
Glycated Proteins, Glycine, Acetate, and Monounsaturated Fatty Acids May Act as New Biomarkers to Predict the Progression of Type 2 Diabetes: Secondary Analyses of a Randomized Controlled Trial

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Blood Sampling and Biochemical Routine Measures
2.3. NMR Spectroscopy
2.4. Isolation of Peripheral Blood Mononuclear Cells and RNA
2.5. Nanostring Gene Expression Assay
2.6. Statistics and Bioinformatics Analyses
2.6.1. Tools
2.6.2. Linear Regression Models
2.6.3. Pre-Processing
2.6.4. Stratification of the Participants and Metabolome and Transcriptome Characterization before Intervention
2.6.5. Analyses of Metabolic and Gene Expression Profiles in the Subgroups after Intervention
2.6.6. Gene Set Enrichment Analysis and Competitive Gene Set Testing
3. Results
3.1. Subgroup Characterization before Intervention
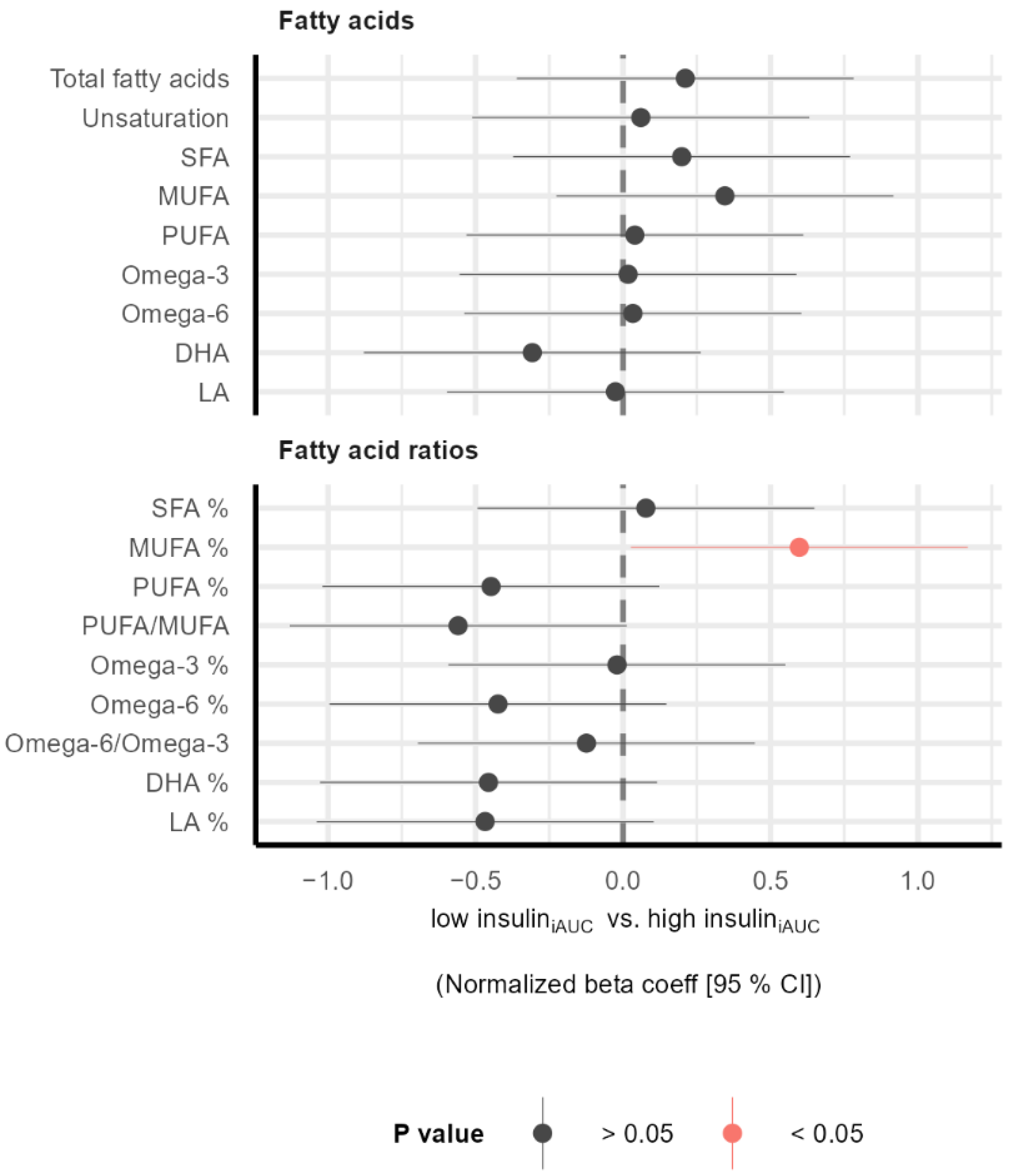
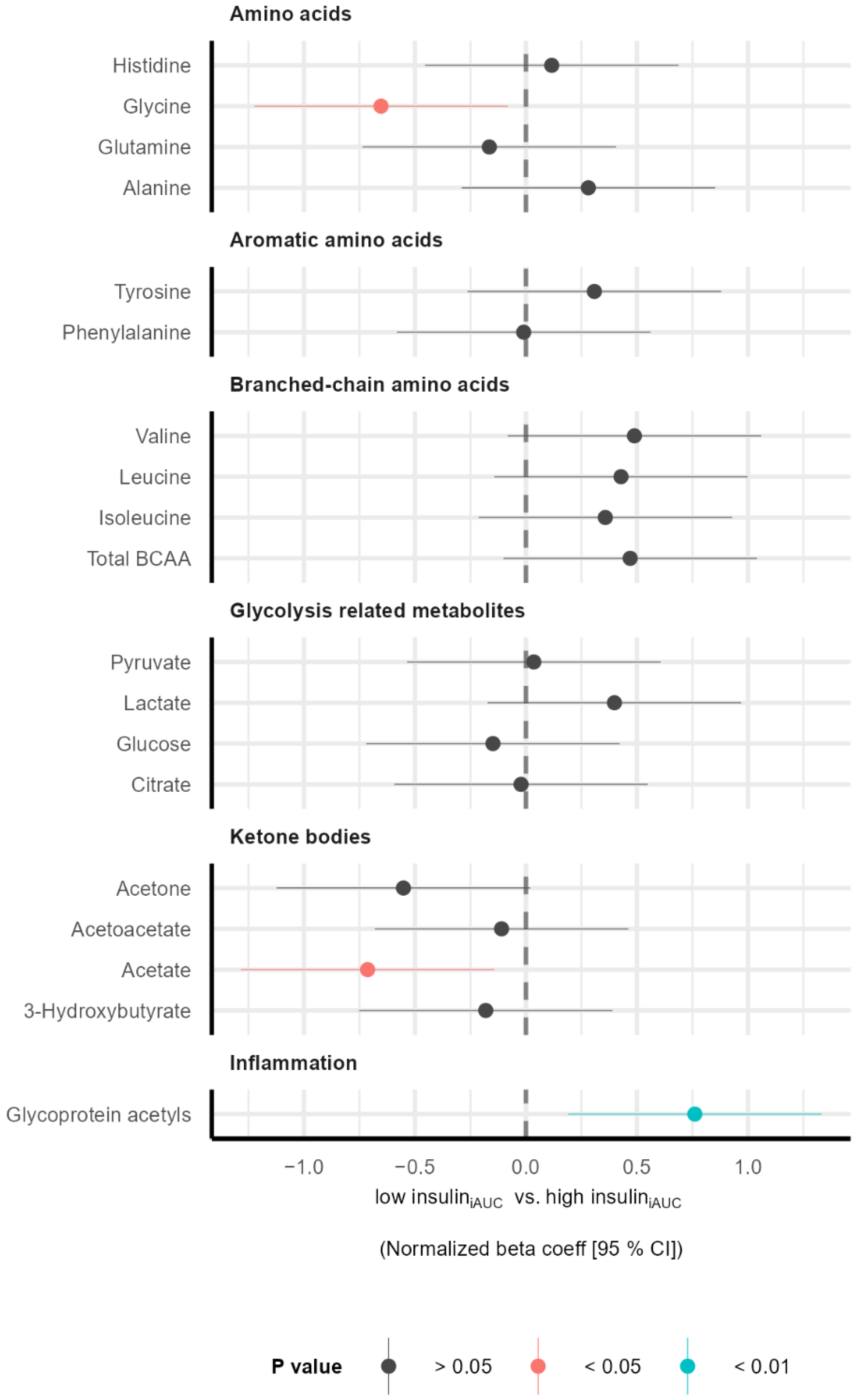
3.2. Subgroup Differences in Metabolic Profile before Intervention
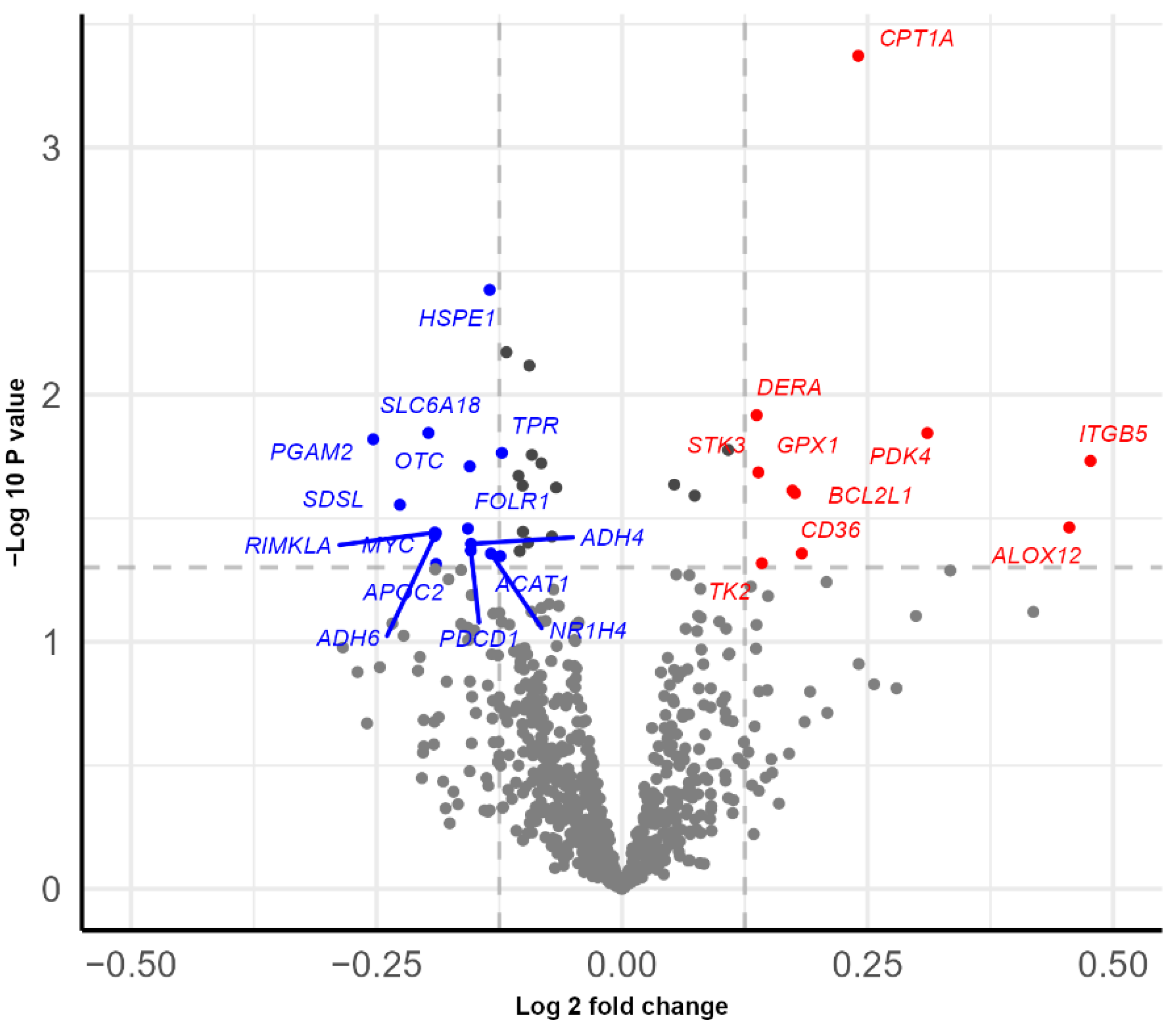
3.3. Subgroup Differences in Gene Expression before Intervention
3.4. Gene Set Enrichment Analysis and Competitive Gene Set Testing before Intervention
3.5. Characterization of the Study Population in the Different Intervention Groups
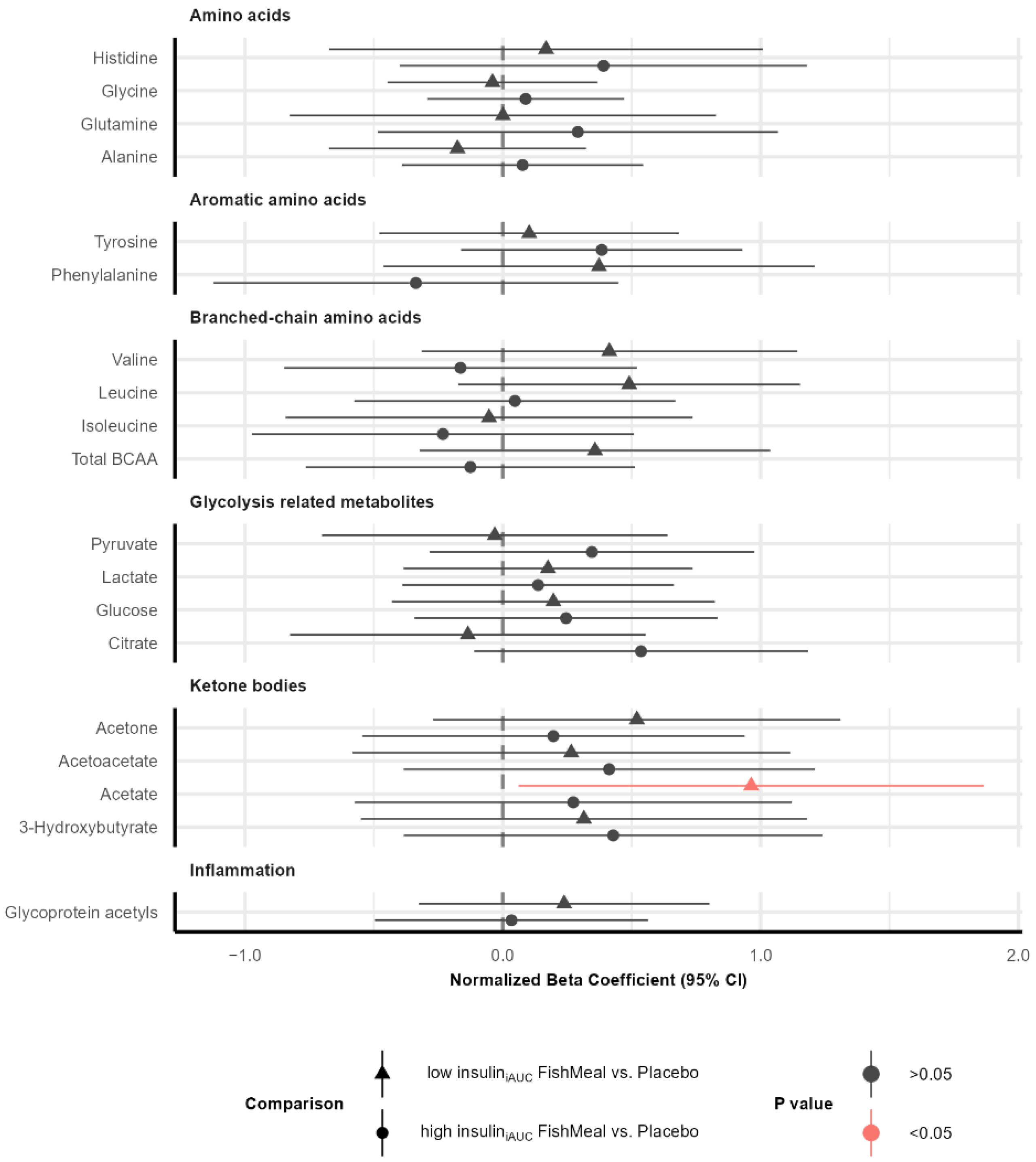
3.6. Effect on Metabolic Profile after Fish Protein Intervention
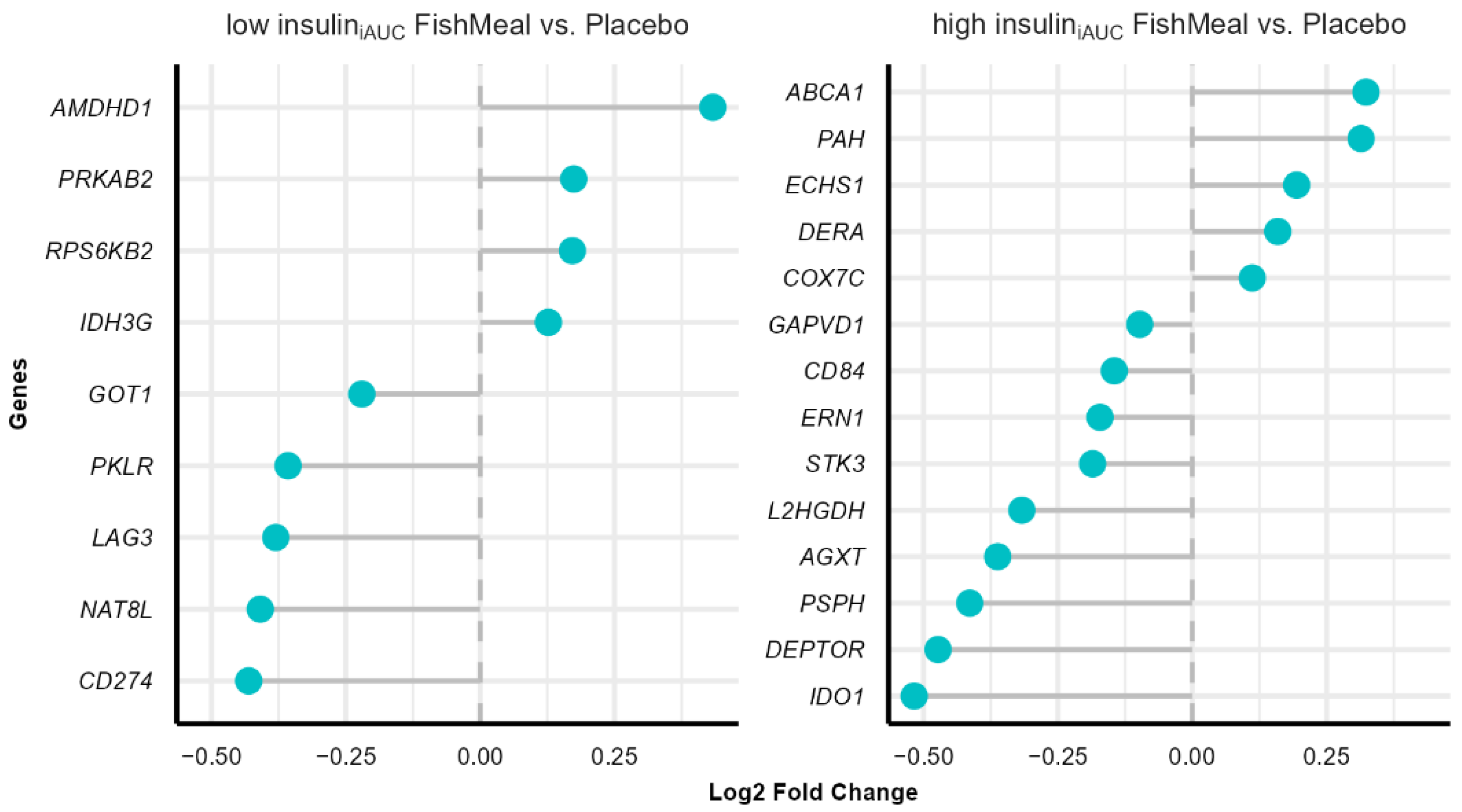
3.7. Effect on Gene Expression after Fish Protein Intervention
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Primers 2015, 1, 15019. [Google Scholar] [CrossRef]
- American Diabetes, A. Diagnosis and classification of diabetes mellitus. Diabetes Care 2013, 36 (Suppl. 1), S67–S74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alok, K.; Gupta, A.M.; Brashear, M.; William, D.J. Chapter 2—Prediabetes: Prevalence, Pathogenesis, and Recognition of Enhanced Risk. In Nutritional and Therapeutic Interventions for Diabetes and Metabolic Syndrome, 2nd ed.; Academic Press: Cambridge, MA, USA, 2018; p. 18. [Google Scholar] [CrossRef]
