1. Introduction
Obesity is a global health issue characterized by greater energy intake than energy expenditure, with excess energy stored as triglycerides (TGs) in white adipose tissues (WATs), resulting in fat accumulation. In addition, obesity-related complications include dyslipidemia, hypertension, non-alcoholic fatty liver, type 2 diabetes, heart disease, and some types of cancers [
1,
2], which increase the burden of social medical care. Therefore, it is important to find treatments and interventions for obesity.
Menopause is the spontaneous cessation of menstrual cycles for one year and is caused by the loss of ovarian function, usually around 45–55 years old [
3]. Postmenopausal women have a higher incidence of body fat accumulation and obesity than premenopausal women. Decreased estrogen secretion in postmenopausal women induces changes in the body fat distribution, especially an increased visceral fat accumulation, leading to abdominal obesity [
4,
5,
6,
7]. Abdominal fat has a high rate of lipolysis, which triggers excess free fatty acid (FFA) production from excessive visceral fat lipolysis, promotes insulin resistance (IR), and leads to metabolic diseases [
8]. An ovariectomy (OVX), surgery to remove the ovaries to emulate a state of low estrogen, is a common animal model of menopause [
9]. An estrogen deficiency induced by an OVX in rats results in moderate excess weight gain [
10,
11] and fat accumulation in the liver or WAT. Previous studies showed that the expressions of genes related to β-oxidation (such as carnitine palmitoyl transferase I (CPT-1)) and fatty acid synthesis (such as acetyl-CoA carboxylase (ACC)) respectively decreased and increased in OVX rodents, resulting in hepatic fat accumulation [
12,
13,
14,
15]. Furthermore, other studies showed that menopausal women had lower lipolysis rates and higher lipoprotein lipase (LPL) activity in adipose tissue, which might contribute to body fat accumulation [
16].
Irisin is a myokine, which is produced by the cleavage of fibronectin type III domain-containing 5 (FNDC5) in muscles. Irisin can stimulate the browning of WATs by increasing the uncoupling protein-1 (UCP-1) inducing non-shivering thermogenesis, thereby, increasing the energy expenditure [
17,
18]. A previous study showed that OVX rats exhibited lower irisin levels and higher body weight (BW) gain, which recovered to a level comparable to a control group with continuous irisin treatment, thereby, reducing weight gain. [
19].
Melatonin (Mel;
N-acetyl-5-methoxytryptamine) is mostly provided by the pineal gland [
20]. Mel is responsible for regulating circadian rhythms [
21,
22,
23], reducing inflammatory responses [
24,
25,
26] and oxidative stress [
26,
27,
28], and as an anti-obesity agent [
29]. Several studies showed that Mel supplementation suppressed BW gain in obese rodents without affecting food intake [
30,
31,
32,
33]. Another study suggested that Mel could regulate lipid metabolism, as male rats treated with 10 mg/kg BW Mel for 60 days had significantly reduced the lipogenesis-related gene expression, and promoted the lipolysis-related gene expression, and significantly reduced the hepatic lipid and plasma lipid indices [
34]. In addition, some physiological or pathophysiological states, such as aging [
35], shift work [
36], and nocturnal light pollution [
37,
38], can lead to lower circulating Mel levels, which are accompanied by an energy metabolism imbalance, IR, obesity, and metabolic syndrome. A clinical study showed that postmenopausal women had significantly lower Mel levels than premenopausal women [
39,
40]. However, the effects and mechanisms of Mel on improving obesity in OVX rats are still unclear.
As a result, this study aimed to explore the effects and underlying mechanisms of Mel on lipid metabolism, body fat accumulation, and obesity in OVX rats.
