
Meals That Differ in Nutrient Composition and Inflammatory Potential Do Not Differentially Impact Postprandial Circulating Cytokines in Older Adults above a Healthy Weight

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Participants
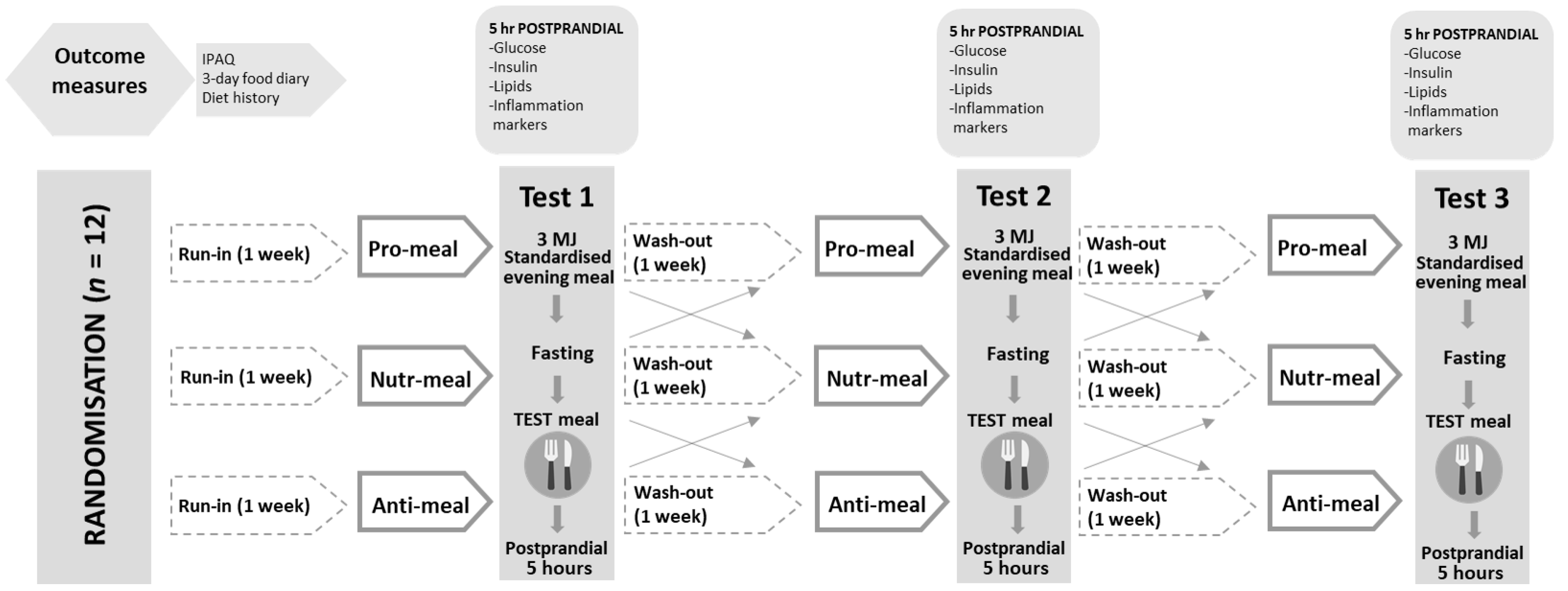
2.2. Study Design and Procedures
2.3. Meal Development and Composition
2.4. Baseline Nutritional Intake and Physical Activity Levels
2.5. Anthropometry, Body Composition, and Blood Pressure
2.6. Inflammatory and Metabolic Markers
2.7. Randomisation, Blinding, and Statistical Analysis
3. Results
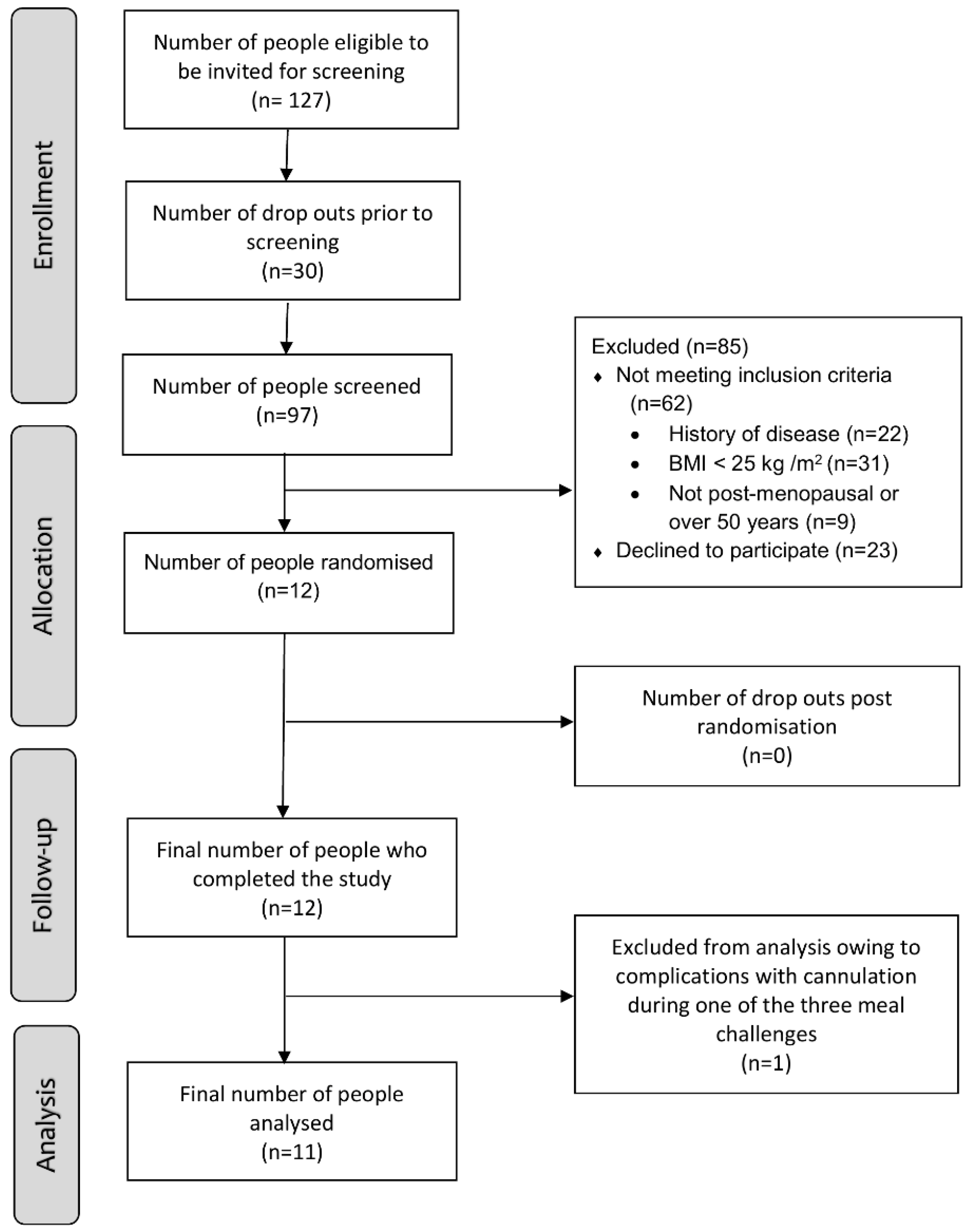
3.1. Participant Enrolment and Characteristics at Baseline
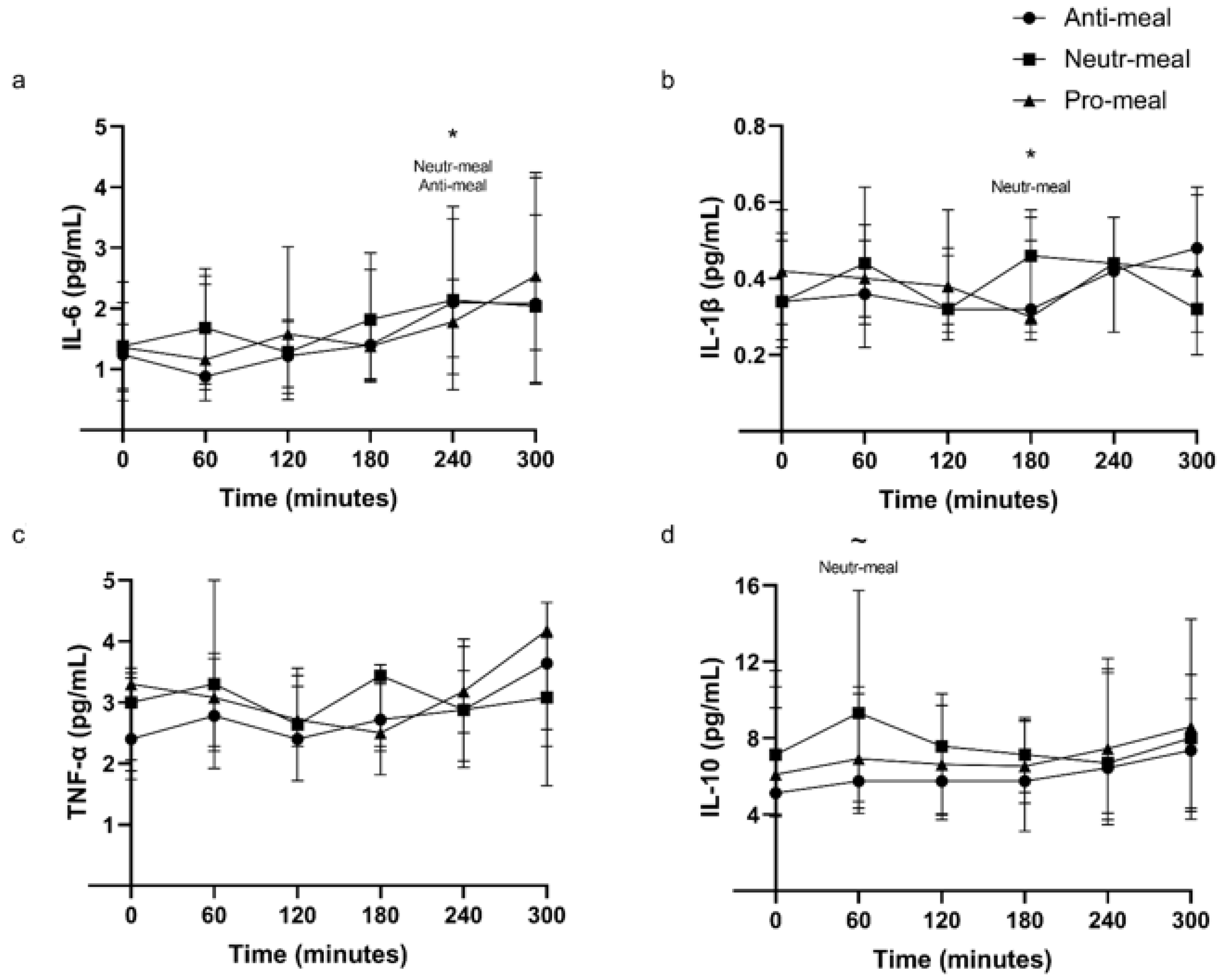
3.2. Plasma Inflammatory Markers
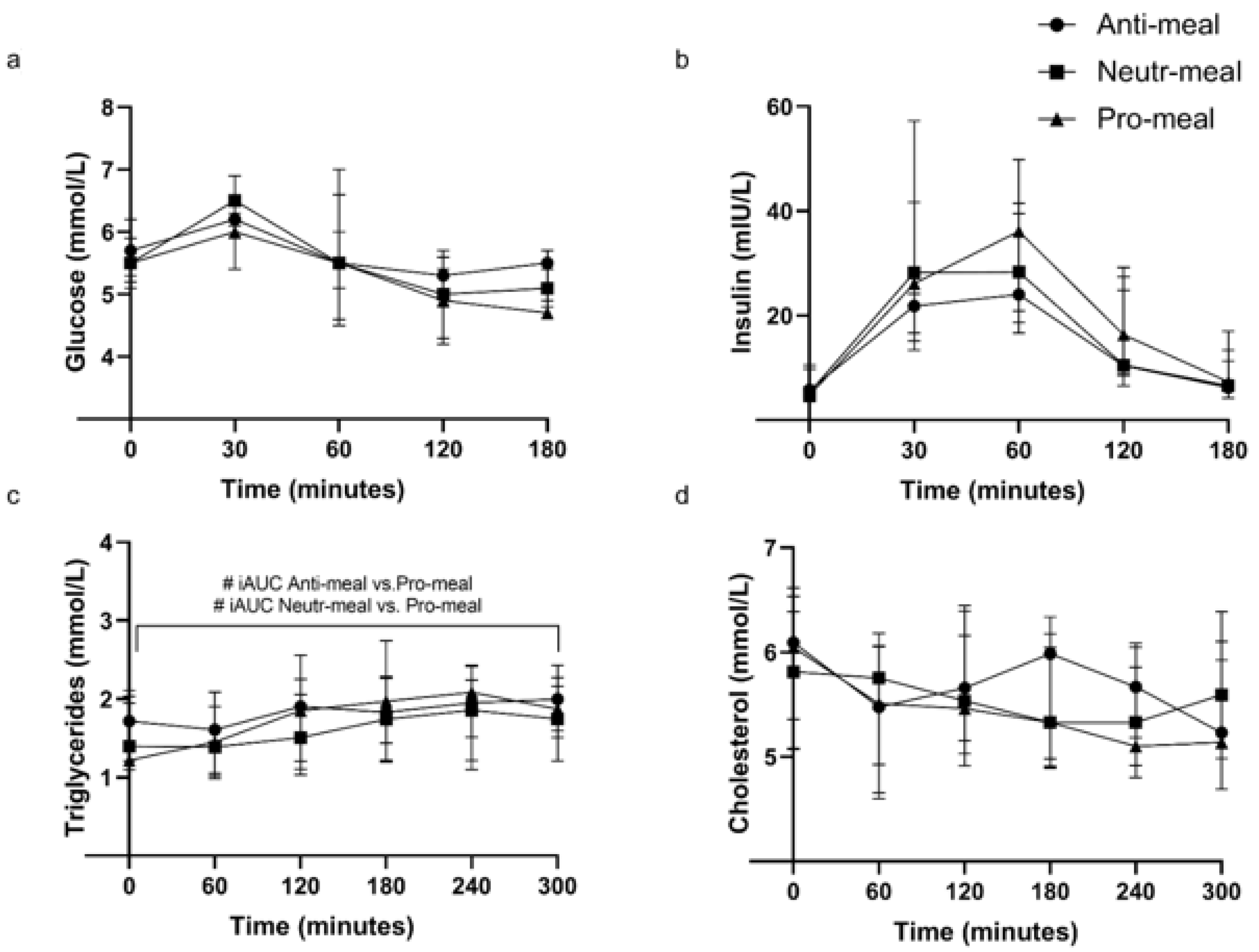
3.3. Plasma Glucose, Insulin, and Serum Lipids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Wild, S.; Roglic, G.; Green, A.; Sicree, R.; King, H. Global prevalence of diabetes: Estimates for the year 2000 and projections for 2030. Diabetes Care 2004, 27, 1047–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Rane, G.; Dai, X.; Shanmugam, M.K.; Arfuso, F.; Samy, R.P.; Lai, M.K.; Kappei, D.; Kumar, A.P.; Sethi, G. Ageing and the telomere connection: An intimate relationship with inflammation. Ageing Res. Rev. 2016, 25, 55–69. [Google Scholar] [CrossRef] [PubMed]
- de Magalhaes, J.P.; Curado, J.; Church, G.M. Meta-analysis of age-related gene expression profiles identifies common signatures of aging. Bioinformatics 2009, 25, 875–881. [Google Scholar] [CrossRef] [Green Version]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Inflammaging: Disturbed interplay between autophagy and inflammasomes. Aging 2012, 4, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C.; Ahluwalia, N.; Brouns, F.; Buetler, T.; Clement, K.; Cunningham, K.; Esposito, K.; Jönsson, L.S.; Kolb, H.; Lansink, M. Dietary factors and low-grade inflammation in relation to overweight and obesity. Br. J. Nutr. 2011, 106, S1–S78. [Google Scholar] [CrossRef]
- Calder, P.C.; Bosco, N.; Bourdet-Sicard, R.; Capuron, L.; Delzenne, N.; Doré, J.; Franceschi, C.; Lehtinen, M.J.; Recker, T.; Salvioli, S. Health relevance of the modification of low grade inflammation in ageing (inflammageing) and the role of nutrition. Ageing Res. Rev. 2017, 40, 95–119. [Google Scholar] [CrossRef]
- Calder, P.C.; Ahluwalia, N.; Albers, R.; Bosco, N.; Bourdet-Sicard, R.; Haller, D.; Holgate, S.T.; Jönsson, L.S.; Latulippe, M.E.; Marcos, A. A consideration of biomarkers to be used for evaluation of inflammation in human nutritional studies. Br. J. Nutr. 2013, 109, S1–S34. [Google Scholar] [CrossRef] [Green Version]