- Rajala, M.W.; Scherer, P.E. Minireview: The adipocyte--at the crossroads of energy homeostasis, inflammation, and atherosclerosis. Endocrinology 2003, 144, 3765–3773. [Google Scholar] [CrossRef]
- Chung, S.T.; Matta, S.T.; Meyers, A.G.; Cravalho, C.K.; Villalobos-Perez, A.; Dawson, J.M.; Sharma, V.R.; Sampson, M.L.; Otvos, J.D.; Magge, S.N. Nuclear Magnetic Resonance Derived Biomarkers for Evaluating Cardiometabolic Risk in Youth and Young Adults Across the Spectrum of Glucose Tolerance. Front. Endocrinol. 2021, 12, 665292. [Google Scholar] [CrossRef] [PubMed]
- Shalaurova, I.; Connelly, M.A.; Garvey, W.T.; Otvos, J.D. Lipoprotein insulin resistance index: A lipoprotein particle-derived measure of insulin resistance. Metab. Syndr. Relat. Disord. 2014, 12, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Harada, P.H.N.; Demler, O.V.; Dugani, S.B.; Akinkuolie, A.O.; Moorthy, M.V.; Ridker, P.M.; Cook, N.R.; Pradhan, A.D.; Mora, S. Lipoprotein insulin resistance score and risk of incident diabetes during extended follow-up of 20 years: The Women’s Health Study. J. Clin. Lipidol. 2017, 11, 1257–1267.e1252. [Google Scholar] [CrossRef] [Green Version]
- Otvos, J.D.; Shalaurova, I.; Wolak-Dinsmore, J.; Connelly, M.A.; Mackey, R.H.; Stein, J.H.; Tracy, R.P. GlycA: A Composite Nuclear Magnetic Resonance Biomarker of Systemic Inflammation. Clin. Chem. 2015, 61, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Jump, D.B. Fatty acid regulation of gene transcription. Crit. Rev. Clin. Lab. Sci. 2004, 41, 41–78. [Google Scholar] [CrossRef]
- Gonzalez-Becerra, K.; Ramos-Lopez, O.; Barron-Cabrera, E.; Riezu-Boj, J.I.; Milagro, F.I.; Martinez-Lopez, E.; Martinez, J.A. Fatty acids, epigenetic mechanisms and chronic diseases: A systematic review. Lipids Health Dis. 2019, 18, 178. [Google Scholar] [CrossRef] [Green Version]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurtz, P.; Havulinna, A.S.; Soininen, P.; Tynkkynen, T.; Prieto-Merino, D.; Tillin, T.; Ghorbani, A.; Artati, A.; Wang, Q.; Tiainen, M.; et al. Metabolite profiling and cardiovascular event risk: A prospective study of 3 population-based