepatitis B virus vaccines | Bacterial diversity and abundance of Enterobacteriales, Pseudomonadales and Clostridiales were associated with neutrophilia and lower vaccine responses. | [9,215] |
| Human | Cholera vaccine | High abundance of Clostridiales in responders, while poor vaccine response was associated with Enterobacterales | [216] |
| Human | Rotavirus vaccine | Negative association between vaccine immunogenicity and gut microbiota diversity. | [217,218] |
| Pig | Influenza A virus | Influenza vaccine is more effective in pigs with Prevotella colonization than Helicobacter and Bacteroides colonization. | [219] |
| Chicken | Campylobacter jejuni/Vaccine | Abundance of Clostridium spp., Ruminococcaceae and Lachnospiraceae was positively associated with vaccine-induced antigen-specific IgY responses. | [220] |
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Villamor, E.; Fawzi, W.W. Effects of vitamin A supplementation on immune responses and correlation with clinical outcomes. Clin. Microbiol. Rev. 2005, 18, 446–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, S.A.; McCaffery, P.J.; Drager, U.C.; De Luca, L.M. Retinoids in embryonal development. Physiol. Rev. 2000, 80, 1021–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, S.R.; Moise, A.R.; Trainor, P.A. New insights and changing paradigms in the regulation of vitamin a metabolism in development. Wiley Interdiscip. Rev. Dev. Biol. 2017, 6, e264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riabroy, N.; Tanumihardjo, S.A. Oral doses of alpha-retinyl ester track chylomicron uptake and distribution of vitamin a in a male piglet model for newborn infants. J. Nutr. 2014, 144, 1188–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora, J.R.; Iwata, M.; von Andrian, U.H. Vitamin effects on the immune system: Vitamins a and d take centre stage. Nat. Rev. Immunol. 2008, 8, 685–698. [Google Scholar] [CrossRef] [Green Version]
- Gombart, A.F.; Pierre, A.; Maggini, S. A review of micronutrients and the immune system-working in harmony to reduce the risk of infection. Nutrients 2020, 12, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maggini, S.; Beveridge, S.; Sorbara, J.; Senatore, G. Feeding the immune system: The role of micronutrients in restoring resistance to infections. CAB Rev. 2008, 3, 1–21. [Google Scholar] [CrossRef]
- Chen, B.W.; Zhang, K.W.; Chen, S.J.; Yang, C.; Li, P.G. Vitamin A deficiency exacerbates gut microbiota dysbiosis and cognitive deficits in amyloid precursor protein/presenilin 1 transgenic mice. Front. Aging Neurosci. 2021, 13, 753351. [Google Scholar] [CrossRef]
- Huda, M.N.; Ahmad, S.M.; Kalanetra, K.M.; Taft, D.H.; Alam, M.J.; Khanam, A.; Raqib, R.; Underwood, M.A.; Mills, D.A.; Stephensen, C.B. Neonatal vitamin A supplementation and vitamin A status are associated with gut microbiome composition in bangladeshi infants in early infancy and at 2 years of age. J. Nutr. 2019, 149, 1075–1088. [Google Scholar] [CrossRef] [Green Version]
- Timoneda, J.; Rodriguez-Fernandez, L.; Zaragoza, R.; Marin, M.P.; Cabezuelo, M.T.; Torres, L.; Vina, J.R.; Barber, T. Vitamin A deficiency and the lung. Nutrients 2018, 10, 1132. [Google Scholar] [CrossRef]
- Nauss, K.M.; Anderson, C.A.; Conner, M.W.; Newberne, P.M. Ocular infection with herpes simplex virus (HSV-1) in vitamin A-deficient and control rats. J. Nutr. 1985, 115, 1300–1315. [Google Scholar] [CrossRef] [PubMed]
- Wiedermann, U.; Tarkowski, A.; Bremell, T.; Hanson, L.A.; Kahu, H.; Dahlgren, U.I. Vitamin A deficiency predisposes to Staphylococcus aureus infection. Infect. Immun. 1996, 64, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, D.; Moldoveanu, Z.; Stephensen, C.B. High-level dietary vitamin A enhances T-helper type 2 cytokine production and secretory immunoglobulin A response to influenza A virus infection in balb/c mice. J. Nutr. 2000, 130, 1132–1139. [Google Scholar] [CrossRef] [Green Version]
- Nikawa, T.; Odahara, K.; Koizumi, H.; Kido, Y.; Teshima, S.; Rokutan, K.; Kishi, K. Vitamin a prevents the decline in immunoglobulin A and Th2 cytokine levels in small intestinal mucosa of protein-malnourished mice. J. Nutr. 1999, 129, 934–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirisinha, S.; Darip, M.D.; Moongkarndi, P.; Ongsakul, M.; Lamb, A.J. Impaired local immune response in vitamin A-deficient rats. Clin. Exp. Immunol. 1980, 40, 127–135. [Google Scholar] [PubMed]
- Chattha, K.S.; Kandasamy, S.; Vlasova, A.N.; Saif, L.J. Vitamin A deficiency impairs adaptive B and T cell responses to a prototype monovalent attenuated human rotavirus vaccine and virulent human rotavirus challenge in a gnotobiotic piglet model. PLoS ONE 2013, 8, e82966. [Google Scholar] [CrossRef] [Green Version]
- Stevens, G.A.; Bennett, J.E.; Hennocq, Q.; Lu, Y.; De-Regil, L.M.; Rogers, L.; Danaei, G.; Li, G.; White, R.A.; Flaxman, S.R.; et al. Trends and mortality effects of vitamin A deficiency in children in 138 low-income and middle-income countries between 1991 and 2013: A pooled analysis of population-based surveys. Lancet Glob. Health 2015, 3, e528–e536. [Google Scholar] [CrossRef] [Green Version]
- Stephensen, C.B.; Alvarez, J.O.; Kohatsu, J.; Hardmeier, R.; Kennedy, J.I., Jr.; Gammon, R.B., Jr. Vitamin A is excreted in the urine during acute infection. Am. J. Clin. Nutr. 1994, 60, 388–392. [Google Scholar] [CrossRef]
- Quadro, L.; Blaner, W.S.; Salchow, D.J.; Vogel, S.; Piantedosi, R.; Gouras, P.; Freeman, S.; Cosma, M.P.; Colantuoni, V.; Gottesman, M.E. Impaired retinal function and vitamin A availability in mice lacking retinol-binding protein. EMBO J. 1999, 18, 4633–4644. [Google Scholar] [CrossRef]
- Quadro, L.; Hamberger, L.; Colantuoni, V.; Gottesman, M.E.; Blaner, W.S. Understanding the physiological role of retinol-binding protein in vitamin A metabolism using transgenic and knockout mouse models. Mol. Asp. Med. 2003, 24, 421–430. [Google Scholar] [CrossRef]
- Raghu, P.; Sivakumar, B. Interactions amongst plasma retinol-binding protein, transthyretin and their ligands: Implications in vitamin A homeostasis and transthyretin amyloidosis. Biochim. Biophys. Acta (BBA)—Proteins Proteom. 2004, 1703, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chow, E.K.; Castrillo, A.; Shahangian, A.; Pei, L.; O’Connell, R.M.; Modlin, R.L.; Tontonoz, P.; Cheng, G. A role for irf3-dependent rxralpha repression in hepatotoxicity associated with viral infections. J. Exp. Med. 2006, 203, 2589–2602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, M.H.; Green, J.B.; Berg, T.; Norum, K.R.; Blomhoff, R. Changes in hepatic parenchymal and nonparenchymal cell vitamin A content during vitamin A depletion in the rat. J. Nutr. 1988, 118, 1331–1335. [Google Scholar] [CrossRef]
- Matsuura, T.; Ross, A.C. Regulation of hepatic lecithin: Retinol acyltransferase activity by retinoic acid. Arch. Biochem. Biophys. 1993, 301, 221–227. [Google Scholar] [CrossRef]
- Grumet, L.; Taschler, U.; Lass, A. Hepatic retinyl ester hydrolases and the mobilization of retinyl ester stores. Nutrients 2016, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Saeed, A.; Bartuzi, P.; Heegsma, J.; Dekker, D.; Kloosterhuis, N.; de Bruin, A.; Jonker, J.W.; van de Sluis, B.; Faber, K.N. Impaired hepatic vitamin A metabolism in nafld mice leading to vitamin A accumulation in hepatocytes. Cell. Mol. Gastroenterol. Hepatol. 2021, 11, 309–325. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.; Dullaart, R.P.F.; Schreuder, T.; Blokzijl, H.; Faber, K.N. Disturbed vitamin A metabolism in non-alcoholic fatty liver disease (nafld). Nutrients 2017, 10, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Binali, H.A. Night blindness and ancient remedy. Heart Views Off. J. Gulf Heart Assoc. 2014, 15, 136–139. [Google Scholar] [CrossRef] [PubMed]
- Velasco Cruz, A.A.; Attie-Castro, F.A.; Fernandes, S.L.; Cortes, J.F.; de Tarso, P.P.-F.P.; Rocha, E.M.; Marchini, J.S. Adult blindness secondary to vitamin A deficiency associated with an eating disorder. Nutrition 2005, 21, 630–633. [Google Scholar] [CrossRef] [PubMed]
- Lateef, H.; Abatan, O.I.; Aslam, M.N.; Stevens, M.J.; Varani, J. Topical pretreatment of diabetic rats with all-trans retinoic acid improves healing of subsequently induced abrasion wounds. Diabetes 2005, 54, 855–861. [Google Scholar] [CrossRef]