- Cowan, S.F.; Leeming, E.R.; Sinclair, A.; Dordevic, A.L.; Truby, H.; Gibson, S.J. Effect of whole foods and dietary patterns on markers of subclinical inflammation in weight-stable overweight and obese adults: A systematic review. Nutr. Rev. 2019, 79, 19–38. [Google Scholar] [CrossRef]
- Ceriello, A. Postprandial hyperglycemia and diabetes complications: Is it time to treat? Diabetes 2005, 54, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Jackson, K.G.; Poppitt, S.D.; Minihane, A.M. Postprandial lipemia and cardiovascular disease risk: Interrelationships between dietary, physiological and genetic determinants. Atherosclerosis 2012, 220, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Esposito, K.; Nappo, F.; Marfella, R.; Giugliano, G.; Giugliano, F.; Ciotola, M.; Quagliaro, L.; Ceriello, A.; Giugliano, D. Inflammatory cytokine concentrations are acutely increased by hyperglycemia in humans: Role of oxidative stress. Circulation 2002, 106, 2067–2072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Vries, M.A.; Klop, B.; Janssen, H.W.; Njo, T.L.; Westerman, E.M.; Cabezas, M.C. Postprandial Inflammation: Targeting Glucose and Lipids. In Oxidative Stress and Inflammation in Non-Communicable Diseases-Molecular Mechanisms and Perspectives in Therapeutics; Springer: Berlin/Heidelberg, Germany, 2014; pp. 161–170. [Google Scholar]
- Klop, B.; Proctor, S.D.; Mamo, J.C.; Botham, K.M.; Castro Cabezas, M. Understanding postprandial inflammation and its relationship to lifestyle behaviour and metabolic diseases. Int. J. Vasc. Med. 2012, 2012, 947417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nappo, F.; Esposito, K.; Cioffi, M.; Giugliano, G.; Molinari, A.M.; Paolisso, G.; Marfella, R.; Giugliano, D. Postprandial endothelial activation in healthy subjects and in type 2 diabetic patients: Role of fat and carbohydrate meals. J. Am. Coll. Cardiol. 2002, 39, 1145–1150. [Google Scholar] [CrossRef] [Green Version]
- Dali-Youcef, N.; Mecili, M.; Ricci, R.; Andrès, E. Metabolic inflammation: Connecting obesity and insulin resistance. Ann. Med. 2013, 45, 242–253. [Google Scholar] [CrossRef]
- Newsholme, P.; Keane, D.; Welters, H.J.; Morgan, N.G. Life and death decisions of the pancreatic β-cell: The role of fatty acids. Clin. Sci. 2007, 112, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Prentki, M.; Joly, E.; El-Assaad, W.; Roduit, R. Malonyl-CoA signaling, lipid partitioning, and glucolipotoxicity: Role in β-cell adaptation and failure in the etiology of diabetes. Diabetes 2002, 51 (Suppl. S3), S405–S413. [Google Scholar] [CrossRef] [Green Version]