cohorts. Circulation 2015, 131, 774–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, E.A.; Hu, P.J.; Mehta, S. Plasma fatty acids in de novo lipogenesis pathway are associated with diabetogenic indicators among adults: NHANES 2003-2004. Am. J. Clin. Nutr. 2018, 108, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Odegaard, J.I.; Chawla, A. Pleiotropic actions of insulin resistance and inflammation in metabolic homeostasis. Science 2013, 339, 172–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delarue, J.; Magnan, C. Free fatty acids and insulin resistance. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 142–148. [Google Scholar] [CrossRef]
- Sanders, F.W.; Griffin, J.L. De novo lipogenesis in the liver in health and disease: More than just a shunting yard for glucose. Biol. Rev. Camb. Philos. Soc. 2016, 91, 452–468. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.R.; Li, G.W.; Hu, Y.H.; Wang, J.X.; Yang, W.Y.; An, Z.X.; Hu, Z.X.; Lin, J.; Xiao, J.Z.; Cao, H.B.; et al. Effects of diet and exercise in preventing NIDDM in people with impaired glucose tolerance. The Da Qing IGT and Diabetes Study. Diabetes Care 1997, 20, 537–544. [Google Scholar] [CrossRef]
- Li, G.; Zhang, P.; Wang, J.; Gregg, E.W.; Yang, W.; Gong, Q.; Li, H.; Li, H.; Jiang, Y.; An, Y.; et al. The long-term effect of lifestyle interventions to prevent diabetes in the China Da Qing Diabetes Prevention Study: A 20-year follow-up study. Lancet 2008, 371, 1783–1789. [Google Scholar] [CrossRef]
- Micha, R.; Shulkin, M.L.; Penalvo, J.L.; Khatibzadeh, S.; Singh, G.M.; Rao, M.; Fahimi, S.; Powles, J.; Mozaffarian, D. Etiologic effects and optimal intakes of foods and nutrients for risk of cardiovascular diseases and diabetes: Systematic reviews and meta-analyses from the Nutrition and Chronic Diseases Expert Group (NutriCoDE). PLoS ONE 2017, 12, e0175149. [Google Scholar] [CrossRef]
- Nong, N.T.P.; Hsu, J.L. Characteristics of Food Protein-Derived Antidiabetic Bioactive Peptides: A Literature Update. Int. J. Mol. Sci. 2021, 22, 9508. [Google Scholar] [CrossRef]
