
Endogenous Androgens Diminish Food Intake and Activation of Orexin A Neurons in Response to Reduced Glucose Availability in Male Rats

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Surgery
2.2. Response of Feeding Behavior to 2DG Administration
2.3. Response of Orexin Neurons in the LH/PFA and Neurons in the Arc to 2DG Administration
2.4. Immunohistochemistry
2.5. Data Analysis and Hormone Assay
2.6. Statistics
3. Results
3.1. Plasma Testosterone Concentration, Daily Food Intake, and Body Weight Change
3.2. Response of Feeding Behavior to 2DG Administration
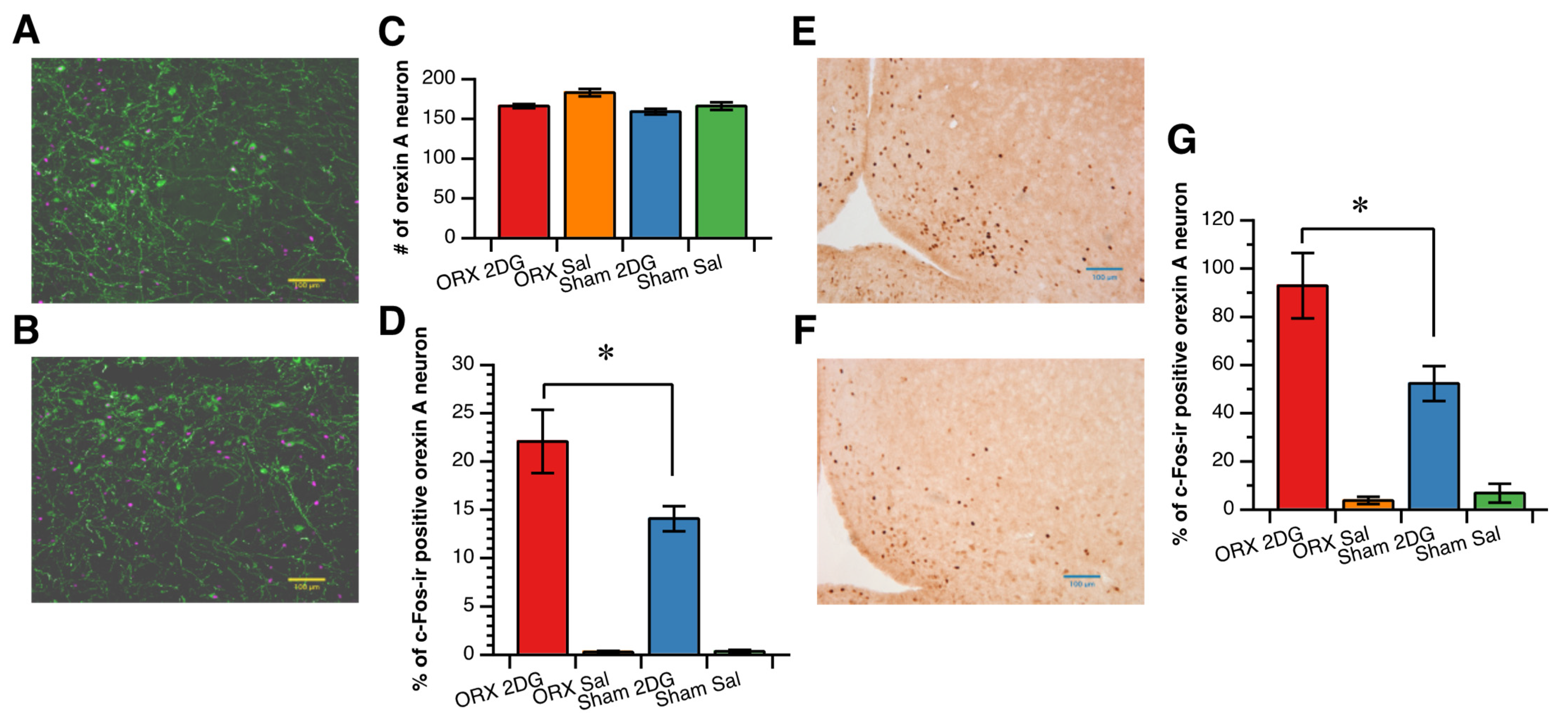
3.3. Hypothalamic Neuronal Response to 2DG Administration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Asarian, L.; Geary, N. Sex Differences in the Physiology of Eating. AJP Regul. Integr. Comp. Physiol. 2013, 305, R1215–R1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asarian, L.; Geary, N. Modulation of Appetite by Gonadal Steroid Hormones. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1251–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschberg, A.L. Sex Hormones, Appetite and Eating Behaviour in Women. Maturitas 2012, 71, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Manson, J.E.; Colditz, G.A.; Stampfer, M.J.; Willett, W.C.; Rosner, B.; Monson, R.R.; Speizer, F.E.; Hennekens, C.H. A Prospective Study of Obesity and Risk of Coronary Heart Disease in Women. N. Engl. J. Med. 1990, 322, 882–889. [Google Scholar] [CrossRef] [PubMed]
- Lobo, R.A. Metabolic Syndrome after Menopause and the Role of Hormones. Maturitas 2008, 60, 10–18. [Google Scholar] [CrossRef]
- Brown, L.M.; Gent, L.; Davis, K.; Clegg, D.J. Metabolic Impact of Sex Hormones on Obesity. Brain Res. 2010, 1350, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Traish, A.M. Testosterone and Weight Loss: The Evidence. Curr. Opin. Endocrinol. Diabetes Obes. 2014, 21, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Kelly, D.M.; Jones, T.H. Testosterone and Obesity. Obes. Rev. 2015, 16, 581–606. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F. Estrogen and Androgen Receptors: Regulators of Fuel Homeostasis and Emerging Targets for Diabetes and Obesity. Trends Endocrinol. Metab. 2011, 22, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Mouser, J.G.; Loprinzi, P.D.; Loenneke, J.P. The Association between Physiologic Testosterone Levels, Lean Mass, and Fat Mass in a Nationally Representative Sample of Men in the United States. Steroids 2016, 115, 62–66. [Google Scholar] [CrossRef]