2. Materials and Methods
2.1. Animals
Twenty-four female Sprague–Darley rats that underwent an OVX (9 weeks of age) were purchased from BioLasco (Taipei, Taiwan). The experimental rats were housed in an animal room with a 12 h light/dark cycle (lights on at 07:00). Additionally, the temperature and relative humidity were controlled at 22 ± 2 °C, 65% ± 5%. The experiment started after 4 weeks of the surgical recovery period. During the surgical recovery period, water and the standard laboratory diet (Rodent Laboratory Chow 5001; PMI Nutrition International, St. Louis, MO, USA) were allowed ad libitum. This experiment was approved by the Institutional Animal Care and Use Committee of Taipei Medical University (IACUC no.: LAC-2020-0355).
2.2. Experimental Design
All OVX rats were randomly assigned to different experimental groups (n = 6 in each group) after a surgical recovery period, including a control (C, received the 8% ethanol vehicle) group, low-dose (L, 10 mg Mel/kg BW/day), medium-dose (M, 20 mg Mel/kg BW/day), and high-dose (H, 50 mg Mel/kg BW/day) groups. Mel (Sigma-Aldrich, St Louis, MO, USA) was dissolved in an 8% ethanol (w/v) aqueous solution and given by gavage (at 18:00–19:00). During the 8 weeks of the Mel intervention, BW, food intake, and drinking water were recorded weekly, and the feed efficiency ratio (FER, %; food intake/BW gain) was calculated. Rats were sacrificed with a mixed solution of zoletil and rompun (1 mL/kg BW, intraperitoneal injection) after 8 weeks of the experiment. Serum was collected and centrifuged at 3500× g at 4 °C for 15 min. The uterus, liver, gonadal WATs, perirenal WATs, quadricep femur muscle tissues, and gastrocnemius muscle tissues were dissected and weighed. All samples were stored at −80 °C for further analysis.
2.3. Serum Measurements
The serum estradiol and follicular-stimulating hormone (FSH) level were analyzed by an enzyme-linked immunosorbent assay (ELISA) Kit (Wuhan Fine Biotech, Wuhan, China). The fasting glucose level was tested by a glucose monitor (Abbott Diabetes Care, Oyl, Witney, UK); the serum insulin level was analyzed using an ELISA kit (Mercodia, Uppsala, Sweden); and the homeostasis model assessment (HOMA)-IR index was calculated by applying the equation: HOMA-IR = (fasting blood glucose (mg/dL) × insulin (mIU/L))/405. The serum leptin concentration was measured using an ELISA Kit (BioVender, Brno, Czech Republic); the serum adiponectin concentration was analyzed using an ELISA Kit (AssayPro, St. Charles, MI, USA); and the serum irisin level was analyzed with an ELISA Kit (Bio Vender, Brno, Czech Republic).
2.4. Hepatic Lipid Measurements
Total hepatic lipid was extracted via the Folch methods [
41]; the hepatic TC and TG contents (milligrams per gram (mg/g) of liver tissue) were analyzed by a colorimetric assay kit (Randox Laboratories).
2.5. Real-Time Reverse-Transcription Polymerase Chain Reaction (RT-PCR)
Total RNA from the liver, gonadal adipose tissues, and quadricep femur muscle tissues were extracted with a Trizol reagent (Life Technologies, Carlsbad, CA, USA). The messenger (m)RNA levels of ACC, FAS, CPT-1, acyl CoA oxidase (ACO), peroxisome proliferator-activated receptor alpha (PPARα), 5′AMP-activated protein kinase (AMPK), LPL, hormone-sensitive lipase (HSL), PPARγ, fibronectin type III domain-containing protein 5 (FNDC5), and peroxisome proliferator-activated receptor-gamma coactivator (PGC-1α) were quantified by a real-time qPCR following previous methods [
42]. Data were normalized using β-actin as an internal control, and the relative gene expression was calculated using the 2
−ΔΔCt method. The primer sequences are shown in
Table 1.