- Jin, R.; Teng, X.; Shang, J.; Wang, D.; Liu, N. Identification of novel DPP-IV inhibitory peptides from Atlantic salmon (Salmo salar) skin. Food Res. Int. 2020, 133, 109161. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Gong, H.; Mao, X. Dipeptidyl Peptidase-IV Inhibitory Activity and Related Molecular Mechanism of Bovine alpha-Lactalbumin-Derived Peptides. Molecules 2020, 25, 3009. [Google Scholar] [CrossRef] [PubMed]
- Fosgerau, K.; Hoffmann, T. Peptide therapeutics: Current status and future directions. Drug Discov. Today 2015, 20, 122–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, H.F.; Jensen, C.; Hausken, T.; Lied, E.; Hatlebakk, J.G.; Bronstad, I.; Lihaug Hoff, D.A.; Lied, G.A. Effect of a cod protein hydrolysate on postprandial glucose metabolism in healthy subjects: A double-blind cross-over trial. J. Nutr. Sci. 2018, 7, e33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawasaki, T.; Seki, E.; Osajima, K.; Yoshida, M.; Asada, K.; Matsui, T.; Osajima, Y. Antihypertensive effect of valyl-tyrosine, a short chain peptide derived from sardine muscle hydrolyzate, on mild hypertensive subjects. J. Hum. Hypertens 2000, 14, 519–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobile, V.; Duclos, E.; Michelotti, A.; Bizzaro, G.; Negro, M.; Soisson, F. Supplementation with a fish protein hydrolysate (Micromesistius poutassou): Effects on body weight, body composition, and CCK/GLP-1 secretion. Food Nutr. Res. 2016, 60, 29857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vikoren, L.A.; Nygard, O.K.; Lied, E.; Rostrup, E.; Gudbrandsen, O.A. A randomised study on the effects of fish protein supplement on glucose tolerance, lipids and body composition in overweight adults. Br. J. Nutr. 2013, 109, 648–657. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.F.; Li, G.Z.; Peng, H.B.; Zhang, F.; Chen, Y.; Li, Y. Treatment with marine collagen peptides modulates glucose and lipid metabolism in Chinese patients with type 2 diabetes mellitus. Appl. Physiol. Nutr. Metab. 2010, 35, 797–804. [Google Scholar] [CrossRef]