- Wicke, C.; Halliday, B.; Allen, D.; Roche, N.S.; Scheuenstuhl, H.; Spencer, M.M.; Roberts, A.B.; Hunt, T.K. Effects of steroids and retinoids on wound healing. Arch. Surg. 2000, 135, 1265–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.Y.; Chung, J.; Kim, M.K.; Kwon, S.O.; Cho, B.H. Antioxidant nutrient intakes and corresponding biomarkers associated with the risk of atopic dermatitis in young children. Eur. J. Clin. Nutr. 2010, 64, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Schindler, M.; Drozdenko, G.; Kuhl, A.A.; Worm, M. Immunomodulation in patients with chronic hand eczema treated with oral alitretinoin. Int. Arch. Allergy Immunol. 2014, 165, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Sommer, A. Vitamin A deficiency and clinical disease: An historical overview. J. Nutr. 2008, 138, 1835–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolbach, S.B.; Howe, P.R. Epithelial repair in recovery from vitamin a deficiency: An experimental study. J. Exp. Med. 1933, 57, 511–526. [Google Scholar] [CrossRef] [PubMed]
- de Medeiros, P.; Pinto, D.V.; de Almeida, J.Z.; Rego, J.M.C.; Rodrigues, F.A.P.; Lima, A.A.M.; Bolick, D.T.; Guerrant, R.L.; Oria, R.B. Modulation of intestinal immune and barrier functions by vitamin A: Implications for current understanding of malnutrition and enteric infections in children. Nutrients 2018, 10, 1128. [Google Scholar] [CrossRef] [Green Version]
- Ghishan, F.K.; Kiela, P.R. Vitamins and minerals in inflammatory bowel disease. Gastroenterol. Clin. North Am. 2017, 46, 797–808. [Google Scholar] [CrossRef]
- Surman, S.L.; Penkert, R.R.; Sealy, R.E.; Jones, B.G.; Marion, T.N.; Vogel, P.; Hurwitz, J.L. Consequences of vitamin A deficiency: Immunoglobulin dysregulation, squamous cell metaplasia, infectious disease, and death. Int. J. Mol. Sci. 2020, 21, 5570. [Google Scholar] [CrossRef]
- Ugalde-Silva, P.; Gonzalez-Lugo, O.; Navarro-Garcia, F. Tight junction disruption induced by type 3 secretion system effectors injected by enteropathogenic and enterohemorrhagic Escherichia coli. Front. Cell. Infect. Microbiol. 2016, 6, 87. [Google Scholar] [CrossRef] [Green Version]
- Stephensen, C.B.; Blount, S.R.; Schoeb, T.R.; Park, J.Y. Vitamin A deficiency impairs some aspects of the host response to influenza a virus infection in balb/c mice. J. Nutr. 1993, 123, 823–833. [Google Scholar] [CrossRef]
- Penkert, R.R.; Surman, S.L.; Jones, B.G.; Sealy, R.E.; Vogel, P.; Neale, G.; Hurwitz, J.L. Vitamin A deficient mice exhibit increased viral antigens and enhanced cytokine/chemokine production in nasal tissues following respiratory virus infection despite the presence of FOXP3+ T cells. Int. Immunol. 2016, 28, 139–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Liu, S.; Liu, G.; Zhao, J.; Jiao, H.; Wang, X.; Song, Z.; Lin, H. Vitamin A deficiency impairs mucin expression and suppresses the mucosal immune function of the respiratory tract in chicks. PLoS ONE 2015, 10, e0139131. [Google Scholar] [CrossRef] [Green Version]
- Amit-Romach, E.; Uni, Z.; Cheled, S.; Berkovich, Z.; Reifen, R. Bacterial population and innate immunity-related genes in rat gastrointestinal tract are altered by vitamin A-deficient diet. J. Nutr. Biochem. 2009, 20, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Jones, D.B.; Jackson, A.A. The interaction of vitamin A deficiency and rotavirus infection in the mouse. Br. J. Nutr. 1990, 63, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Yi, P.; Wang, X.; Zhang, B.; Jie, Z.; Soong, L.; Sun, J. Retinoic acid modulates hyperactive T cell responses and protects vitamin a-deficient mice against persistent lymphocytic choriomeningitis virus infection. J. Immunol. 2020, 204, 2984–2994. [Google Scholar] [CrossRef] [PubMed]
- Kuwata, T.; Wang, I.M.; Tamura, T.; Ponnamperuma, R.M.; Levine, R.; Holmes, K.L.; Morse, H.C.; De Luca, L.M.; Ozato, K. Vitamin A deficiency in mice causes a systemic expansion of myeloid cells. Blood 2000, 95, 3349–3356. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kitano, T.; Oma, M.; Hiyama, Y.; Yumoto, S.; Matsui, M. Supplemental effect of retinoids on immume function in vitamin A deficient rats. Nutr. Res. 1988, 8, 529–538. [Google Scholar] [CrossRef]
- Nauss, K.M.; Newberne, P.M. Local and regional immune function of vitamin A-deficient rats with ocular herpes simplex virus (HSV) infections. J. Nutr. 1985, 115, 1316–1324. [Google Scholar] [CrossRef]
- Wolbach, S.B.; Howe, P.R. Tissue changes following deprivation of fat-soluble a vitamin. J. Exp. Med. 1925, 42, 753–777. [Google Scholar] [CrossRef] [Green Version]
- Clagett-Dame, M.; Knutson, D. Vitamin A in reproduction and development. Nutrients 2011, 3, 385–428. [Google Scholar] [CrossRef]
- Zile, M.H. Function of vitamin A in vertebrate embryonic development. J. Nutr. 2001, 131, 705–708. [Google Scholar] [CrossRef] [Green Version]
- El-Khashab, E.K.; Hamdy, A.M.; Maher, K.M.; Fouad, M.A.; Abbas, G.Z. Effect of maternal vitamin A deficiency during pregnancy on neonatal kidney size. J. Perinat. Med. 2013, 41, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.G.; Roth, C.B.; Warkany, J. An analysis of the syndrome of malformations induced by maternal vitamin A deficiency. Effects of restoration of vitamin A at various times during gestation. Am. J. Anat. 1953, 92, 189–217. [Google Scholar] [CrossRef]
- Chien, C.Y.; Lee, H.S.; Cho, C.H.; Lin, K.I.; Tosh, D.; Wu, R.R.; Mao, W.Y.; Shen, C.N. Maternal vitamin A deficiency during pregnancy affects vascularized islet development. J. Nutr. Biochem. 2016, 36, 51–59. [Google Scholar] [CrossRef]
- Matthews, K.A.; Rhoten, W.B.; Driscoll, H.K.; Chertow, B.S. Vitamin A deficiency impairs fetal islet development and causes subsequent glucose intolerance in adult rats. J. Nutr. 2004, 134, 1958–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlasova, A.N.; Chattha, K.S.; Kandasamy, S.; Siegismund, C.S.; Saif, L.J. Prenatally acquired vitamin A deficiency alters innate immune responses to human rotavirus in a gnotobiotic pig model. J. Immunol. 2013, 190, 4742–4753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandasamy, S.; Chattha, K.S.; Vlasova, A.N.; Saif, L.J. Prenatal vitamin A deficiency impairs adaptive immune responses to pentavalent rotavirus vaccine (Rotateq®) in a neonatal gnotobiotic pig model. Vaccine 2014, 32, 816–824. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Y.; Qi, G.; Brand, D.; Zheng, S.G. Role of vitamin A in the immune system. J. Clin. Med. 2018, 7, 258. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.M.; Haskell, M.J.; Raqib, R.; Stephensen, C.B. Markers of innate immune function are associated with vitamin A stores in men. J. Nutr. 2009, 139, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Maggini, S.; Pierre, A.; Calder, P.C. Immune function and micronutrient requirements change over the life course. Nutrients 2018, 10, 1531. [Google Scholar] [CrossRef]
- Twining, S.S.; Schulte, D.P.; Wilson, P.M.; Fish, B.L.; Moulder, J.E. Vitamin A deficiency alters rat neutrophil function. J. Nutr. 1997, 127, 558–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or adaptive immunity? The example of natural killer cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, L.M.; Teixeira, F.M.E.; Sato, M.N. Impact of retinoic acid on immune cells and inflammatory diseases. Mediat. Inflamm. 2018, 2018, 3067126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, S.P.; Wilhelm, C.; Yang, Q.; Hall, J.A.; Bouladoux, N.; Boyd, A.; Nutman, T.B.; Urban, J.F., Jr.; Wang, J.; Ramalingam, T.R.; et al. Adaptation of innate lymphoid cells to a micronutrient deficiency promotes type 2 barrier immunity. Science 2014, 343, 432–437. [Google Scholar] [CrossRef] [Green Version]
- van de Pavert, S.A.; Ferreira, M.; Domingues, R.G.; Ribeiro, H.; Molenaar, R.; Moreira-Santos, L.; Almeida, F.F.; Ibiza, S.; Barbosa, I.; Goverse, G.; et al. Maternal retinoids control type 3 innate lymphoid cells and set the offspring immunity. Nature 2014, 508, 123–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Mumm, J.B.; Herbst, R.; Kolbeck, R.; Wang, Y. Il-22 increases permeability of intestinal epithelial tight junctions by enhancing claudin-2 expression. J. Immunol. 2017, 199, 3316–3325. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Tian, Q.; Wu, Y.; Peng, X.; Chen, Y.; Li, Q.; Zhang, G.; Tian, X.; Ren, L.; Luo, Z. Vitamin A supplement after neonatal Streptococcus pneumoniae pneumonia inhibits the progression of experimental asthma by altering CD4+ T cell subsets. Sci. Rep. 2020, 10, 4214. [Google Scholar] [CrossRef] [Green Version]