2.6. Histopathological Examination
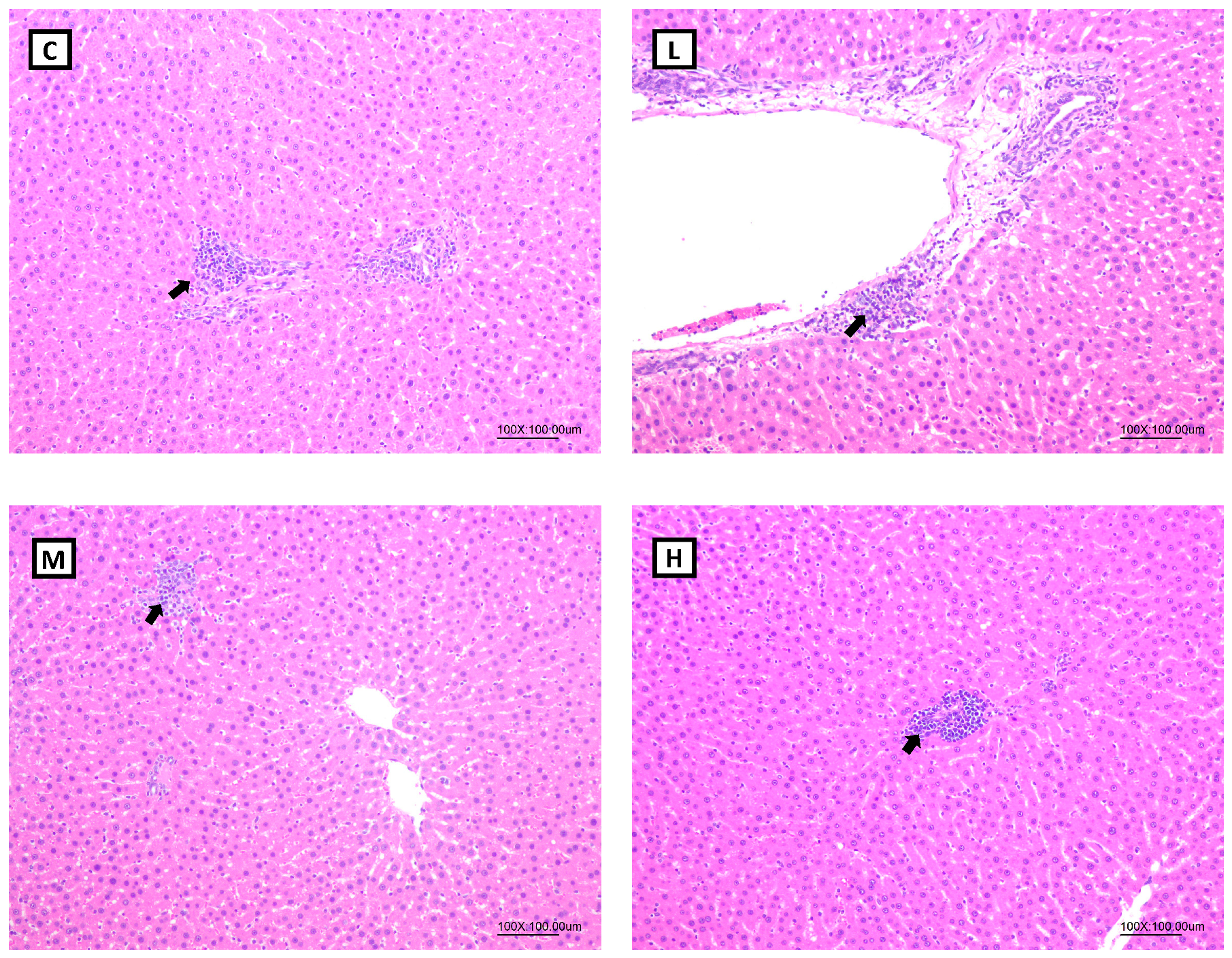
Liver and gonadal WATs were fixed in 10% neutral buffered formaldehyde. Samples were trimmed, embedded in paraffin, sectioned, stained with hematoxylin and eosin (H&E), and examined microscopically by a veterinarian pathologist who was blinded to the nature of the treatment received by the respective groups. Histopathological evaluations were graded as described by Shackelford et al. and Kleiner et al. [
43,
44].