- Woodhouse, L.J.; Gupta, N.; Bhasin, M.; Singh, A.B.; Ross, R.; Phillips, J.; Bhasin, S. Dose-Dependent Effects of Testosterone on Regional Adipose Tissue Distribution in Healthy Young Men. J. Clin. Endocrinol. Metab. 2004, 89, 718–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baik, M.; Jeong, J.Y.; Park, S.J.; Yoo, S.P.; Lee, J.O.; Lee, J.S.; Haque, M.N.; Lee, H.-J. Testosterone Deficiency Caused by Castration Increases Adiposity in Male Rats in a Tissue-Specific and Diet-Dependent Manner. Genes Nutr. 2020, 15, 14. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Yanase, T.; Nomura, M.; Okabe, T.; Goto, K.; Sato, T.; Kawano, H.; Kato, S.; Nawata, H. Androgen Receptor Null Male Mice Develop Late-Onset Obesity Caused by Decreased Energy Expenditure and Lipolytic Activity but Show Normal Insulin Sensitivity with High Adiponectin Secretion. Diabetes 2005, 54, 1000–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, J.M.; Nunez, A.A.; Siegel, L.I.; Wade, G.N. Effects of Testosterone on Body Weight and Adipose Tissue: Role of Aromatization. Physiol. Behav. 1979, 23, 465–469. [Google Scholar] [CrossRef]
- Sasaki, T. Neural and Molecular Mechanisms Involved in Controlling the Quality of Feeding Behavior: Diet Selection and Feeding Patterns. Nutrients 2017, 9, 1151. [Google Scholar] [CrossRef] [Green Version]
- Brown, L.M.; Clegg, D.J. Central Effects of Estradiol in the Regulation of Food Intake, Body Weight, and Adiposity. J. Steroid Biochem. Mol. Biol. 2010, 122, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Geary, N. Estradiol, CCK and Satiation. Peptides 2001, 22, 1251–1263. [Google Scholar] [CrossRef]
- Eckel, L.A.; Houpt, T.A.; Geary, N. Estradiol Treatment Increases CCK-Induced c-Fos Expression in the Brains of Ovariectomized Rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R1378–R1385. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Horvath, T.L. Cross-Talk between Estrogen and Leptin Signaling in the Hypothalamus. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E817–E826. [Google Scholar] [CrossRef] [Green Version]
- Chai, J.K.; Blaha, V.; Meguid, M.M.; Laviano, A.; Yang, Z.J.; Varma, M. Use of Orchiectomy and Testosterone Replacement to Explore Meal Number-to-Meal Size Relationship in Male Rats. Am. J. Physiol. 1999, 276, R1366–R1373. [Google Scholar] [CrossRef]
- Fukushima, A.; Hagiwara, H.; Fujioka, H.; Kimura, F.; Akema, T.; Funabashi, T. Sex Differences in Feeding Behavior in Rats: The Relationship with Neuronal Activation in the Hypothalamus. Front. Neurosci. 2015, 9, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogi, K.; Funabashi, T.; Mitsushima, D.; Hagiwara, H.; Kimura, F. Sex Difference in the Response of Melanin-Concentrating Hormone Neurons in the Lateral Hypothalamic Area to Glucose, as Revealed by the Expression of Phosphorylated Cyclic Adenosine 3’,5’-Monophosphate Response Element-Binding Protein. Endocrinology 2005, 146, 3325–3333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, Y.; Mabuchi, K.; Taguchi, S.; Ikeda, S.; Aida, E.; Negishi, H.; Takamata, A. Involvement of Orexin-A Neurons but Not Melanin-Concentrating Hormone Neurons in the Short-Term Regulation of Food Intake in Rats. J. Physiol. Sci. 2014, 64, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Muthmainah, M.; Gogos, A.; Sumithran, P.; Brown, R.M. Orexins (Hypocretins): The Intersection between Homeostatic and Hedonic Feeding. J. Neurochem. 