- Mora, J.R.; Iwata, M.; Eksteen, B.; Song, S.Y.; Junt, T.; Senman, B.; Otipoby, K.L.; Yokota, A.; Takeuchi, H.; Ricciardi-Castagnoli, P.; et al. Generation of gut-homing IgA-secreting B cells by intestinal dendritic cells. Science 2006, 314, 1157–1160. [Google Scholar] [CrossRef] [Green Version]
- Tokuyama, H.; Tokuyama, Y. Retinoids enhance IgA production by lipopolysaccharide-stimulated murine spleen cells. Cell. Immunol. 1993, 150, 353–363. [Google Scholar] [CrossRef]
- Bono, M.R.; Tejon, G.; Flores-Santibanez, F.; Fernandez, D.; Rosemblatt, M.; Sauma, D. Retinoic acid as a modulator of T cell immunity. Nutrients 2016, 8, 349. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Liu, Y.; Li, Y.; Gu, J.; Liu, J.; Tang, J.; Wang, J.; Ryffel, B.; Shen, Y.; Brand, D.; et al. Differential role of all-trans retinoic acid in promoting the development of CD4+ and CD8+ regulatory T cells. J. Leukoc. Biol. 2014, 95, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schambach, F.; Schupp, M.; Lazar, M.A.; Reiner, S.L. Activation of retinoic acid receptor-alpha favours regulatory T cell induction at the expense of IL-17-secreting T helper cell differentiation. Eur. J. Immunol. 2007, 37, 2396–2399. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.M.; Farley, T.K.; Richoz, N.; Yao, C.; Shih, H.Y.; Petermann, F.; Zhang, Y.; Sun, H.W.; Hayes, E.; Mikami, Y.; et al. Retinoic acid receptor alpha represses a Th9 transcriptional and epigenomic program to reduce allergic pathology. Immunity 2019, 50, 106–120.e10. [Google Scholar] [CrossRef] [Green Version]
- Iwata, M. Retinoic acid production by intestinal dendritic cells and its role in T-cell trafficking. Semin. Immunol. 2009, 21, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Iwata, M.; Hirakiyama, A.; Eshima, Y.; Kagechika, H.; Kato, C.; Song, S.Y. Retinoic acid imprints gut-homing specificity on T cells. Immunity 2004, 21, 527–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, R.; Oderup, C.; Yuan, R.; Lee, M.; Habtezion, A.; Hadeiba, H.; Butcher, E.C. Retinoic acid regulates the development of a gut-homing precursor for intestinal dendritic cells. Mucosal Immunol. 2013, 6, 847–856. [Google Scholar] [CrossRef] [Green Version]
- Mwanza-Lisulo, M.; Kelly, P. Potential for use of retinoic acid as an oral vaccine adjuvant. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2015, 370, 20140145. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, S.I.; Friedrichsen, M.; Boelter, J.; Lyszkiewicz, M.; Kremmer, E.; Pabst, O.; Forster, R. Retinoic acid induces homing of protective T and B cells to the gut after subcutaneous immunization in mice. J. Clin. Investig. 2011, 121, 3051–3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, J.; Gangopadhyay, N.N.; Moldoveanu, Z.; Mestecky, J.; Stephensen, C.B. Vitamin A is required for regulation of polymeric immunoglobulin receptor (pIgR) expression by interleukin-4 and interferon-gamma in a human intestinal epithelial cell line. J. Nutr. 1998, 128, 1063–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenouchi-Ohkubo, N.; Asano, M.; Chihaya, H.; Chung-Hsuing, W.U.; Ishikasa, K.; Moro, I. Retinoic acid enhances the gene expression of human polymeric immunoglobulin receptor (pIgR) by TNF-alpha. Clin. Exp. Immunol. 2004, 135, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Goetghebuer, T.; Brasseur, D.; Dramaix, M.; DeMol, P.; Donnen, P.; Bahwere, P.; Duchateau, J.; Hennart, P. Significance of very low retinol levels during severe protein-energy malnutrition. J. Trop. Pediatr. 1996, 42, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Miura, T.; Takahashi, K. Vitamin A is involved in maintenance of epithelial cells on the bronchioles and cells in the alveoli of rats. J. Nutr. 1993, 123, 634–641. [Google Scholar] [CrossRef]
- Garcia, O.P. Effect of vitamin A deficiency on the immune response in obesity. Proc. Nutr. Soc. 2012, 71, 290–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glasziou, P.P.; Mackerras, D.E. Vitamin A supplementation in infectious diseases: A meta-analysis. BMJ 1993, 306, 366–370. [Google Scholar] [CrossRef] [Green Version]
- Darlow, B.A.; Graham, P.J. Vitamin A supplementation to prevent mortality and short- and long-term morbidity in very low birthweight infants. Cochrane Database Syst. Rev. 2011, CD000501. [Google Scholar] [CrossRef]
- Villamor, E.; Fawzi, W.W. Vitamin a supplementation: Implications for morbidity and mortality in children. J. Infect. Dis. 2000, 182 (Suppl. S1), S122–S133. [Google Scholar] [CrossRef] [Green Version]
- Wiseman, E.M.; Bar-El Dadon, S.; Reifen, R. The vicious cycle of vitamin A deficiency: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 3703–3714. [Google Scholar] [CrossRef] [PubMed]
- McCullough, F.S.; Northrop-Clewes, C.A.; Thurnham, D.I. The effect of vitamin A on epithelial integrity. Proc. Nutr. Soc. 1999, 58, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, J.H.; Agoestina, T.; Wu, L.; Usman, A.; Nurachim, M.; Subardja, D.; Hidayat, S.; Tielsch, J.; West, K.P., Jr.; Sommer, A. Impact of neonatal vitamin A supplementation on infant morbidity and mortality. J. Pediatr. 1996, 128, 489–496. [Google Scholar] [CrossRef]
- Long, K.Z.; Garcia, C.; Ko, G.; Santos, J.I.; Al Mamun, A.; Rosado, J.L.; DuPont, H.L.; Nathakumar, N. Vitamin A modifies the intestinal chemokine and cytokine responses to norovirus infection in mexican children. J. Nutr. 2011, 141, 957–963. [Google Scholar] [CrossRef]
- Xing, Y.; Sheng, K.; Xiao, X.; Li, J.; Wei, H.; Liu, L.; Zhou, W.; Tong, X. Vitamin A deficiency is associated with severe mycoplasma pneumoniae pneumonia in children. Ann. Transl. Med. 2020, 8, 120. [Google Scholar] [CrossRef] [PubMed]
- Langel, S.N.; Paim, F.C.; Alhamo, M.A.; Lager, K.M.; Vlasova, A.N.; Saif, L.J. Oral vitamin A supplementation of porcine epidemic diarrhea virus infected gilts enhances IgA and lactogenic immune protection of nursing piglets. Vet. Res. 2019, 50, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Yuan, Y.; Tao, Y.; Wang, W. Effects of vitamin A deficiency on mucosal immunity and response to intestinal infection in rats. Nutrition 2011, 27, 227–232. [Google Scholar] [CrossRef]
- McDaniel, K.L.; Restori, K.H.; Dodds, J.W.; Kennett, M.J.; Ross, A.C.; Cantorna, M.T. Vitamin A-deficient hosts become nonsymptomatic reservoirs of Escherichia coli-like enteric infections. Infect. Immun. 2015, 83, 2984–2991. [Google Scholar] [CrossRef] [Green Version]
- Thornton, K.A.; Mora-Plazas, M.; Marin, C.; Villamor, E. Vitamin A deficiency is associated with gastrointestinal and respiratory morbidity in school-age children. J. Nutr. 2014, 144, 496–503. [Google Scholar] [CrossRef] [Green Version]
- Solaymani-Mohammadi, S.; Singer, S.M. Host immunity and pathogen strain contribute to intestinal disaccharidase impairment following gut infection. J. Immunol. 2011, 187, 3769–3775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astiazaran-Garcia, H.; Lopez-Teros, V.; Valencia, M.E.; Vazquez-Ortiz, F.; Sotelo-Cruz, N.; Quihui-Cota, L. Giardia lamblia infection and its implications for vitamin A liver stores in school children. Ann. Nutr. Metab. 2010, 57, 228–233. [Google Scholar] [CrossRef]