- Emerson, S.R.; Kurti, S.P.; Harms, C.A.; Haub, M.D.; Melgarejo, T.; Logan, C.; Rosenkranz, S.K. Magnitude and Timing of the Postprandial Inflammatory Response to a High-Fat Meal in Healthy Adults: A Systematic Review. Adv. Nutr. 2017, 8, 213–225. [Google Scholar] [CrossRef] [Green Version]
- Herieka, M.; Erridge, C. High-fat meal induced postprandial inflammation. Mol. Nutr. Food Res. 2014, 58, 136–146. [Google Scholar] [CrossRef]
- Mazidi, M.; Valdes, A.M.; Ordovas, J.M.; Hall, W.L.; Pujol, J.C.; Wolf, J.; Hadjigeorgiou, G.; Segata, N.; Sattar, N.; Koivula, R.; et al. Meal-induced inflammation: Postprandial insights from the Personalised REsponses to DIetary Composition Trial (PREDICT) study in 1000 participants. Am. J. Clin. Nutr. 2021, 114, 1028–1038. [Google Scholar] [CrossRef]
- Edirisinghe, I.; Banaszewski, K.; Cappozzo, J.; Sandhya, K.; Ellis, C.L.; Tadapaneni, R.; Kappagoda, C.T.; Burton-Freeman, B.M. Strawberry anthocyanin and its association with postprandial inflammation and insulin. Br. J. Nutr. 2011, 106, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Huebbe, P.; Giller, K.; de Pascual-Teresa, S.; Arkenau, A.; Adolphi, B.; Portius, S.; Arkenau, C.N.; Rimbach, G. Effects of blackcurrant-based juice on atherosclerosis-related biomarkers in cultured macrophages and in human subjects after consumption of a high-energy meal. Br. J. Nutr. 2012, 108, 234–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Gómez, Y.; López-Miranda, J.; Blanco-Colio, L.M.; Marín, C.; Pérez-Martínez, P.; Ruano, J.; Paniagua, J.A.; Rodríguez, F.; Egido, J.; Pérez-Jiménez, F. Olive oil and walnut breakfasts reduce the postprandial inflammatory response in mononuclear cells compared with a butter breakfast in healthy men. Atherosclerosis 2009, 204, e70–e76. [Google Scholar] [CrossRef] [PubMed]
- Ono-Moore, K.D.; Snodgrass, R.G.; Huang, S.; Singh, S.; Freytag, T.L.; Burnett, D.J.; Bonnel, E.L.; Woodhouse, L.R.; Zunino, S.J.; Peerson, J.M. Postprandial Inflammatory Responses and Free Fatty Acids in Plasma of Adults Who Consumed a Moderately High-Fat Breakfast with and without Blueberry Powder in a Randomized Placebo-Controlled Trial. J. Nutr. 2016, 146, 1411–1419. [Google Scholar] [CrossRef]
- Peluso, I.; VVillano, D.; ARoberts, S.; Cesqui, E.; Raguzzini, A.; Borges, G.; Crozier, A.; Catasta, G.; Toti, E.; Serafini, M. Consumption of mixed fruit-juice drink and vitamin C reduces postprandial stress induced by a high fat meal in healthy overweight subjects. Curr. Pharm. Des. 2014, 20, 1020–1024. [Google Scholar] [CrossRef]
- Devaraj, S.; Wang-Polagruto, J.; Polagruto, J.; Keen, C.L.; Jialal, I. High-fat, energy-dense, fast-food–style breakfast results in an increase in oxidative stress in metabolic syndrome. Metabolism 2008, 57, 867–870. [Google Scholar] [CrossRef] [Green Version]