2.7. Statistical Analysis
Data were presented as the mean ± standard error of the mean (SEM) and analyzed by GraphPad Prism vers. 8.0 software (GraphPad Software, San Diego, CA, USA). During the Mel intervention period, the differences among all groups were compared using a one-way analysis of variance (ANOVA) followed by the Tukey’s test. Statistical significance was accepted at p < 0.05.
4. Discussion
In this study, we used the same study design of OVX rats as our laboratory’s previous study [
45] to mimic the decrease in the estrogen levels during menopause. Yeh et al. found that compared to sham-operated rats, OVX rats had significantly lower estrogen levels and uterine atrophy, accompanied by increasing BW gain and abdominal fat accumulation, which was consistent with other previous studies [
12,
46,
47]. In addition, Yeh et al. showed that the serum estradiol level in the sham group was 57.52 ± 7.09 (ng/mL), and the weight of the uterus was 0.56 ± 0.08 (g), which were much higher than the OVX rats in this experiment, and there were no significant differences among all the groups. Therefore, it was speculated that the ovariectomy in this experiment was successful. Estrogen plays a crucial role in regulating energy metabolism. Estrogen can reduce appetite by acting directly on anorexigenic and orexigenic neurons in the hypothalamus. In addition, it can also increase thermogenesis in brown adipose tissues (BATs) through the ventromedial hypothalamus-sympathetic nervous system (VMH-SNS)-BAT pathway. As a result, obesity caused by OVX may be attributed to increased food intake and reduced energy expenditure due to an estrogen deficiency [
48].
In the present study, OVX rats treated with Mel supplementation reduced BW gain without affecting food intake, but no dose-dependent effect was found. Similar to previous studies, exogenous Mel supplementation could prevent weight gain in male mice [
34], especially in mice with high-fat diet (HFD)-induced obesity [
49,
50], and no dose effect was found [
51]. A meta-analysis clinical study indicated that compared to a placebo, Mel supplementation reduced BW, and the results were better in studies using doses of ≤8 mg/day. This means that the effect of Mel on BW is not dose dependent. Moreover, the present study found that the abdominal visceral fat mass, such as the gonadal and perirenal fat masses, decreased in OVX rats with Mel supplementation. Similar to previous studies, such as that by Majumdar et al. who found that the intra-abdominal fat mass was decreased in OVX rats with fructose diet-induced obesity and 3 mg/kg BW Mel supplementation in drinking water [