- Lima, A.A.; Soares, A.M.; Lima, N.L.; Mota, R.M.; Maciel, B.L.; Kvalsund, M.P.; Barrett, L.J.; Fitzgerald, R.P.; Blaner, W.S.; Guerrant, R.L. Effects of vitamin A supplementation on intestinal barrier function, growth, total parasitic, and specific giardia spp infections in brazilian children: A prospective randomized, double-blind, placebo-controlled trial. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Suchdev, P.S.; Davis, S.M.; Bartoces, M.; Ruth, L.J.; Worrell, C.M.; Kanyi, H.; Odero, K.; Wiegand, R.E.; Njenga, S.M.; Montgomery, J.M.; et al. Soil-transmitted helminth infection and nutritional status among urban slum children in kenya. Am. J. Trop. Med. Hyg. 2014, 90, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Hurst, R.J.; Else, K.J. Retinoic acid signalling in gastrointestinal parasite infections: Lessons from mouse models. Parasite Immunol. 2012, 34, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Maciel, B.L.; Lacerda, H.G.; Queiroz, J.W.; Galvao, J.; Pontes, N.N.; Dimenstein, R.; McGowan, S.E.; Pedrosa, L.F.; Jeronimo, S.M. Association of nutritional status with the response to infection with Leishmania chagasi. Am. J. Trop. Med. Hyg. 2008, 79, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Luz, K.G.; Succi, R.C.; Torres, E. Vitamin A serum level in children with visceral leishmaniasis. Rev. Soc. Bras. Med. Trop. 2001, 34, 381–384. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, M.J.S.D.; Silva, M.N.P.D.; Paiva, A.A.; Marreiro, D.D.N.; Luzia, L.A.; Henriques, G.S.; Rondo, P.H.C.; Sene, I.S.; Almeida, A.T.A.; Costa, C.H.N.; et al. Nutritional status and vitamin a and zinc levels in patients with kala-azar in piaui, brazil. Rev. Soc. Bras. Med. Trop. 2021, 54, e08002020. [Google Scholar] [CrossRef]
- Costa, D.L.; Rocha, R.L.; Carvalho, R.M.; Lima-Neto, A.S.; Harhay, M.O.; Costa, C.H.; Barral-Neto, M.; Barral, A.P. Serum cytokines associated with severity and complications of kala-azar. Pathog. Glob. Health 2013, 107, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Vellozo, N.S.; Pereira-Marques, S.T.; Cabral-Piccin, M.P.; Filardy, A.A.; Ribeiro-Gomes, F.L.; Rigoni, T.S.; DosReis, G.A.; Lopes, M.F. All-trans retinoic acid promotes an M1- to M2-phenotype shift and inhibits macrophage-mediated immunity to leishmania major. Front. Immunol. 2017, 8, 1560. [Google Scholar] [CrossRef] [Green Version]
- Gundra, U.M.; Girgis, N.M.; Gonzalez, M.A.; San Tang, M.; Van Der Zande, H.J.P.; Lin, J.D.; Ouimet, M.; Ma, L.J.; Poles, J.; Vozhilla, N.; et al. Vitamin A mediates conversion of monocyte-derived macrophages into tissue-resident macrophages during alternative activation. Nat. Immunol. 2017, 18, 642–653. [Google Scholar] [CrossRef]
- Hautvast, J.L.; Tolboom, J.J.; West, C.E.; Kafwembe, E.M.; Sauerwein, R.W.; van Staveren, W.A. Malaria is associated with reduced serum retinol levels in rural zambian children. Int. J. Vitam. Nutr. Res. 1998, 68, 384–388. [Google Scholar]
- Shankar, A.H.; Genton, B.; Semba, R.D.; Baisor, M.; Paino, J.; Tamja, S.; Adiguma, T.; Wu, L.; Rare, L.; Tielsch, J.M.; et al. Effect of vitamin A supplementation on morbidity due to Plasmodium falciparum in young children in papua new guinea: A randomised trial. Lancet 1999, 354, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Krishnan, A.D.; Mustafa, A.S.; Talwar, G.P.; Ramalingaswami, V. Effect of vitamin A and undernutrition on the susceptibility of rodents to a malarial parasite Plasmodium berghei. J. Nutr. 1976, 106, 784–791. [Google Scholar] [CrossRef] [PubMed]
- Stoltzfus, R.J.; Jalal, F.; Harvey, P.W.; Nesheim, M.C. Interactions between vitamin A deficiency and Plasmodium berghei infection in the rat. J. Nutr. 1989, 119, 2030–2037. [Google Scholar] [CrossRef] [PubMed]
- Villamor, E.; Mbise, R.; Spiegelman, D.; Hertzmark, E.; Fataki, M.; Peterson, K.E.; Ndossi, G.; Fawzi, W.W. Vitamin A supplements ameliorate the adverse effect of HIV-1, malaria, and diarrheal infections on child growth. Pediatrics 2002, 109, E6. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, L.; Musumeci, S. The immune response to Plasmodium falciparum malaria. Lancet Infect. Dis. 2002, 2, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, L.; Pignatelli, S.; Simpore, J.; Malaguarnera, M.; Musumeci, S. Plasma levels of interleukin-12 (IL-12), interleukin-18 (IL-18) and transforming growth factor beta (TGF-beta) in Plasmodium falciparum malaria. Eur. Cytokine Netw. 2002, 13, 425–430. [Google Scholar]
- Smith, T.G.; Serghides, L.; Patel, S.N.; Febbraio, M.; Silverstein, R.L.; Kain, K.C. CD36-mediated nonopsonic phagocytosis of erythrocytes infected with stage I and IIa gametocytes of plasmodium falciparum. Infect. Immun. 2003, 71, 393–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Checkley, W.; Epstein, L.D.; Gilman, R.H.; Black, R.E.; Cabrera, L.; Sterling, C.R. Effects of Cryptosporidium parvum infection in peruvian children: Growth faltering and subsequent catch-up growth. Am. J. Epidemiol. 1998, 148, 497–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, P.; Kleinova, T.; Volek, Z.; Simunek, J. Effect of Cryptosporidium parvum infection on the absorptive capacity and paracellular permeability of the small intestine in neonatal calves. Vet. Parasitol. 2008, 152, 53–59. [Google Scholar] [CrossRef]
- Goodgame, R.W.; Kimball, K.; Ou, C.N.; White, A.C., Jr.; Genta, R.M.; Lifschitz, C.H.; Chappell, C.L. Intestinal function and injury in acquired immunodeficiency syndrome-related cryptosporidiosis. Gastroenterology 1995, 108, 1075–1082. [Google Scholar] [CrossRef]
- Kirkpatrick, B.D.; Daniels, M.M.; Jean, S.S.; Pape, J.W.; Karp, C.; Littenberg, B.; Fitzgerald, D.W.; Lederman, H.M.; Nataro, J.P.; Sears, C.L. Cryptosporidiosis stimulates an inflammatory intestinal response in malnourished haitian children. J. Infect. Dis. 2002, 186, 94–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carman, J.A.; Pond, L.; Nashold, F.; Wassom, D.L.; Hayes, C.E. Immunity to Trichinella spiralis infection in vitamin A-deficient mice. J. Exp. Med. 1992, 175, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.A.; Cannons, J.L.; Grainger, J.R.; Dos Santos, L.M.; Hand, T.W.; Naik, S.; Wohlfert, E.A.; Chou, D.B.; Oldenhove, G.; Robinson, M.; et al. Essential role for retinoic acid in the promotion of CD4+ T cell effector responses via retinoic acid receptor alpha. Immunity 2011, 34, 435–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.G.; Wang, C.; Matsumoto, S.; Kim, C.H. High and low vitamin A therapies induce distinct FOXP3+ T-cell subsets and effectively control intestinal inflammation. Gastroenterology 2009, 137, 1391–1402.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.A.; Grainger, J.R.; Spencer, S.P.; Belkaid, Y. The role of retinoic acid in tolerance and immunity. Immunity 2011, 35, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Semba, R.D. The role of vitamin A and related retinoids in immune function. Nutr. Rev. 1998, 56, S38–S48. [Google Scholar] [CrossRef] [PubMed]
- Frieden, T.R.; Sowell, A.L.; Henning, K.J.; Huff, D.L.; Gunn, R.A. Vitamin A levels and severity of measles. New york city. Am. J. Dis. Child. 1992, 146, 182–186. [Google Scholar] [CrossRef]
- Surman, S.L.; Rudraraju, R.; Sealy, R.; Jones, B.; Hurwitz, J.L. Vitamin A deficiency disrupts vaccine-induced antibody-forming cells and the balance of IgA/IgG isotypes in the upper and lower respiratory tract. Viral Immunol. 2012, 25, 341–344. [Google Scholar] [CrossRef] [Green Version]
- Surman, S.L.; Jones, B.G.; Rudraraju, R.; Sealy, R.E.; Hurwitz, J.L. Intranasal administration of retinyl palmitate with a respiratory virus vaccine corrects impaired mucosal IgA response in the vitamin A-deficient host. Clin. Vaccine Immunol. 2014, 21, 598–601. [Google Scholar] [CrossRef] [Green Version]
- Surman, S.L.; Jones, B.G.; Sealy, R.E.; Rudraraju, R.; Hurwitz, J.L. Oral retinyl palmitate or retinoic acid corrects mucosal IgA responses toward an intranasal influenza virus vaccine in vitamin A deficient mice. Vaccine 2014, 32, 2521–2524. [Google Scholar] [CrossRef] [Green Version]
- Biesalski, H.K.; Stofft, E.; Wellner, U.; Niederauer, U.; Bassler, K.H. Vitamin A and ciliated cells. I. Respiratory epithelia. Z. Ernahr. 1986, 25, 114–122. [Google Scholar] [CrossRef]
- Penkert, R.R.; Smith, A.P.; Hrincius, E.R.; McCullers, J.A.; Vogel, P.; Smith, A.M.; Hurwitz, J.L. Effect of vitamin A deficiency in dysregulating immune responses to influenza virus and increasing mortality rates after bacterial coinfections. J. Infect. Dis. 2021, 223, 1806–1816. [Google Scholar] [CrossRef]
- Hurwitz, J.L.; Jones, B.G.; Penkert, R.R.; Gansebom, S.; Sun, Y.; Tang, L.; Bramley, A.M.; Jain, S.; McCullers, J.A.; Arnold, S.R. Low retinol-binding protein and vitamin D levels are associated with severe outcomes in children hospitalized with lower respiratory tract infection and Respiratory Syncytial Virus or human metapneumovirus detection. J. Pediatr. 2017, 187, 323–327. [Google Scholar] [CrossRef]