2021, 157, 1473–1494. [Google Scholar] [CrossRef]
- Simerly, R.B.; Swanson, L.W.; Chang, C.; Muramatsu, M. Distribution of Androgen and Estrogen Receptor MRNA-containing Cells in the Rat Brain: An In Situ Hybridization Study. J. Comp. Neurol. 1990, 294, 76–95. [Google Scholar] [CrossRef]
- Funabashi, T.; Hagiwara, H.; Mogi, K.; Mitsushima, D.; Shinohara, K.; Kimura, F. Sex Differences in the Responses of Orexin Neurons in the Lateral Hypothalamic Area and Feeding Behavior to Fasting. Neurosci. Lett. 2009, 463, 31–34. [Google Scholar] [CrossRef]
- Hoffman, G.E.; Smith, M.S.; Verbalis, J.G. C-Fos and Related Immediate Early Gene Products as Markers of Activity in Neuroendocrine Systems. Front. Neuroendocr. 1993, 14, 173–213. [Google Scholar] [CrossRef]
- Niimi, M.; Sato, M.; Tamaki, M.; Wada, Y.; Takahara, J.; Kawanishi, K. Induction of Fos Protein in the Rat Hypothalamus Elicited by Insulin-Induced Hypoglycemia. Neurosci. Res. 1995, 23, 361–364. [Google Scholar] [CrossRef]
- Kovács, K.J. Measurement of Immediate-Early Gene Activation- c-Fos and Beyond. J. Neuroendocr. 2008, 20, 665–672. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 4th ed.; Academic Press: Cambridge, UK, 2004; ISBN 9780080474120. [Google Scholar]
- Muschamp, J.W.; Dominguez, J.M.; Sato, S.M.; Shen, R.-Y.; Hull, E.M. A Role for Hypocretin (Orexin) in Male Sexual Behavior. J. Neurosci. 2007, 27, 2837–2845. [Google Scholar] [CrossRef] [Green Version]
- Harter, C.J.L.; Kavanagh, G.S.; Smith, J.T. The Role of Kisspeptin Neurons in Reproduction and Metabolism. J. Endocrinol. 2018, 238, R173–R183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dungan, H.M.; Clifton, D.K.; Steiner, R.A. Minireview: Kisspeptin Neurons as Central Processors in the Regulation of Gonadotropin-Releasing Hormone Secretion. Endocrinology 2006, 147, 1154–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izzi-Engbeaya, C.; Dhillo, W.S. Emerging Roles for Kisspeptin in Metabolism. J. Physiol. 2021, 600, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Orlando, G.; Leone, S.; Ferrante, C.; Chiavaroli, A.; Mollica, A.; Stefanucci, A.; Macedonio, G.; Dimmito, M.P.; Leporini, L.; Menghini, L.; et al. Effects of Kisspeptin-10 on Hypothalamic Neuropeptides and Neurotransmitters Involved in Appetite Control. Molecules 2018, 23, 3071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backholer, K.; Smith, J.T.; Rao, A.; Pereira, A.; Iqbal, J.; Ogawa, S.; Li, Q.; Clarke, I.J. Kisspeptin Cells in the Ewe Brain Respond to Leptin and Communicate with Neuropeptide Y and Proopiomelanocortin Cells. Endocrinology 2010, 151, 2233–2243. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.; Cai, X.J.; Elliott, J.C.; Harrold, J.A. Anabolic Neuropeptides. Physiol. Behav. 2004, 81, 211–222. [Google Scholar] [CrossRef]