- Ouellet, V.; Marois, J.; Weisnagel, S.J.; Jacques, H. Dietary cod protein improves insulin sensitivity in insulin-resistant men and women: A randomized controlled trial. Diabetes Care 2007, 30, 2816–2821. [Google Scholar] [CrossRef]
- Hovland, I.H.; Leikanger, I.S.; Stokkeland, O.; Waage, K.H.; Mjos, S.A.; Brokstad, K.A.; McCann, A.; Ueland, P.M.; Slizyte, R.; Carvajal, A.; et al. Effects of low doses of fish and milk proteins on glucose regulation and markers of insulin sensitivity in overweight adults: A randomised, double blind study. Eur. J. Nutr. 2020, 59, 1013–1029. [Google Scholar] [CrossRef]
- Nenseter, M.S.; Osterud, B.; Larsen, T.; Strom, E.; Bergei, C.; Hewitt, S.; Holven, K.B.; Hagve, T.A.; Mjos, S.A.; Solvang, M.; et al. Effect of Norwegian fish powder on risk factors for coronary heart disease among hypercholesterolemic individuals. Nutr. Metab. Cardiovasc. Dis. 2000, 10, 323–330. [Google Scholar] [PubMed]
- Jensen, C.; Fjeldheim Dale, H.; Hausken, T.; Hatlebakk, J.G.; Bronstad, I.; Lied, G.A.; Hoff, D.A.L. Supplementation with Low Doses of a Cod Protein Hydrolysate on Glucose Regulation and Lipid Metabolism in Adults with Metabolic Syndrome: A Randomized, Double-Blind Study. Nutrients 2020, 12, 1991. [Google Scholar] [CrossRef] [PubMed]
- Hustad, K.S.; Ottestad, I.; Hjorth, M.; Dalen, K.T.; Saether, T.; Sheikh, N.A.; Stromnes, M.; Ulven, S.M.; Holven, K.B. No effect of salmon fish protein on 2-h glucose in adults with increased risk of type 2 diabetes: A randomised controlled trial. Br. J. Nutr. 2021, 126, 1304–1313. [Google Scholar] [CrossRef] [PubMed]
- Soininen, P.; Kangas, A.J.; Wurtz, P.; Suna, T.; Ala-Korpela, M. Quantitative serum nuclear magnetic resonance metabolomics in cardiovascular epidemiology and genetics. Circ. Cardiovasc. Genet. 2015, 8, 192–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, 13. [Google Scholar] [CrossRef]
- Signorell, A.; Aho, K.; Alfons, A.; Anderegg, N.; Aragon, T.; Arachchige, C.; Arppe, A.; Baddeley, A.; Barton, K.; Bolker, B.; et al. DescTools: Tools for Descriptive Statistics. 2022. Available online: https://cran.r-project.org/package=DescTools (accessed on 22 October 2022).