- Milan, A.M.; Pundir, S.; Pileggi, C.A.; Markworth, J.F.; Lewandowski, P.A.; Cameron-Smith, D. Comparisons of the postprandial inflammatory and endotoxaemic responses to mixed meals in young and older individuals: A randomised trial. Nutrients 2017, 9, 354. [Google Scholar] [CrossRef] [Green Version]
- Shivappa, N.; Steck, S.E.; Hurley, T.G.; Hussey, J.R.; Hébert, J.R. Designing and developing a literature-derived, population-based dietary inflammatory index. Public Health Nutr. 2014, 17, 1689–1696. [Google Scholar] [CrossRef] [Green Version]
- Manning, P.J.; Sutherland, W.H.; McGrath, M.M.; De Jong, S.A.; Walker, R.J.; Williams, M.J. Postprandial cytokine concentrations and meal composition in obese and lean women. Obesity 2008, 16, 2046–2052. [Google Scholar] [CrossRef]
- Jacobs, D.R., Jr.; Gross, M.D.; Tapsell, L.C. Food synergy: An operational concept for understanding nutrition. Am. J. Clin. Nutr. 2009, 89, 1543S–1548S. [Google Scholar] [CrossRef]
- Aguilera, J.M. The food matrix: Implications in processing, nutrition and health. Crit. Rev. Food Sci. Nutr. 2019, 59, 3612–3629. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Dartois, A.; Kaur, L. Starch digestibility in food matrix: A review. Trends Food Sci. Technol. 2010, 21, 168–180. [Google Scholar] [CrossRef]
- Lattimer, J.M.; Haub, M.D. Effects of dietary fiber and its components on metabolic health. Nutrients 2010, 2, 1266–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Miao, M. Dietary polyphenols modulate starch digestion and glycaemic level: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 541–555. [Google Scholar] [CrossRef]
- Haack, M.; Kraus, T.; Schuld, A.; Dalal, M.; Koethe, D.; Pollmächer, T. Diurnal variations of interleukin-6 plasma levels are confounded by blood drawing procedures. Psychoneuroendocrinology 2002, 27, 921–931. [Google Scholar] [CrossRef]
- Thompson, D.; Dixon, N. Measurement of postprandial interleukin-6 via a catheter: What does it tell us? Eur. J. Appl. Physiol. 2009, 107, 621–622. [Google Scholar] [CrossRef]
- Agorastos, A.; Hauger, R.L.; Barkauskas, D.A.; Moeller-Bertram, T.; Clopton, P.L.; Haji, U.; Lohr, J.B.; Geracioti, T.D.; Patel, P.M., Jr.; Chrousos, G.P.; et al. Circadian rhythmicity, variability and correlation of interleukin-6 levels in plasma and cerebrospinal fluid of healthy men. Psychoneuroendocrinology 2014, 44, 71–82. [Google Scholar] [CrossRef]
- Vgontzas, A.N.; Bixler, E.O.; Lin, H.M.; Prolo, P.; Trakada, G.; Chrousos, G.P. IL-6 and its circadian secretion in humans. Neuroimmunomodulation 2005, 12, 131–140. [Google Scholar] [CrossRef]