- Leibowitz, S.F.; Wortley, K.E. Hypothalamic Control of Energy Balance: Different Peptides, Different Functions. Peptides 2004, 25, 473–504. [Google Scholar] [CrossRef]
- Yamanaka, A.; Kunii, K.; Nambu, T.; Tsujino, N.; Sakai, A.; Matsuzaki, I.; Miwa, Y.; Goto, K.; Sakurai, T. Orexin-Induced Food Intake Involves Neuropeptide Y Pathway. Brain Res. 2000, 859, 404–409. [Google Scholar] [CrossRef]
- Fu, L.-Y.; Pol, A.N. van den Kisspeptin Directly Excites Anorexigenic Proopiomelanocortin Neurons but Inhibits Orexigenic Neuropeptide Y Cells by an Indirect Synaptic Mechanism. J. Neurosci. 2010, 30, 10205–10219. [Google Scholar] [CrossRef] [Green Version]
- Stengel, A.; Wang, L.; Goebel-Stengel, M.; Taché, Y. Centrally Injected Kisspeptin Reduces Food Intake by Increasing Meal Intervals in Mice. Neuroreport 2011, 22, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Thompson, E.L.; Patterson, M.; Murphy, K.G.; Smith, K.L.; Dhillo, W.S.; Todd, J.F.; Ghatei, M.A.; Bloom, S.R. Central and Peripheral Administration of Kisspeptin-10 Stimulates the Hypothalamic-Pituitary-Gonadal Axis. J. Neuroendocr. 2004, 16, 850–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takamata, A.; Nishimura, Y.; Oka, A.; Nagata, M.; Kosugi, N.; Eguchi, S.; Negishi, H.; Morimoto, K. Endogenous Androgens Diminish Food Intake and Activation of Orexin A Neurons in Response to Reduced Glucose Availability in Male Rats. Nutrients 2022, 14, 1235. https://doi.org/10.3390/nu14061235
Takamata A, Nishimura Y, Oka A, Nagata M, Kosugi N, Eguchi S, Negishi H, Morimoto K. Endogenous Androgens Diminish Food Intake and Activation of Orexin A Neurons in Response to Reduced Glucose Availability in Male Rats. Nutrients. 2022; 14(6):1235. https://doi.org/10.3390/nu14061235
Chicago/Turabian StyleTakamata, Akira, Yuri Nishimura, Ayano Oka, Mayuna Nagata, Natsumi Kosugi, Sayaka Eguchi, Hiroko Negishi, and Keiko Morimoto. 2022. "Endogenous Androgens Diminish Food Intake and Activation of Orexin A Neurons in Response to Reduced Glucose Availability in Male Rats" Nutrients 14, no. 6: 1235. https://doi.org/10.3390/nu14061235
APA StyleTakamata, A., Nishimura, Y., Oka, A., Nagata, M., Kosugi, N., Eguchi, S., Negishi, H., & Morimoto, K. (2022). Endogenous Androgens Diminish Food Intake and Activation of Orexin A Neurons in Response to Reduced Glucose Availability in Male Rats. Nutrients, 14(6), 1235. https://doi.org/10.3390/nu14061235