- Neuzil, K.M.; Gruber, W.C.; Chytil, F.; Stahlman, M.T.; Engelhardt, B.; Graham, B.S. Serum vitamin A levels in Respiratory Syncytial Virus infection. J. Pediatr. 1994, 124, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.J.; Niu, Q.L.; Zhu, X.L.; Zhao, X.Z.; Yang, W.W.; Wang, X.J. Relationship between deficiencies in vitamin A and E and occurrence of infectious diseases among children. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 5009–5012. [Google Scholar] [PubMed]
- Markowitz, L.E.; Nzilambi, N.; Driskell, W.J.; Sension, M.G.; Rovira, E.Z.; Nieburg, P.; Ryder, R.W. Vitamin A levels and mortality among hospitalized measles patients, kinshasa, zaire. J. Trop. Pediatr. 1989, 35, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Hussey, G.D.; Klein, M. A randomized, controlled trial of vitamin A in children with severe measles. N. Engl. J. Med. 1990, 323, 160–164. [Google Scholar] [CrossRef]
- Jensen, K.J.; Fisker, A.B.; Andersen, A.; Sartono, E.; Yazdanbakhsh, M.; Aaby, P.; Erikstrup, C.; Benn, C.S. The effects of vitamin A supplementation with measles vaccine on leucocyte counts and in vitro cytokine production. Br. J. Nutr. 2016, 115, 619–628. [Google Scholar] [CrossRef] [Green Version]
- Barclay, A.J.; Foster, A.; Sommer, A. Vitamin A supplements and mortality related to measles: A randomised clinical trial. Br. Med. J. 1987, 294, 294–296. [Google Scholar] [CrossRef] [Green Version]
- Ellison, J.B. Intensive vitamin therapy in measles. Br. Med. J. 1932, 2, 708–711. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Xie, X.; Tu, Z.; Fu, J.; Xu, D.; Zhou, Y. The signal pathways and treatment of cytokine storm in covid-19. Signal Transduct. Target. Ther. 2021, 6, 255. [Google Scholar] [CrossRef]
- Lai, Y.J.; Chang, H.S.; Yang, Y.P.; Lin, T.W.; Lai, W.Y.; Lin, Y.Y.; Chang, C.C. The role of micronutrient and immunomodulation effect in the vaccine era of COVID-19. J. Chin. Med. Assoc. 2021, 84, 821–826. [Google Scholar] [CrossRef]
- Gasmi, A.; Tippairote, T.; Mujawdiya, P.K.; Peana, M.; Menzel, A.; Dadar, M.; Gasmi Benahmed, A.; Bjorklund, G. Micronutrients as immunomodulatory tools for COVID-19 management. Clin. Immunol. 2020, 220, 108545. [Google Scholar] [CrossRef]
- Zhong, J.C.; Huang, D.Y.; Yang, Y.M.; Li, Y.F.; Liu, G.F.; Song, X.H.; Du, K. Upregulation of angiotensin-converting enzyme 2 by all-trans retinoic acid in spontaneously hypertensive rats. Hypertension 2004, 44, 907–912. [Google Scholar] [CrossRef] [Green Version]
- Rubin, L.P.; Ross, A.C.; Stephensen, C.B.; Bohn, T.; Tanumihardjo, S.A. Metabolic effects of inflammation on vitamin A and carotenoids in humans and animal models. Adv. Nutr. 2017, 8, 197–212. [Google Scholar] [CrossRef]
- McGill, J.L.; Kelly, S.M.; Guerra-Maupome, M.; Winkley, E.; Henningson, J.; Narasimhan, B.; Sacco, R.E. Vitamin A deficiency impairs the immune response to intranasal vaccination and rsv infection in neonatal calves. Sci. Rep. 2019, 9, 15157. [Google Scholar] [CrossRef] [Green Version]
- Schäffer, M.W.; Roy, S.S.; Mukherjee, S.; Ong, D.E.; Das, S.K. Uptake of all-trans retinoic acid-containing aerosol by inhalation to lungs in a guinea pig model system—A pilot study. Exp. Lung Res. 2010, 36, 593–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imdad, A.; Mayo-Wilson, E.; Herzer, K.; Bhutta, Z.A. Vitamin A supplementation for preventing morbidity and mortality in children from six months to five years of age. Cochrane Database Syst. Rev. 2017, 3, CD008524. [Google Scholar]
- Wang, Y.; Gao, Y.; Liu, Q.; Zhan, X.; Li, Z.; Hu, H.; Li, T.; Chen, J. Effect of vitamin A and zinc supplementation on indices of vitamin A status, haemoglobin level and defecation of children with persistent diarrhea. J. Clin. Biochem. Nutr. 2016, 59, 58–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reifen, R.; Mor, A.; Nyska, A. Vitamin A deficiency aggravates rotavirus infection in CD-1 mice through extensive involvement of the gut. Int. J. Vitam. Nutr. Res. 2004, 74, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Cha, H.R.; Chang, S.Y.; Chang, J.H.; Kim, J.O.; Yang, J.Y.; Kim, C.H.; Kweon, M.N. Downregulation of Th17 cells in the small intestine by disruption of gut flora in the absence of retinoic acid. J. Immunol. 2010, 184, 6799–6806. [Google Scholar] [CrossRef] [Green Version]
- Long, K.Z.; Garcia, C.; Santos, J.I.; Rosado, J.L.; Hertzmark, E.; Dupont, H.L.; Ko, G. Vitamin A supplementation has divergent effects on norovirus infections and clinical symptoms among mexican children. J. Infect. Dis. 2007, 196, 978–985. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Ko, G. Antiviral effect of vitamin A on norovirus infection via modulation of the gut microbiome. Sci. Rep. 2016, 6, 25835. [Google Scholar] [CrossRef] [Green Version]
- Bitetto, D.; Bortolotti, N.; Falleti, E.; Vescovo, S.; Fabris, C.; Fattovich, G.; Cussigh, A.; Cmet, S.; Fornasiere, E.; Ceriani, E.; et al. Vitamin A deficiency is associated with hepatitis C virus chronic infection and with unresponsiveness to interferon-based antiviral therapy. Hepatology 2013, 57, 925–933. [Google Scholar] [CrossRef]
- Peres, W.A.; Chaves, G.V.; Goncalves, J.C.; Ramalho, A.; Coelho, H.S. Vitamin A deficiency in patients with hepatitis C virus-related chronic liver disease. Br. J. Nutr. 2011, 106, 1724–1731. [Google Scholar] [CrossRef] [Green Version]
- Mateos-Munoz, B.; Devesa-Medina, M.J.; Matia-Martin, M.P.; Torrejon, M.J.; Suarez, A.; Larrad-Sainz, A.; Rey-Diaz-Rubio, E.; Cardenas, M.C.; Ortega-Medina, L.; Ladero, J.M. The relation of fibrosis stage with nutritional deficiencies and bioelectrical impedance analysis of body composition in patients with chronic hepatitis c. Ann. Hepatol. 2016, 15, 492–500. [Google Scholar]
- Loguercio, C.; Federico, A. Oxidative stress in viral and alcoholic hepatitis. Free. Radic. Biol. Med. 2003, 34, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bocher, W.O.; Wallasch, C.; Hohler, T.; Galle, P.R. All-trans retinoic acid for treatment of chronic hepatitis C. Liver Int. Off. J. Int. Assoc. Study Liver 2008, 28, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Thavagnanam, S.; Parker, J.C.; McBrien, M.E.; Skibinski, G.; Heaney, L.G.; Shields, M.D. Effects of IL-13 on mucociliary differentiation of pediatric asthmatic bronchial epithelial cells. Pediatr. Res. 2011, 69, 95–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sher, R.; Shulman, G.; Baily, P.; Politzer, W.M. Serum trace elements and vitamin A in leprosy subtypes. Am. J. Clin. Nutr. 1981, 34, 1918–1924. [Google Scholar] [CrossRef] [Green Version]
- Aibana, O.; Franke, M.F.; Huang, C.C.; Galea, J.T.; Calderon, R.; Zhang, Z.; Becerra, M.C.; Smith, E.R.; Ronnenberg, A.G.; Contreras, C.; et al. Impact of vitamin A and carotenoids on the risk of tuberculosis progression. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2017, 65, 900–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenforde, M.W.; Yadav, A.; Dowdy, D.W.; Gupte, N.; Shivakoti, R.; Yang, W.T.; Mwelase, N.; Kanyama, C.; Pillay, S.; Samaneka, W.; et al. Vitamin A and D deficiencies associated with incident tuberculosis in HIV-infected patients initiating antiretroviral therapy in multinational case-cohort study. J. Acquir. Immune Defic. Syndr. 2017, 75, e71–e79. [Google Scholar] [CrossRef]
- Rwangabwoba, J.M.; Fischman, H.; Semba, R.D. Serum vitamin A levels during tuberculosis and human immunodeficiency virus infection. Int. J. Tuberc. Lung Dis. Off. J. Int. Union Against Tuberc. Lung Dis. 1998, 2, 771–773. [Google Scholar]
- Greenstein, R.J.; Su, L.; Shahidi, A.; Brown, W.D.; Clifford, A.; Brown, S.T. Unanticipated Mycobacterium tuberculosis complex culture inhibition by immune modulators, immune suppressants, a growth enhancer, and vitamins A and D: Clinical implications. Int. J. Infect. Dis. Off. Publ. Int. Soc. Infect. Dis. 2014, 26, 37–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baum, M.; Cassetti, L.; Bonvehi, P.; Shor-Posner, G.; Lu, Y.; Sauberlich, H. Inadequate dietary intake and altered nutrition status in early HIV-1 infection. Nutrition 1994, 10, 16–20. [Google Scholar]
- Semba, R.D.; Caiaffa, W.T.; Graham, N.M.; Cohn, S.; Vlahov, D. Vitamin A deficiency and wasting as predictors of mortality in human immunodeficiency virus-infected injection drug users. J. Infect. Dis. 1995, 171, 1196–1202. [Google Scholar] [CrossRef]