- Gill, S.K.; Allerton, D.M.; Ansley-Robson, P.; Hemmings, K.; Cox, M.; Costa, R.J. Does Short-Term High Dose Probiotic Supplementation Containing Lactobacillus casei Attenuate Exertional-Heat Stress Induced Endotoxaemia and Cytokinaemia? Int. J. Sport Nutr. Exerc. Metab. 2016, 26, 268–275. [Google Scholar] [CrossRef]
- Rocha, D.; Lopes, L.L.; da Silva, A.; Oliveira, L.L.; Bressan, J.; Hermsdorff, H.H.M. Orange juice modulates proinflammatory cytokines after high-fat saturated meal consumption. Food Funct. 2017, 8, 4396–4403. [Google Scholar] [CrossRef]
- Fogarty, C.L.; Nieminen, J.K.; Peraneva, L.; Lassenius, M.I.; Ahola, A.J.; Taskinen, M.R.; Jauhiainen, M.; Kirveskari, J.; Pussinen, P.; Horkko, S.; et al. High-fat meals induce systemic cytokine release without evidence of endotoxemia-mediated cytokine production from circulating monocytes or myeloid dendritic cells. Acta Diabetol. 2015, 52, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Rimmer, M.; Tan, B.K.; Teede, H.; Thangaratinam, S.; Al Wattar, B.H. Metabolic inflexibility in women with polycystic ovary syndrome: A systematic review. Gynecol. Endocrinol. 2020, 36, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Goodpaster, B.H.; Sparks, L.M. Metabolic flexibility in health and disease. Cell Metab. 2017, 25, 1027–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galgani, J.E.; Moro, C.; Ravussin, E. Metabolic flexibility and insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1009–E1017. [Google Scholar] [CrossRef] [Green Version]
- Tchernof, A.; Després, J.-P. Pathophysiology of human visceral obesity: An update. Physiol. Rev. 2013, 93, 359–404. [Google Scholar] [CrossRef]
- Schipper, H.S.; Prakken, B.; Kalkhoven, E.; Boes, M. Adipose tissue-resident immune cells: Key players in immunometabolism. Trends Endocrinol. Metab. 2012, 23, 407–415. [Google Scholar] [CrossRef]
- Akinkuolie, A.O.; Pradhan, A.D.; Buring, J.E.; Ridker, P.M.; Mora, S. Novel protein glycan side-chain biomarker and risk of incident type 2 diabetes mellitus. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1544–1550. [Google Scholar] [CrossRef] [Green Version]
- Duprez, D.A.; Otvos, J.; Sanchez, O.A.; Mackey, R.H.; Tracy, R.; Jacobs, D.R., Jr. Comparison of the Predictive Value of GlycA and Other Biomarkers of Inflammation for Total Death, Incident Cardiovascular Events, Noncardiovascular and Noncancer Inflammatory-Related Events, and Total Cancer Events. Clin. Chem. 2016, 62, 1020–1031. [Google Scholar] [CrossRef] [Green Version]
- Kaikkonen, J.E.; Wurtz, P.; Suomela, E.; Lehtovirta, M.; Kangas, A.J.; Jula, A.; Mikkila, V.; Viikari, J.S.; Juonala, M.; Ronnemaa, T.; et al. Metabolic profiling of fatty liver in young and middle-aged adults: Cross-sectional and prospective analyses of the Young Finns Study. Hepatology 2017, 65, 491–500. [Google Scholar] [CrossRef]