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Gene Ontology, C. The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Ritchie, S.C.; Wurtz, P.; Nath, A.P.; Abraham, G.; Havulinna, A.S.; Fearnley, L.G.; Sarin, A.P.; Kangas, A.J.; Soininen, P.; Aalto, K.; et al. The Biomarker GlycA Is Associated with Chronic Inflammation and Predicts Long-Term Risk of Severe Infection. Cell Syst. 2015, 1, 293–301. [Google Scholar] [CrossRef] [Green Version]
- Liberzon, A.; Birger, C.; Thorvaldsdottir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Smyth, G.K. Camera: A competitive gene set test accounting for inter-gene correlation. Nucleic Acids Res. 2012, 40, e133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dullaart, R.P.; Gruppen, E.G.; Connelly, M.A.; Otvos, J.D.; Lefrandt, J.D. GlycA, a biomarker of inflammatory glycoproteins, is more closely related to the leptin/adiponectin ratio than to glucose tolerance status. Clin. Biochem. 2015, 48, 811–814. [Google Scholar] [CrossRef]
- Lorenzo, C.; Festa, A.; Hanley, A.J.; Rewers, M.J.; Escalante, A.; Haffner, S.M. Novel Protein Glycan-Derived Markers of Systemic Inflammation and C-Reactive Protein in Relation to Glycemia, Insulin Resistance, and Insulin Secretion. Diabetes Care 2017, 40, 375–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawler, P.R.; Mora, S. Glycosylation Signatures of Inflammation Identify Cardiovascular Risk: Some Glyc It Hot. Circ. Res. 2016, 119, 1154–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connelly, M.A.; Otvos, J.D.; Shalaurova, I.; Playford, M.P.; Mehta, N.N. GlycA, a novel biomarker of systemic inflammation and cardiovascular disease risk. J. Transl. Med. 2017, 15, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, C.Q.; Parnell, L.D.; Smith, C.E.; Guo, T.; Sayols-Baixeras, S.; Aslibekyan, S.; Tiwari, H.K.; Irvin, M.R.; Bender, C.; Fei, D.; et al. Carbohydrate and fat intake associated with risk of metabolic diseases through epigenetics of CPT1A. Am. J. Clin. Nutr. 2020, 112, 1200–1211. [Google Scholar] [CrossRef] [PubMed]
- Koves, T.R.; Ussher, J.R.; Noland, R.C.; Slentz, D.; Mosedale, M.; Ilkayeva, O.; Bain, J.; Stevens, R.; Dyck, J.R.; Newgard, C.B.; et al. Mitochondrial overload and incomplete fatty acid oxidation contribute to skeletal muscle insulin resistance. Cell Metab. 2008, 7, 45–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guasch-Ferre, M.; Hruby, A.; Toledo, E.; Clish, C.B.; Martinez-Gonzalez, M.A.; Salas-Salvado, J.; Hu, F.B. Metabolomics in Prediabetes and Diabetes: A Systematic Review and Meta-analysis. Diabetes Care 2016, 39, 833–846. [Google Scholar] [CrossRef]
- Thalacker-Mercer, A.E.; Ingram, K.H.; Guo, F.; Ilkayeva, O.; Newgard, C.B.; Garvey, W.T. BMI, RQ, diabetes, and sex affect the relationships between amino acids and clamp measures of insulin action in humans. Diabetes 2014, 63, 791–800. [Google Scholar] [CrossRef] [Green Version]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 9, 311–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, N.D.; Stevens, R.D.; Antinozzi, P.A.; Anderson, A.; Bergman, R.N.; Wagenknecht, L.E.; Newgard, C.B.; Bowden, D.W. Metabolomic profile associated with insulin resistance and conversion to diabetes in the Insulin Resistance Atherosclerosis Study. J. Clin. Endocrinol. Metab. 2015, 100, E463–E468. [Google Scholar] [CrossRef] [PubMed]
- Vanweert, F.; Schrauwen, P.; Phielix, E. Role of branched-chain amino acid metabolism in the pathogenesis of obesity and type 2 diabetes-related metabolic disturbances BCAA metabolism in type 2 diabetes. Nutr Diabetes 2022, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef] [Green Version]
- Tulipani, S.; Griffin, J.; Palau-Rodriguez, M.; Mora-Cubillos, X.; Bernal-Lopez, R.M.; Tinahones, F.J.; Corkey, B.E.; Andres-Lacueva, C. Metabolomics-guided insights on bariatric surgery versus behavioral interventions for weight loss. Obesity 2016, 24, 2451–2466. [Google Scholar] [CrossRef]
- Glynn, E.L.; Piner, L.W.; Huffman, K.M.; Slentz, C.A.; Elliot-Penry, L.; AbouAssi, H.; White, P.J.; Bain, J.R.; Muehlbauer, M.J.; Ilkayeva, O.R.; et al. Impact of combined resistance and aerobic exercise training on branched-chain amino acid turnover, glycine metabolism and insulin sensitivity in overweight humans. Diabetologia 2015, 58, 2324–2335. [Google Scholar] [CrossRef] [Green Version]
- Yan-Do, R.; MacDonald, P.E. Impaired “Glycine”-mia in Type 2 Diabetes and Potential Mechanisms Contributing to Glucose Homeostasis. Endocrinology 2017, 158, 1064–1073. [Google Scholar] [CrossRef] [Green Version]