- Camp, W.L.; Allen, S.; Alvarez, J.O.; Jolly, P.E.; Weiss, H.L.; Phillips, J.F.; Karita, E.; Serufilira, A.; Vermund, S.H. Serum retinol and HIV-1 RNA viral load in rapid and slow progressors. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. Off. Publ. Int. Retrovirol. Assoc. 1998, 18, 401–406. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, K.; Groopman, J.E.; Byrn, R.A. The regulation of hiv by retinoic acid correlates with cellular expression of the retinoic acid receptors. Aids 1994, 8, 1675–1682. [Google Scholar] [CrossRef] [PubMed]
- Hussey, G.; Hughes, J.; Potgieter, S.; Kossew, G.; Burgess, J.; Beatty, D.M.; Keraan, A.E.C. Vitamin A status and supplementation and its effect on immunity in children with AIDs. In Report of the xvii International Vitamin a Consultative Group Meeting, Guatemala City, Guatemala; International Life Sciences Institute: Washington, DC, USA, 1996; p. 81. [Google Scholar]
- Fawzi, W.W.; Msamanga, G.I.; Spiegelman, D.; Urassa, E.J.; McGrath, N.; Mwakagile, D.; Antelman, G.; Mbise, R.; Herrera, G.; Kapiga, S.; et al. Randomised trial of effects of vitamin supplements on pregnancy outcomes and T cell counts in HIV-1-infected women in Tanzania. Lancet 1998, 351, 1477–1482. [Google Scholar] [CrossRef] [PubMed]
- Fawzi, W.W.; Msamanga, G.I.; Spiegelman, D.; Wei, R.; Kapiga, S.; Villamor, E.; Mwakagile, D.; Mugusi, F.; Hertzmark, E.; Essex, M.; et al. A randomized trial of multivitamin supplements and hiv disease progression and mortality. N. Engl. J. Med. 2004, 351, 23–32. [Google Scholar] [CrossRef]
- Lietman, T.M.; Dhital, S.P.; Dean, D. Conjunctival impression cytology for vitamin A deficiency in the presence of infectious trachoma. Br. J. Ophthalmol. 1998, 82, 1139–1142. [Google Scholar] [CrossRef] [Green Version]
- Blodi, B.A.; Byrne, K.A.; Tabbara, K.F. Goblet cell population among patients with inactive trachoma. Int. Ophthalmol. 1988, 12, 41–45. [Google Scholar] [CrossRef]
- Cantorna, M.T.; Hayes, C.E. Vitamin A deficiency exacerbates murine lyme arthritis. J. Infect. Dis. 1996, 174, 747–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, A.; Meidovsky, A.; Leitner, G.; Sklan, D. Decreased resistance and immune response to Escherichia coli infection in chicks with low or high intakes of vitamin A. J. Nutr. 1991, 121, 395–400. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Li, Y.; Li, M.; Jia, G.; Dong, H.; Zhang, Y.; He, C.; Wang, C.; Deng, L.; Yang, Y. Hypovitaminosis A coupled to secondary bacterial infection in beef cattle. BMC Vet. Res. 2012, 8, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.M.; Mahalanabis, D.; Hossain, S.; Wahed, M.A.; Alvarez, J.O.; Siber, G.R.; Thompson, C.; Santosham, M.; Fuchs, G.J. Simultaneous vitamin A administration at routine immunization contact enhances antibody response to diphtheria vaccine in infants younger than six months. J. Nutr. 1999, 129, 2192–2195. [Google Scholar] [CrossRef] [Green Version]
- Bhaskaram, P.; Rao, K.V. Enhancement in seroconversion to measles vaccine with simultaneous administration of vitamin a in 9-months-old indian infants. Indian J. Pediatr. 1997, 64, 503–509. [Google Scholar] [CrossRef]
- Rudraraju, R.; Surman, S.L.; Jones, B.G.; Sealy, R.; Woodland, D.L.; Hurwitz, J.L. Reduced frequencies and heightened cd103 expression among virus-induced cd8(+) t cells in the respiratory tract airways of vitamin a-deficient mice. Clin. Vaccine Immunol. 2012, 19, 757–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penkert, R.R.; Cortez, V.; Karlsson, E.A.; Livingston, B.; Surman, S.L.; Li, Y.; Catharine Ross, A.; Schultz-Cherry, S.; Hurwitz, J.L. Vitamin A corrects tissue deficits in diet-induced obese mice and reduces influenza infection after vaccination and challenge. Obesity 2020, 28, 1631–1636. [Google Scholar] [CrossRef]
- Surman, S.L.; Penkert, R.R.; Jones, B.G.; Sealy, R.E.; Hurwitz, J.L. Vitamin supplementation at the time of immunization with a cold-adapted influenza virus vaccine corrects poor mucosal antibody responses in mice deficient for vitamins A and D. Clin. Vaccine Immunol. 2016, 23, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Penkert, R.R.; Rowe, H.M.; Surman, S.L.; Sealy, R.E.; Rosch, J.; Hurwitz, J.L. Influences of vitamin A on vaccine immunogenicity and efficacy. Front. Immunol. 2019, 10, 1576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molrine, D.C.; Polk, D.B.; Ciamarra, A.; Phillips, N.; Ambrosino, D.M. Impaired human responses to tetanus toxoid in vitamin A-deficient SCID mice reconstituted with human peripheral blood lymphocytes. Infect. Immun. 1995, 63, 2867–2872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, D.R.; De Calisto, J.; Simmons, N.L.; Cruz, A.N.; Villablanca, E.J.; Mora, J.R.; Barouch, D.H. Vitamin A deficiency impairs vaccine-elicited gastrointestinal immunity. J. Immunol. 2011, 187, 1877–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatun, S.; Tezic, T.; Kunak, B.; Cengiz, A.B. Vitamin A levels of children with measles in ankara, turkey. Turk. J. Pediatr. 1995, 37, 193–200. [Google Scholar] [PubMed]
- Benn, C.S.; Aaby, P.; Bale, C.; Olsen, J.; Michaelsen, K.F.; George, E.; Whittle, H. Randomised trial of effect of vitamin A supplementation on antibody response to measles vaccine in Guinea-Bissau, West Africa. Lancet 1997, 350, 101–105. [Google Scholar] [CrossRef]
- Newton, S.; Owusu-Agyei, S.; Ampofo, W.; Zandoh, C.; Adjuik, M.; Adjei, G.; Tchum, S.; Filteau, S.; Kirkwood, B.R. Vitamin A supplementation enhances infants’ immune responses to hepatitis B vaccine but does not affect responses to Haemophilus influenzae type B vaccine. J. Nutr. 2007, 137, 1272–1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semba, R.D.; Muhilal; Mohgaddam, N.E.; Munasir, Z.; Akib, A.; Permaesih, D.; Muherdiyantiningsih; Osterhaus, A. Integration of vitamin A supplementation with the expanded program on immunization does not affect seroconversion to oral poliovirus vaccine in infants. J. Nutr. 1999, 129, 2203–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Church, J.A.; Rukobo, S.; Govha, M.; Carmolli, M.P.; Diehl, S.A.; Chasekwa, B.; Ntozini, R.; Mutasa, K.; Humphrey, J.H.; Kirkpatrick, B.D.; et al. Neonatal vitamin A supplementation and immune responses to oral polio vaccine in Zimbabwean infants. Trans. R. Soc. Trop. Med. Hyg. 2019, 113, 110–115. [Google Scholar] [CrossRef] [Green Version]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Kinross, J.M.; Darzi, A.W.; Nicholson, J.K. Gut microbiome-host interactions in health and disease. Genome Med. 2011, 3, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Deng, Y.; Chu, Q.; Zhang, P. Gut microbiome and cancer immunotherapy. Cancer Lett. 2019, 447, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Wilks, J.; Golovkina, T. Influence of microbiota on viral infections. PLoS Pathog. 2012, 8, e1002681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hord, N.G. Eukaryotic-microbiota crosstalk: Potential mechanisms for health benefits of prebiotics and probiotics. Annu. Rev. Nutr. 2008, 28, 215–231. [Google Scholar] [CrossRef]
- Pull, S.L.; Doherty, J.M.; Mills, J.C.; Gordon, J.I.; Stappenbeck, T.S. Activated macrophages are an adaptive element of the colonic epithelial progenitor niche necessary for regenerative responses to injury. Proc. Natl. Acad. Sci. USA 2005, 102, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakoff-Nahoum, S.; Medzhitov, R. Regulation of spontaneous intestinal tumorigenesis through the adaptor protein MyD88. Science 2007, 317, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Sirisinha, S. The pleiotropic role of vitamin A in regulating mucosal immunity. Asian Pac. J. Allergy Immunol. 2015, 33, 71–89. [Google Scholar]
- Frasca, L.; Lande, R. Role of defensins and cathelicidin LL37 in auto-immune and auto-inflammatory diseases. Curr. Pharm. Biotechnol. 2012, 13, 1882–1897. [Google Scholar] [CrossRef]
- Besten, G.; Eunen, K.; Groen, A.; Venema, K.; Reijngoud, D.; Bakker, B. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, M.; Thaiss, C.A.; Elinav, E. Metabolites: Messengers between the microbiota and the immune system. Genes Dev. 2016, 30, 1589–1597. [Google Scholar] [CrossRef]
- Bonakdar, M.; Czuba, L.C.; Han, G.; Zhong, G.; Luong, H.; Isoherrannen, N.; Vaishnava, S. Gut commensals expand vitamin A metabolic capacity of the mammalian host. Cell Host Microbe 2022, 30, 1084–1092.e5. [Google Scholar] [CrossRef] [PubMed]