- Bondia-Pons, I.; Ryan, L.; Martinez, J.A. Oxidative stress and inflammation interactions in human obesity. J. Physiol. Biochem. 2012, 68, 701–711. [Google Scholar] [CrossRef]
- Bloomer, R.J.; Kabir, M.M.; Marshall, K.E.; Canale, R.E.; Farney, T.M. Postprandial oxidative stress in response to dextrose and lipid meals of differing size. Lipids Health Dis. 2010, 9, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanim, H.; Abuaysheh, S.; Sia, C.L.; Korzeniewski, K.; Chaudhuri, A.; Fernandez-Real, J.M.; Dandona, P. Increase in plasma endotoxin concentrations and the expression of Toll-like receptors and suppressor of cytokine signaling-3 in mononuclear cells after a high-fat, high-carbohydrate meal: Implications for insulin resistance. Diabetes Care 2009, 32, 2281–2287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanim, H.; Sia, C.L.; Korzeniewski, K.; Lohano, T.; Abuaysheh, S.; Marumganti, A.; Chaudhuri, A.; Dandona, P. A resveratrol and polyphenol preparation suppresses oxidative and inflammatory stress response to a high-fat, high-carbohydrate meal. J. Clin. Endocrinol. Metab. 2011, 96, 1409–1414. [Google Scholar] [CrossRef] [Green Version]
- Biobaku, F.; Ghanim, H.; Batra, M.; Dandona, P. Macronutrient-Mediated Inflammation and Oxidative Stress: Relevance to Insulin Resistance, Obesity, and Atherogenesis. J. Clin. Endocrinol. Metab. 2019, 104, 6118–6128. [Google Scholar] [CrossRef] [PubMed]
- Chaves, D.F.S.; Solis, M.Y.; Gandin, P.; Benatti, F.B.; Rodrigues, V.L.; Paschoal, V.; Naves, A.; Lancha, A.H., Jr. Acute effects of isocaloric meals with different fiber and antioxidant contents on inflammatory markers in healthy individuals. Ann. Nutr. Metab. 2013, 62, 164–168. [Google Scholar] [CrossRef]
- Dong, H.; Rendeiro, C.; Kristek, A.; Sargent, L.J.; Saunders, C.; Harkness, L.; Rowland, I.; Jackson, K.G.; Spencer, J.P.; Lovegrove, J.A. Addition of orange pomace to orange juice attenuates the increases in peak glucose and insulin concentrations after sequential meal ingestion in men with elevated cardiometabolic risk. J. Nutr. 2016, 146, 1197–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food Standards Australia New Zealand. Australian Food Composition Database. 2019. Available online: https://www.foodstandards.gov.au/science/monitoringnutrients/afcd/Pages/default.aspx (accessed on 20 February 2020).
- U.S. Department of Agriculture. USDA Database for the Flavonoid Content of Selected Foods. 2016. Available online: https://data.nal.usda.gov/dataset/usda-database-flavonoid-content-selected-foods-release-32-november-2015 (accessed on 20 February 2020).