- Gannon, M.C.; Nuttall, J.A.; Nuttall, F.Q. The metabolic response to ingested glycine. Am. J. Clin. Nutr. 2002, 76, 1302–1307. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Ortiz, M.; Medina-Santillan, R.; Martinez-Abundis, E.; von Drateln, C.R. Effect of glycine on insulin secretion and action in healthy first-degree relatives of type 2 diabetes mellitus patients. Horm. Metab. Res. 2001, 33, 358–360. [Google Scholar] [CrossRef]
- Hjorth, M.; Galigniana, N.M.; Ween, O.; Ulven, S.M.; Holven, K.B.; Dalen, K.T.; Saether, T. Postprandial Effects of Salmon Fishmeal and Whey on Metabolic Markers in Serum and Gene Expression in Liver Cells. Nutrients 2022, 14, 1593. [Google Scholar] [CrossRef]
- Yan, L.J. Pathogenesis of chronic hyperglycemia: From reductive stress to oxidative stress. J. Diabetes Res. 2014, 2014, 137919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatti, J.S.; Bhatti, G.K.; Reddy, P.H. Mitochondrial dysfunction and oxidative stress in metabolic disorders—A step towards mitochondria based therapeutic strategies. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Rosselot, C.; Baumel-Alterzon, S.; Li, Y.; Brill, G.; Lambertini, L.; Katz, L.S.; Lu, G.; Garcia-Ocana, A.; Scott, D.K. The many lives of Myc in the pancreatic beta-cell. J. Biol. Chem. 2021, 296, 100122. [Google Scholar] [CrossRef] [PubMed]
- Eizirik, D.L.; Pasquali, L.; Cnop, M. Pancreatic beta-cells in type 1 and type 2 diabetes mellitus: Different pathways to failure. Nat. Rev. Endocrinol. 2020, 16, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Z.; Gerszten, R.E. Metabolomics and Proteomics in Type 2 Diabetes. Circ Res 2020, 126, 1613–1627. [Google Scholar] [CrossRef]
- Cani, P.D.; Lecourt, E.; Dewulf, E.M.; Sohet, F.M.; Pachikian, B.D.; Naslain, D.; De Backer, F.; Neyrinck, A.M.; Delzenne, N.M. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am. J. Clin. Nutr. 2009, 90, 1236–1243. [Google Scholar] [CrossRef] [Green Version]
- Tsutsumi, R.; Yamasaki, Y.; Takeo, J.; Miyahara, H.; Sebe, M.; Bando, M.; Tanba, Y.; Mishima, Y.; Takeji, K.; Ueshima, N.; et al. Long-chain monounsaturated fatty acids improve endothelial function with altering microbial flora. Transl. Res. 2021, 237, 16–30. [Google Scholar] [CrossRef]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Bjorck, I.; Backhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Low Insuliniauc | High Insuliniauc | p Value |
|---|---|---|---|
| n = 24 | n = 24 | ||
| Sex (n female, %) | 17 (71%) | 15 (63%) | |
| Tobacco use daily (n, %) | 2 (9.1%) | 5 (26.3%) | |
| Age (y) | 53.0 (46.3–64.0) | 61.5 (49.5–65.0) | n.s |
| Weight (kg) | 94.9 ± 16.8 | 100.3 ± 19.0 | n.s |
| BMI (kg m−2) | 32.4 (28.9–35.1) | 33.0 (31.0–36.4) | n.s |
| f. Glucose (mmol L−1) | 5.4 (5.0–5.8) | 5.2 (5.1–5.7) | n.s |
| Glucose 2 h (mmol L−1) | 4.7 ± 1.1 | 6.9 ± 1.1 | *** |
| HbA1c (%) | 5.8 ± 0.3 | 5.9 ± 0.3 | n.s |
| f. Insulin (pmol L−1) | 56 (44–92) | 124 (102–196) | *** |
| Insulin 2 h (pmol L−1) | 154 (120–225) | 804 (661–1152) | *** |
| Insulin iAUC (pmol h L−1) | 113 (60–163) | 766 (647–1058) | *** |
| HOMA-IR | 2.4 (1.7–3.7) | 4.9 (3.7–5.8) | *** |
| Matsuda index | 6.9 (4.9–10.7) | 1.8 (1.1–2.1) | *** |
| Triglycerides (mmol L−1) | 1.4 ± 0.7 | 1.6 ± 0.6 | n.s |
| Total cholesterol (mmol L−1) | 5.3 ± 1.3 | 5.0 ± 0.8 | n.s |
| HDL-C (mmol L−1) | 1.4 (1.2–1.8) | 1.2 (1.1–1.4) | n.s |
| LDL-C (mmol L−1) | 3.5 (2.6–4.3) | 3.3 (2.9–4.2) | n.s |
| ApoA1 (g L−1) | 1.7 (1.5–1.8) | 1.5 (1.4–1.7) | n.s |
| Apo B (g L−1) | 1.1 ± 0.3 | 1.0 ± 0.2 | n.s |
| hsCRP (mg L−1) | 3.4 (2.1–4.6) | 4.2 (2.6–7.3) | n.s |
| Systolic BP (mm Hg) | 117 ± 13 | 123 ± 17 | n.s |
| Diastolic BP (mm Hg) | 69 (58–78) | 71 (66–78) | n.s |
| Pathway Name | Ratio | p Value | Genes Symbol | |
|---|---|---|---|---|
| KEGG Pathway ID | ||||
| hsa05216 | Thyroid cancer | 4/12 | 2.00 × 10−3 | MYC, MAPK1, TP53, TPR |
| hsa03010 | Ribosome | 2/2 | 2.45 × 10−3 | RPLP0, RPL23 |
| hsa00071 | Fatty acid degradation | 4/17 | 8.02 × 10−3 | ACAT1, ADH4, ADH6, CPT1A |
| hsa05219 | Bladder cancer | 3/14 | 2.89 × 10−2 | MYC, MAPK1, TP53 |
| GO term ID | ||||
| GO:0019843 | rRNA binding | 3/3 | 1.17 × 10−4 | NPM1, RPLP0, RPL23 |
| GO:0070180 | large ribosomal subunit rRNA binding | 2/2 | 2.45 × 10−3 | RPLP0, RPL23 |
| GO:0001046 | core promoter sequence-specific DNA binding | 3/7 | 3.55 × 10−3 | MYC, NPM1, TP53 |
| GO:0003735 | structural constituent of ribosome | 2/3 | 7.12 × 10−3 | RPLP0, RPL23 |
| GO:0003723 | RNA binding | 7/50 | 9.25 × 10−3 | HSPE1, IMPDH2, NPM1, RPLP0, TP53, TPR, RPL23 |