- Hibberd, M.C.; Wu, M.; Rodionov, D.A.; Li, X.; Cheng, J.; Griffin, N.W.; Barratt, M.J.; Giannone, R.J.; Hettich, R.L.; Osterman, A.L.; et al. The effects of micronutrient deficiencies on bacterial species from the human gut microbiota. Sci. Transl. Med. 2017, 9, eaal4069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grizotte-Lake, M.; Zhong, G.; Duncan, K.; Kirkwood, J.; Iyer, N.; Smolenski, I.; Isoherranen, N.; Vaishnava, S. Commensals suppress intestinal epithelial cell retinoic acid synthesis to regulate interleukin-22 activity and prevent microbial dysbiosis. Immunity 2018, 49, 1103–1115.e6. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, A.; Singh, R.K.; Moniaux, N.; El-Metwally, T.H.; Aubert, J.P.; Batra, S.K. Retinoic acid-dependent transforming growth factor-beta 2-mediated induction of MUC4 mucin expression in human pancreatic tumor cells follows retinoic acid receptor-alpha signaling pathway. J. Biol. Chem. 2000, 275, 33929–33936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, J.Y.; Lee, D.K.; Ha, N.J.; Shin, H.S. Antiviral effects of Lactobacillus ruminis SPM0211 and Bifidobacterium longum SPM1205 and SPM1206 on rotavirus-infected Caco-2 cells and a neonatal mouse model. J. Microbiol. 2015, 53, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Takahashi, T.; Oishi, K.; Tanaka, H.; Masuda, M.; Takahashi, S.; Takano, M.; Kawakami, T.; Fukushima, K.; Kanazawa, H.; et al. Consecutive oral administration of Bifidobacterium longum MM-2 improves the defense system against influenza virus infection by enhancing natural killer cell activity in a murine model. Microbiol. Immunol. 2015, 59, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Moriya, T.; Sakai, F.; Ikeda, N.; Shiozaki, T.; Hosoya, T.; Nakagawa, H.; Miyazaki, T. Oral administration of Lactobacillus gasseri SBT2055 is effective for preventing influenza in mice. Sci. Rep. 2014, 4, 4638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaboriau-Routhiau, V.; Rakotobe, S.; Lecuyer, E.; Mulder, I.; Lan, A.; Bridonneau, C.; Rochet, V.; Pisi, A.; De Paepe, M.; Brandi, G.; et al. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity 2009, 31, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Liu, S.; Feng, D.; Xiao, L.; Yang, T.; Li, T.; Sun, W.; Chen, J. Vitamin A deficiency in the early-life periods alters a diversity of the colonic mucosal microbiota in rats. Front. Nutr. 2020, 7, 580780. [Google Scholar] [CrossRef]
- Liu, H.X.; Hu, Y.; Wan, Y.J. Microbiota and bile acid profiles in retinoic acid-primed mice that exhibit accelerated liver regeneration. Oncotarget 2016, 7, 1096–1106. [Google Scholar] [CrossRef]
- Scott, K.P.; Gratz, S.W.; Sheridan, P.O.; Flint, H.J.; Duncan, S.H. The influence of diet on the gut microbiota. Pharmacol. Res. 2013, 69, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lukiw, W.J. Microbiome-generated amyloid and potential impact on amyloidogenesis in alzheimer’s disease (AD). J. Nat. Sci. 2015, 1, e138. [Google Scholar] [PubMed]
- Bercik, P.; Park, A.J.; Sinclair, D.; Khoshdel, A.; Lu, J.; Huang, X.; Deng, Y.; Blennerhassett, P.A.; Fahnestock, M.; Moine, D.; et al. The anxiolytic effect of Bifidobacterium longum NCC3001 involves vagal pathways for gut-brain communication. Neurogastroenterol. Motil. Off. J. Eur. Gastrointest. Motil. Soc. 2011, 23, 1132–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central gaba receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef] [Green Version]
- Doifode, T.; Giridharan, V.V.; Generoso, J.S.; Bhatti, G.; Collodel, A.; Schulz, P.E.; Forlenza, O.V.; Barichello, T. The impact of the microbiota-gut-brain axis on alzheimer’s disease pathophysiology. Pharmacol. Res. 2021, 164, 105314. [Google Scholar] [CrossRef] [PubMed]
- Petra, A.I.; Panagiotidou, S.; Hatziagelaki, E.; Stewart, J.M.; Conti, P.; Theoharides, T.C. Gut-microbiota-brain axis and its effect on neuropsychiatric disorders with suspected immune dysregulation. Clin. Ther. 2015, 37, 984–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huda, M.N.; Lewis, Z.; Kalanetra, K.M.; Rashid, M.; Ahmad, S.M.; Raqib, R.; Qadri, F.; Underwood, M.A.; Mills, D.A.; Stephensen, C.B. Stool microbiota and vaccine responses of infants. Pediatrics 2014, 134, e362–e372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chac, D.; Bhuiyan, T.R.; Saha, A.; Alam, M.M.; Salma, U.; Jahan, N.; Chowdhury, F.; Khan, A.I.; Ryan, E.T.; LaRocque, R.; et al. Gut microbiota and development of Vibrio cholerae-specific long-term memory B cells in adults after whole-cell killed oral cholera vaccine. Infect. Immun. 2021, 89, e0021721. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.H.; Armah, G.; Dennis, F.; Wang, L.; Rodgers, R.; Droit, L.; Baldridge, M.T.; Handley, S.A.; Harris, V.C. Enteric virome negatively affects seroconversion following oral rotavirus vaccination in a longitudinally sampled cohort of ghanaian infants. Cell Host Microbe 2022, 30, 110–123.e5. [Google Scholar] [CrossRef] [PubMed]
- Parker, E.P.K.; Bronowski, C.; Sindhu, K.N.C.; Babji, S.; Benny, B.; Carmona-Vicente, N.; Chasweka, N.; Chinyama, E.; Cunliffe, N.A.; Dube, Q.; et al. Impact of maternal antibodies and microbiota development on the immunogenicity of oral rotavirus vaccine in African, Indian, and European infants. Nat. Commun. 2021, 12, 7288. [Google Scholar] [CrossRef] [PubMed]
- Borey, M.; Blanc, F.; Lemonnier, G.; Leplat, J.J.; Jardet, D.; Rossignol, M.N.; Ravon, L.; Billon, Y.; Bernard, M.; Estelle, J.; et al. Links between fecal microbiota and the response to vaccination against influenza A virus in pigs. NPJ Vaccines 2021, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Nothaft, H.; Perez-Munoz, M.E.; Yang, T.; Murugan, A.V.M.; Miller, M.; Kolarich, D.; Plastow, G.S.; Walter, J.; Szymanski, C.M. Improving chicken responses to glycoconjugate vaccination against Campylobacter jejuni. Front. Microbiol. 2021, 12, 734526. [Google Scholar] [CrossRef]
- Bouskra, D.; Brezillon, C.; Berard, M.; Werts, C.; Varona, R.; Boneca, I.G.; Eberl, G. Lymphoid tissue genesis induced by commensals through NoD1 regulates intestinal homeostasis. Nature 2008, 456, 507–510. [Google Scholar] [CrossRef] [PubMed]
- Hansen, R.; Russell, R.K.; Reiff, C.; Louis, P.; McIntosh, F.; Berry, S.H.; Mukhopadhya, I.; Bisset, W.M.; Barclay, A.R.; Bishop, J.; et al. Microbiota of de-novo peadiatric IBD: Increased Faecalibacterium prausnitzii and reduced bacterial diversity in crohn’s but not in ulcerative colitis. Am. J. Gastroenterol. 2012, 107, 1913–1922. [Google Scholar] [CrossRef] [Green Version]
- Macpherson, A.J.; Harris, N.L. Interactions between commensal intestinal bacteria and the immune system. Nat. Rev. Immunol. 2004, 4, 478–485. [Google Scholar] [CrossRef]
- Magwira, C.A.; Taylor, M.B. Composition of gut microbiota and its influence on the immunogenicity of oral rotavirus vaccines. Vaccine 2018, 36, 3427–3433. [Google Scholar] [CrossRef]
- Ivanov, I.I.; Honda, K. Intestinal commensal microbes as immune modulators. Cell Host Microbe 2012, 12, 496–508. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, E.V.; Perunova, N.B.; Valyshev, A.V.; Valysheva, I.V.; Bukharin, O.V. Species characteristic and factors of persistence of gut bifidoflora during healthy state and dysbiosis. Zhurnal Mikrobiol. Epidemiol. Immunobiol. 2009, 2, 89–93. [Google Scholar]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Nava, G.M.; Stappenbeck, T.S. Diversity of the autochthonous colonic microbiota. Gut Microbes 2011, 2, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Lee, S.M.; Li, J.; Tran, G.; Jabri, B.; Chatila, T.A.; Mazmanian, S.K. The toll-like receptor 2 pathway establishes colonization by a commensal of the human microbiota. Science 2011, 332, 974–977. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. Inducible Foxp3+ regulatory t-cell development by a commensal bacterium of the intestinal microbiota. Proc. Natl. Acad. Sci. USA 2010, 107, 12204–12209. [Google Scholar] [CrossRef] [Green Version]