- Rothwell, J.A.; Perez-Jimenez, J.; Neveu, V.; Medina-Remon, A.; M’hiri, N.; García-Lobato, P.; Manach, C.; Knox, C.; Eisner, R.; Wishart, D.S.; et al. Phenol-Explorer 3.0: A major update of the Phenol-Explorer database to incorporate data on the effects of food processing on polyphenol content. Database 2013, 2013, bat070. [Google Scholar] [CrossRef]
- Australian Bureau of Statistics. 4802.0—National Nutrition Survey: Selected Highlights, Australia, 1995. 1997. Available online: https://www.abs.gov.au/AUSSTATS/[email protected]/DetailsPage/4802.01995?OpenDocument (accessed on 20 February 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Pro-Meal 1 | Neutr-Meal 2 | Anti-Meal 3 |
|---|---|---|---|
| Energy, kJ kcal | 2280 545 | 2150 514 | 2335 558 |
| % Fat, % kJ | 64 | 25 | 35 |
| % Carbohydrate, % kJ | 19 | 33 | 36 |
| % Protein, % kJ | 17 | 34 | 19 |
| Fat, % kJ | 39.3 | 14.5 | 21.9 |
| Saturated fat, g | 20.4 | 6.2 | 3.4 |
| Monounsaturated fat, g | 13.7 | 5.3 | 9.4 |
| Polyunsaturated fat, g | 2.50 | 1.46 | 7.18 |
| Carbohydrate, g | 25.2 | 42.8 | 53.1 |
| Fibre, g | 1.96 | 15.5 | 19.0 |
| Protein, g | 22.2 | 43.1 | 25.7 |
| DII score 4 | +9.36 | −2.76 | −6.24 |
| Variable | Total Population (n = 11) | Male (n = 5) | Female (n = 6) | Male vs. Female p-Value 2 |
|---|---|---|---|---|
| Waist circumference, cm | 101 (95–104) | 101 (97–110) | 98 (90–107) | 0.465 |
| Waist-to-hip ratio | 0.92 (0.88–0.97) | 0.94 (0.89–1.00) | 0.89 (0.86–0.97) | 0.462 |
| Total body fat, % | 37 (30–43) | 30 (27–36) | 42 (36–47) | 0.028 |
| Total visceral fat, g | 1285 (1120–2168) | 1285 (1179–2425) | 1553 (766–2387) | 0.715 |
| Systolic blood pressure, mm Hg | 125 (120–137) | 125 (122–140) | 127 (117–132) | 0.409 |
| Diastolic blood pressure, mm Hg | 78 (76–85) | 77 (74–86) | 80 (75–86) | 0.855 |
| Plasma glucose, mmol/L | 5.50 (5.25–6.00) | 5.70 (5.50–6.20) | 5.35 (5.05–5.73) | 0.014 |
| Plasma insulin, mIU/L | 5.30 (3.85–10.35) | 5.30 (3.20–10.50) | 5.50 (3.88–9.40) | 0.885 |
| Insulin resistance (HOMA-IR) | 1.25 (0.94–2.73) | 1.25 (0.85–3.20) | 1.20 (0.97–2.40) | 0.828 |
| Serum TC, mmol/L | 6.04 (5.19–6.56) | 5.46 (5.08–6.17) | 6.18 (5.80–6.95) | 0.023 |
| Serum LDL, mmol/L | 4.02 (3.48–4.55) | 3.69 (3.29–4.29) | 4.15 (3.70–4.57) | 0.065 |
| Serum HDL, mmol/L | 1.23 (1.05–1.76) | 1.06 (0.96–1.26) | 1.41 (1.19–1.84) | 0.004 |
| Serum TC/HDL ratio | 4.65 (3.18–5.22) | 4.65 (3.12–5.05) | 4.46 (3.14–5.31) | 0.800 |
| Serum TAG, mmol/L | 1.44 (1.18–2.03) | 1.40 (1.21–1.95) | 1.56 (1.13–2.20) | 0.745 |
| Plasma CRP, mg/L | 1.00 (0.70–1.90) | 0.70 (0.50–2.75) | 1.05 (0.85–1.70) | 0.313 |
| Plasma IL-6, pg/mL | 1.36 (0.64–1.92) | 0.64 (0.44–1.24) | 1.64 (1.32–2.58) | 0.001 |
| Plasma IL-1β, pg/mL | 0.38 (0.25–0.51) | 0.28 (0.18–0.44) | 0.48 (0.34–0.54) | 0.019 |
| Plasma TNF-α, pg/mL | 3.00 (1.96–3.48) | 2.08 (1.74–3.00) | 3.35 (2.76–3.52) | 0.014 |
| Plasma IL-10, pg/mL | 6.04 (3.98–10.15) | 4.90 (3.66–8.02) | 6.88 (4.67–13.58) | 0.051 |
| Comorbidities, n (%) Hypertension Hypercholesterolemia | 6 (55) 3 (27) | 4 (67) 1 (17) | 2 (33) 2 (33) | - - |
| Medication use, n (%) Antihypertensive Statins/fibrates Anticoagulants | 6 (55) 3 (27) 2 (18) | 4 (67) 1 (17) 1 (17) | 2 (33) 2 (33) 1 (17) | - - - |
| Variable 2 | Total Population (n = 11) | Male (n = 5) | Female (n = 6) | p-Value 3 |
|---|---|---|---|---|
| Energy, kJ | 9169 (7230–10,448) | 9169 (7413–10,503) | 8903 (7212–10,502) | 1.000 |
| Protein, g % energy | 95.8 (87.2–117.2) 20.2 (18.2–21.0) | 100.4 (96.5–123.9) 21.0 (18.6–22.5) | 90.0 (79.6–109.6) 19.9 (14.6–21.1) | 0.100 0.361 |
| Fat, g % energy | 85.0 (76.2–116.8) 39.4 (37.0–40.9) | 76.8 (68.3–112.8) 37.0 (31.3–39.7) | 98.2 (77.1–127.3) 40.0 (39.3–45.2) | 0.201 0.045 |