| Hallmark (MSigDB) Gene Sets | Number of Genes in the SET | Direction | p Value | FDR |
|---|---|---|---|---|
| MYC targets V1 | 22 | Down | 1.18 × 10−5 | 5.92 × 10−4 |
| MYC targets V2 | 6 | Down | 9.73 × 10−4 | 2.43 × 10−2 |
| IL6 JAK STAT3 signaling | 19 | Up | 3.15 × 10−3 | 5.05 × 10−2 |
| Epithelial mesenchymal transition | 11 | Up | 5.12 × 10−3 | 5.05 × 10−2 |
| TGF beta signaling | 4 | Up | 5.63 × 10−3 | 5.05 × 10−2 |
| Wnt beta catenin signaling | 4 | Down | 6.05 × 10−3 | 5.05 × 10−2 |
| TNFA signaling via NFKB | 31 | Up | 1.55 × 10−2 | 1.11 × 10−1 |
| Apoptosis | 21 | Up | 2.63 × 10−2 | 1.64 × 10−1 |
| Protein secretion | 6 | Up | 3.26 × 10−2 | 1.69 × 10−1 |
| Interferon gamma response | 28 | Up | 3.37 × 10−2 | 1.69 × 10−1 |
| Angiogenesis | 2 | Up | 4.46 × 10−2 | 2.03 × 10−1 |
| Low Insuliniauc Placebo | Low Insuliniauc Fish Protein | High Insuliniauc Placebo | High InsuliniAUC Fish Protein | p Value | |
|---|---|---|---|---|---|
| n = 8 | n = 16 | n = 12 | n = 12 | ||
| Sex (n female, %) | 5 (62%) | 12 (75%) | 9 (75%) | 6 (50%) | |
| Tobacco use daily (n, %) | 2 (25%) | 0 (0%) | 2 (10%) | 3 (25%) | |
| Age (y) | 56.9 ± 13.9 | 52.1 ± 10.6 | 59.0 ± 9.2 | 56.1 ± 10.6 | n.s |
| Weight (kg) | 92.9 ± 19.6 | 95.9 ± 15.8 | 97.0 ± 13.9 | 104.0 ± 23.3 | n.s |
| BMI (kg/m2) | 30.2 (28.5–34.1) | 32.8 (29.4–35.7) | 34.1 (31.2–36.0) | 32.9 (30.8–37.6) | n.s |
| f. Glucose (mmol L−1) | 5.8 (5.0–6.2) | 5.2 (5.0–5.6) | 5.3 (5.0–5.7) | 5.2 (5.1–5.8) | n.s |
| Glucose 2 h (mmol L−1) | 4.9 ± 1.1 | 4.7 ± 0.4 | 6.8 ± 1.4 | 6.9 ± 0.8 | **, ### |
| HbA1c (%) | 5.9 (5.7–6.1) | 5.8 (5.7–5.9) | 5.7 (5.6–5.8) | 5.9 (5.7–6.3) | n.s |
| f. Insulin (pmol L−1) | 73 ± 65 | 66 ± 61 | 139 ± 70 | 147 ± 62 | **, ### |
| Insulin 2 h (pmol L−1) | 148 (133–228) | 164 (115–218) | 764 (685–1152) | 861 (661–1171) | ***, ### |
| Insulin iAUC (pmol h L−1) | 116 (73–163) | 113 (53–159) | 766 (647–1058) | 830 (638–1078) | ***, ### |
| HOMA-IR | 2.2 (1.6–3.2) | 2.4 (1.8–3.7) | 4.3 (3.4–8.6) | 5.1 (4.4–7.6) | ### |
| Matsuda index | 6.2 (4.6–11.2) | 6.8 (5.1–8.6) | 1.8 (1.2–2.2) | 1.7 (1.1–2.0) | ***, ### |
| Triglycerides (mmol L−1) | 1.2 ± 0.4 | 1.5 ± 0.4 | 1.6 ± 0.7 | 1.6 ± 0.4 | n.s |
| Total cholesterol (mmol L−1) | 5.0 ± 0.8 | 5.4 ± 1.6 | 4.9 ± 0.8 | 5.2 ± 0.8 | n.s |
| HDL-C (mmol L−1) | 1.5 (1.2–1.8) | 1.4 (1.2–1.8) | 1.3 (1.1–1.5) | 1.2 (1.0–1.3) | n.s |
| LDL-C (mmol L−1) | 3.2 ± 0.6 | 3.7 ± 1.0 | 3.1 ± 0.2 | 3.8 ± 1.0 | n.s |
| ApoA1 (gL−1) | 1.8 (1.5–1.8) | 1.7 (1.5–1.8) | 1.6 (1.4–1.7) | 1.5 (1.4–1.6) | n.s |
| ApoB (gL−1) | 0.9 ± 0.2 | 1.1 ± 0.2 | 1.0 ± 0.2 | 1.1 ± 0.2 | n.s |
| hsCRP (mg L−1) | 4.0 ± 2.4 | 3.5 ± 3.1 | 4.6 ± 3.0 | 5.4 ± 3.1 | n.s |
| Systolic BP (mm Hg) | 119 ± 16 | 116 ± 13 | 124 ± 20 | 122 ± 13 | n.s |
| Diastolic BP (mm Hg) | 68 ± 13 | 70 ± 11 | 70 ± 8 | 71 ± 11 | n.s |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canet, F.; Christensen, J.J.; Victor, V.M.; Hustad, K.S.; Ottestad, I.; Rundblad, A.; Sæther, T.; Dalen, K.T.; Ulven, S.M.; Holven, K.B.; et al. Glycated Proteins, Glycine, Acetate, and Monounsaturated Fatty Acids May Act as New Biomarkers to Predict the Progression of Type 2 Diabetes: Secondary Analyses of a Randomized Controlled Trial. Nutrients 2022, 14, 5165. https://doi.org/10.3390/nu14235165
Canet F, Christensen JJ, Victor VM, Hustad KS, Ottestad I, Rundblad A, Sæther T, Dalen KT, Ulven SM, Holven KB, et al. Glycated Proteins, Glycine, Acetate, and Monounsaturated Fatty Acids May Act as New Biomarkers to Predict the Progression of Type 2 Diabetes: Secondary Analyses of a Randomized Controlled Trial. Nutrients. 2022; 14(23):5165. https://doi.org/10.3390/nu14235165
Chicago/Turabian StyleCanet, Francisco, Jacob J. Christensen, Victor M. Victor, Kristin S. Hustad, Inger Ottestad, Amanda Rundblad, Thomas Sæther, Knut Tomas Dalen, Stine M. Ulven, Kirsten B. Holven, and et al. 2022. "Glycated Proteins, Glycine, Acetate, and Monounsaturated Fatty Acids May Act as New Biomarkers to Predict the Progression of Type 2 Diabetes: Secondary Analyses of a Randomized Controlled Trial" Nutrients 14, no. 23: 5165. https://doi.org/10.3390/nu14235165
APA StyleCanet, F., Christensen, J. J., Victor, V. M., Hustad, K. S., Ottestad, I., Rundblad, A., Sæther, T., Dalen, K. T., Ulven, S. M., Holven, K. B., & Telle-Hansen, V. H. (2022). Glycated Proteins, Glycine, Acetate, and Monounsaturated Fatty Acids May Act as New Biomarkers to Predict the Progression of Type 2 Diabetes: Secondary Analyses of a Randomized Controlled Trial. Nutrients, 14(23), 5165. https://doi.org/10.3390/nu14235165