- Umesaki, Y.; Okada, Y.; Matsumoto, S.; Imaoka, A.; Setoyama, H. Segmented filamentous bacteria are indigenous intestinal bacteria that activate intraepithelial lymphocytes and induce MHC class II molecules and fucosyl asialo GM1 glycolipids on the small intestinal epithelial cells in the ex-germ-free mouse. Microbiol. Immunol. 1995, 39, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Wingender, G.; Stepniak, D.; Krebs, P.; Lin, L.; McBride, S.; Wei, B.; Braun, J.; Mazmanian, S.K.; Kronenberg, M. Intestinal microbes affect phenotypes and functions of invariant natural killer T cells in mice. Gastroenterology 2012, 143, 418–428. [Google Scholar] [CrossRef] [Green Version]
- Harris, V.C.; Armah, G.; Fuentes, S.; Korpela, K.E.; Parashar, U.; Victor, J.C.; Tate, J.; de Weerth, C.; Giaquinto, C.; Wiersinga, W.J.; et al. Significant correlation between the infant gut microbiome and rotavirus vaccine response in rural Ghana. J. Infect. Dis. 2017, 215, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Eloe-Fadrosh, E.A.; McArthur, M.A.; Seekatz, A.M.; Drabek, E.F.; Rasko, D.A.; Sztein, M.B.; Fraser, C.M. Impact of oral typhoid vaccination on the human gut microbiota and correlations with S. typhi-specific immunological responses. PLoS ONE 2013, 8, e62026. [Google Scholar] [CrossRef] [PubMed]
- Chattha, K.S.; Vlasova, A.N.; Kandasamy, S.; Esseili, M.A.; Siegismund, C.; Rajashekara, G.; Saif, L.J. Probiotics and colostrum/milk differentially affect neonatal humoral immune responses to oral rotavirus vaccine. Vaccine 2013, 31, 1916–1923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattha, K.S.; Vlasova, A.N.; Kandasamy, S.; Rajashekara, G.; Saif, L.J. Divergent immunomodulating effects of probiotics on T cell responses to oral attenuated human rotavirus vaccine and virulent human rotavirus infection in a neonatal gnotobiotic piglet disease model. J. Immunol. 2013, 191, 2446–2456. [Google Scholar] [CrossRef] [PubMed]


| Host Model | Pathogen | Key Findings | Reference |
|---|---|---|---|
| Human | Norovirus | VAS reduced MCP-1, IL-8, TNFα and IL-6 fecal levels in children with GII-associated diarrhea. However, VAS increased TNFα and IL-4 levels in children with GI-associated diarrhea. | [90] |
| Human | S. pneumoniae/Asthma | VAS after neonatal S. pneumoniae pneumonia enhanced Foxp3+Treg and Th1 secretion, decreased Th2 and Th17 cell expressions and lessened airway hyperresponsiveness and inflammatory cell infiltration during asthma. | [67] |
| Human | Mycoplasma pneumoniae pneumonia (MPP) | Both severe MPP (sMMP) and nonsevere MPP (nsMMP) led to VAD; however, VA levels were lower in the sMPP than nsMPP serum. | [91] |
| Gnotobiotic pig | Rotavirus (RV) | More severe prolonged diarrhea and increased human RV (HRV) fecal shedding post-challenge in VAD pigs coinciding with lower HRV specific IgA and Abs in serum and intestinal contents reduced HRV-specific IgA ASCs in ileum and duodenum pre- and post-challenge, increased levels of pro-inflammatory cytokine (IL-12) and CD8+ T cells in blood and intestines and lowered anti-inflammatory cytokine (IL-10). Higher numbers of DCs in VAD piglets prechallenge, but these cells decreased markedly postchallenge. Lower frequency of CD103 (integrin αEβ7) expressing DCs. Elevated necrotic MNCs in spleen prechallenge and in intestinal tissues postchallenge. | [16,56,57] |
| Pig | Porcine Epidemic Diarrhea Virus (PEDV) | VAS increased PEDV IgA ASCs and Abs in serum pre-partum and IgA+β7+ (gut homing) B cells in milk post-piglet PEDV challenge. | [92] |
| Rat | Salmonella typhimurium | VAD increased the number of mucosal DCs, IL-12, TLR2 and MyD88 while decreasing IFN-γ and sIgA production. | [93] |
| Mouse | RV | VAD reduced the number of goblet cells per duodenal villi. RV infection caused complete destruction of the villi in VAD mice. Thus, although RV infection and VAD cause few changes alone, together they led to complete damage of the gut mucosal barrier. | [44] |
| Mouse | Enteric infections | Lower T cell frequencies in the gut of VAD mice. C. rodentium infection aggravated inflammation and epithelial hyperplasia in VAD mice; however, RA treatment led to clearance of C. rodentium. | [94] |
| gastrointestinal/respiratory tract infection | Every 10 mg/dL plasma retinol was associated with 18% fewer days of diarrhea with vomiting, 10% fewer days of cough with fever and 6% fewer doctor visits. | [95] | |
| Mouse | lymphocytic choriomeningitis virus | VAD mice showed extreme inflammation and more severe liver pathology, with high deaths during persistent infection. Infected VAD mice had decreased PD-1 but increased cytokine (IFN-γ, TNF-α and IL-2) expression by T cells. | [45] |
| Host Model | Pathogen | Key Findings | Reference |
|---|---|---|---|
| Human | Norovirus | VAS reduced MCP-1, IL-8, TNFα and IL-6 fecal levels in children with GII-associated diarrhea. However, VAS increased TNFα and IL-4 levels in children with GI-associated diarrhea. | [90] |
| Human | S. pneumoniae/Asthma | VAS after neonatal S. pneumoniae pneumonia enhanced Foxp3+Treg and Th1 secretion, decreased Th2 and Th17 cells expressions, lessened airway hyperresponsiveness and inflammatory cell infiltration during asthma. | [67] |
| Human | Mycoplasma pneumoniae pneumonia (MPP) | Both severe MPP (sMMP) and non-severe MPP (nsMMP) led to VAD; however, VA levels were lower in the sMPP than nsMPP serum. | [91] |
| Rat | Salmonella typhimurium | VAD increased the number of mucosal DCs, IL-12, TLR2 and MyD88 while decreasing IFN-γ and sIgA production. | [93] |
| Mouse | Sendai virus | The VAD mice showed aberrant serum Ig isotypes (high IgG2b levels) and cytokine/chemokine patterns (elevated eotaxin), with high frequencies of nephropathy and death. | [38] |
| Mouse | Rotavirus (RV) | VAD reduced the number of goblet cells per duodenal villi. RV infection caused complete destruction of the villi in VAD mice. Thus, although RV infection and VAD cause few changes alone, together they led to complete damage of the gut mucosal barrier. | [44] |
| Mouse | Enteric infections | Lower T cell frequencies in the gut of VAD mice. C. rodentium infection aggravated inflammation and epithelial hyperplasia in VAD mice; however, RA treatment led to clearance of C. rodentium. | [94] |
| gastrointestinal/respiratory tract infection | Every 10 mg/dL plasma retinol was associated with 18% fewer days of diarrhea with vomiting, 10% fewer days of cough with fever and 6% fewer doctor visits. | [95] | |
| Mouse | lymphocytic choriomeningitis virus | VAD mice showed extreme inflammation and more severe liver pathology, with high deaths during persistent infection. Infected VAD mice had decreased PD-1 but increased cytokine (IFN-γ, TNF-α and IL-2) expression by T cells. | [45] |
| Pig | Porcine Epidemic Diarrhea Virus (PEDV) | VAS increased PEDV IgA ASCs and Abs in serum pre-partum and IgA+β7+ (gut homing) B cells in milk post piglet PEDV challenge. | [92] |
| Gn pig | Rotavirus (RV) (human RV vaccines) | Increased levels of IL-8, a pro-inflammatory mediator and decreased IL-10 responses (anti-inflammatory) in vaccinated VAD compared with VAS piglets indicating more severe inflammatory responses in vaccinated VAD piglets post-challenge. Vaccinated VAD pigs had lower serum IgA HRV Ab titers and significantly lower intestinal IgA ASCs post-challenge suggesting lower anamnestic responses | [16,57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amimo, J.O.; Michael, H.; Chepngeno, J.; Raev, S.A.; Saif, L.J.; Vlasova, A.N. Immune Impairment Associated with Vitamin A Deficiency: Insights from Clinical Studies and Animal Model Research. Nutrients 2022, 14, 5038. https://doi.org/10.3390/nu14235038
Amimo JO, Michael H, Chepngeno J, Raev SA, Saif LJ, Vlasova AN. Immune Impairment Associated with Vitamin A Deficiency: Insights from Clinical Studies and Animal Model Research. Nutrients. 2022; 14(23):5038. https://doi.org/10.3390/nu14235038
Chicago/Turabian StyleAmimo, Joshua O., Husheem Michael, Juliet Chepngeno, Sergei A. Raev, Linda J. Saif, and Anastasia N. Vlasova. 2022. "Immune Impairment Associated with Vitamin A Deficiency: Insights from Clinical Studies and Animal Model Research" Nutrients 14, no. 23: 5038. https://doi.org/10.3390/nu14235038
APA StyleAmimo, J. O., Michael, H., Chepngeno, J., Raev, S. A., Saif, L. J., & Vlasova, A. N. (2022). Immune Impairment Associated with Vitamin A Deficiency: Insights from Clinical Studies and Animal Model Research. Nutrients, 14(23), 5038. https://doi.org/10.3390/nu14235038