| Saturated fat, g | 31.2 (24.7–36.5) | 24.7 (21.0–40.2) | 32.6 (25.5–37.4) | 0.584 |
| MUFA, g | 33.2 (29.8–52.4) | 33.2 (27.6–48.1) | 42.4 (31.6–56.8) | 0.273 |
| PUFA, g | 14.7 (12.5–19.1) | 14.2 (11.7–15.1) | 17.3 (12.0–23.4) | 0.361 |
| Omega-3 fat, g | 0.24 (0.13–0.75) | 0.31 (0.18–0.74) | 0.17 (0.12–1.17) | 0.465 |
| Omega-6 fat, g | 12.3 (10.3–17.0) | 11.4 (9.4–12.8) | 15.2 (9.9–19.0) | 0.201 |
| Trans fat, g | 1.17 (0.99–1.55) | 1.05 (0.97–1.90) | 1.24 (0.97–1.60) | 0.855 |
| Carbohydrate, g % energy | 194.9 (147.2–225.9) 34.0 (30.6–38.2) | 208.3 (148.2–241.5) 34.0 (31.1–41.5) | 190.4 (143.3–230.4) 36.0 (30.0–38.7) | 0.715 0.855 |
| Sugar, g | 73.3 (49.8–87.7) | 73.3 (53.1–92.9) | 73.7 (48.8–87.4) | 0.584 |
| Fibre, g | 25.2 (21.7–33.9) | 25.2 (23.1–33.0) | 25.4 (19.5–36.2) | 0.715 |
| Sodium, mg | 2268.7 (1937.7–3056.0) | 2268.7 (1783.3–3048.7) | 2170.3 (1721.9–3744.7) | 1.000 |
| Vegetable serves 4 | 3.80 (3.10–3.93) | 3.71 (3.13–3.82) | 3.91 (2.90–5.37) | 0.273 |
| Fruit serves 4 | 0.74 (0.37–1.20) | 0.74 (0.15–2.81) | 0.66 (0.38–0.99) | 0.855 |
| Grain serves 4 Wholegrain serves Refined grain serves | 5.73 (5.18–6.97) 2.51 (0.95–4.06) 3.80 (2.56–6.00) | 6.51 (5.44–7.88) 2.51 (1.34–3.38) 4.00 (2.14–6.43) | 5.32 (5.06–7.78) 2.47 (0.84–4.66) 3.20 (2.20–5.06) | 0.465 0.855 0.361 |
| Total MET-min/week n (%) inactive (IPAQ 1) n (%) minimally active (IPAQ 2) n (%) HEPA active (IPAQ 3) | 2079 (1152–4586) 1 (9) 5 (45) 5 (45) | 2079 (807–5340) 1 (17) 2 (33) 3 (50) | 3018 (1563–5025) 0 (0) 3 (50) 3 (50) | 0.855 - - - |
| Outcome 1 | Anti-Meal | Neutr-Meal | Pro-Meal | p-Value 2 |
|---|---|---|---|---|
| IL-6 iAUC | 70.8 (23.6–267.6) | 59.2 (27.9–177.2) | 78.8 (6.3–138.3) | 0.695 |
| IL-1β iAUC | 9.6 (6.0–28.3) | 13.4 (4.8–28.2) | 6.9 (0.3–18.6) | 0.695 |
| TNF-α iAUC | 114.6 (6.9–134.4) | 66.9 (5.5–166.9) | 72.7 (16.9–187.8) | 0.761 |
| IL-10 iAUC | 177.0 (59.6–423.9) | 201.6 (84.8–368.8) | 149.3 (0–414.6) | 0.761 |
| Outcome 1 | Anti-Meal | Neutr-Meal | Pro-Meal | p-Value 2 |
|---|---|---|---|---|
| Glucose iAUC | 24.6 (8.9–124.0) | 40.8 (24.1–67.2) | 23.3 (9.8–43.2) | 0.336 |
| Insulin iAUC | 1693.5 (734.8–3258.6) | 2170.5 (1759.5–4014.0) | 2767.5 (1597.7–2984.6) | 0.336 |
| TAG iAUC | 27.0 (9.0–52.6) a | 63.7 (44.0–74.6) b | 135.6 (50.0–156.1) c | 0.009 |
| TC iAUC | 0 (0–0.64) | 0 (0–0.44) | 0 (0–0.50) | 0.482 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cowan, S.; Gibson, S.; Sinclair, A.J.; Truby, H.; Dordevic, A.L. Meals That Differ in Nutrient Composition and Inflammatory Potential Do Not Differentially Impact Postprandial Circulating Cytokines in Older Adults above a Healthy Weight. Nutrients 2022, 14, 1470. https://doi.org/10.3390/nu14071470
Cowan S, Gibson S, Sinclair AJ, Truby H, Dordevic AL. Meals That Differ in Nutrient Composition and Inflammatory Potential Do Not Differentially Impact Postprandial Circulating Cytokines in Older Adults above a Healthy Weight. Nutrients. 2022; 14(7):1470. https://doi.org/10.3390/nu14071470
Chicago/Turabian StyleCowan, Stephanie, Simone Gibson, Andrew J. Sinclair, Helen Truby, and Aimee L. Dordevic. 2022. "Meals That Differ in Nutrient Composition and Inflammatory Potential Do Not Differentially Impact Postprandial Circulating Cytokines in Older Adults above a Healthy Weight" Nutrients 14, no. 7: 1470. https://doi.org/10.3390/nu14071470
APA StyleCowan, S., Gibson, S., Sinclair, A. J., Truby, H., & Dordevic, A. L. (2022). Meals That Differ in Nutrient Composition and Inflammatory Potential Do Not Differentially Impact Postprandial Circulating Cytokines in Older Adults above a Healthy Weight. Nutrients, 14(7), 1470. https://doi.org/10.3390/nu14071470