52]. Another clinical study showed that the fat mass decreased in postmenopausal women with 1 or 3 mg/day Mel supplementation for 1 year [
53]. Thus, melatonin supplementation ameliorated body weight and abdominal visceral fat accumulation in OVX rats.
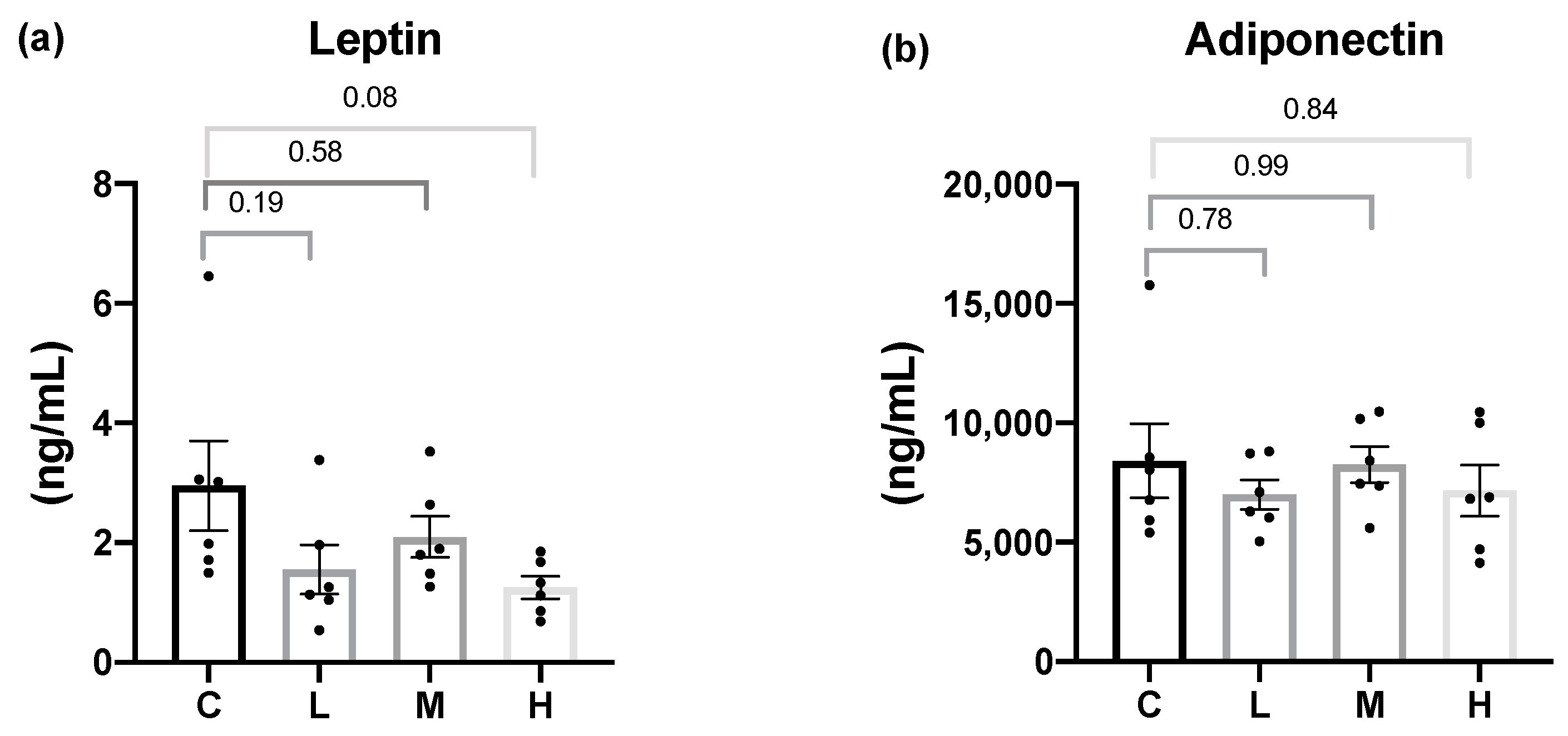
In terms of adipokines, although there were no significant differences in the serum leptin or adiponectin levels among all the groups, compared to the C group, the Mel treatment groups showed a trend of lower serum leptin levels. Previous studies showed that melatonin reduced the serum leptin level [
30,
35], and another study indicated that the reduced leptin level after melatonin treatment was associated with reducing fat mass rather than improving the leptin function [
54]. The result of the leptin level in this study was consistent with the Szewczyk-Golec et al. study, as male rats treated with 20μg/mL Mel for 7 weeks had significantly reduced body weight, but there was no significant difference in the leptin level [
54]. Leptin and melatonin are secreted in a circadian rhythm with acrophases in the nocturnal period. Therefore, melatonin did not affect the leptin level in this study, which might be related to the different time of sample collection, because of the rhythmic secretion of leptin. However, a study by Yeh et al. pointed out that compared with the sham group, there was no significant difference in the leptin level in OVX rats [
45]. Consistent with the study by Babaei et al., eight weeks after ovariectomy, the body weight and visceral fat mass were significantly increased in 11-week-old OVX rats but did not affect the adiponectin level. The possible reason is that OVX can promote adipocytes hyperplasia, not adipocyte hypertrophy; only adipocyte hypertrophy will reduce adiponectin level [
55]. Therefore, we speculate that the OVX animal model in this study did not cause adiponectin secretion disorder. If the secretion is normal, the intervention of melatonin will not affect the adiponectin level.
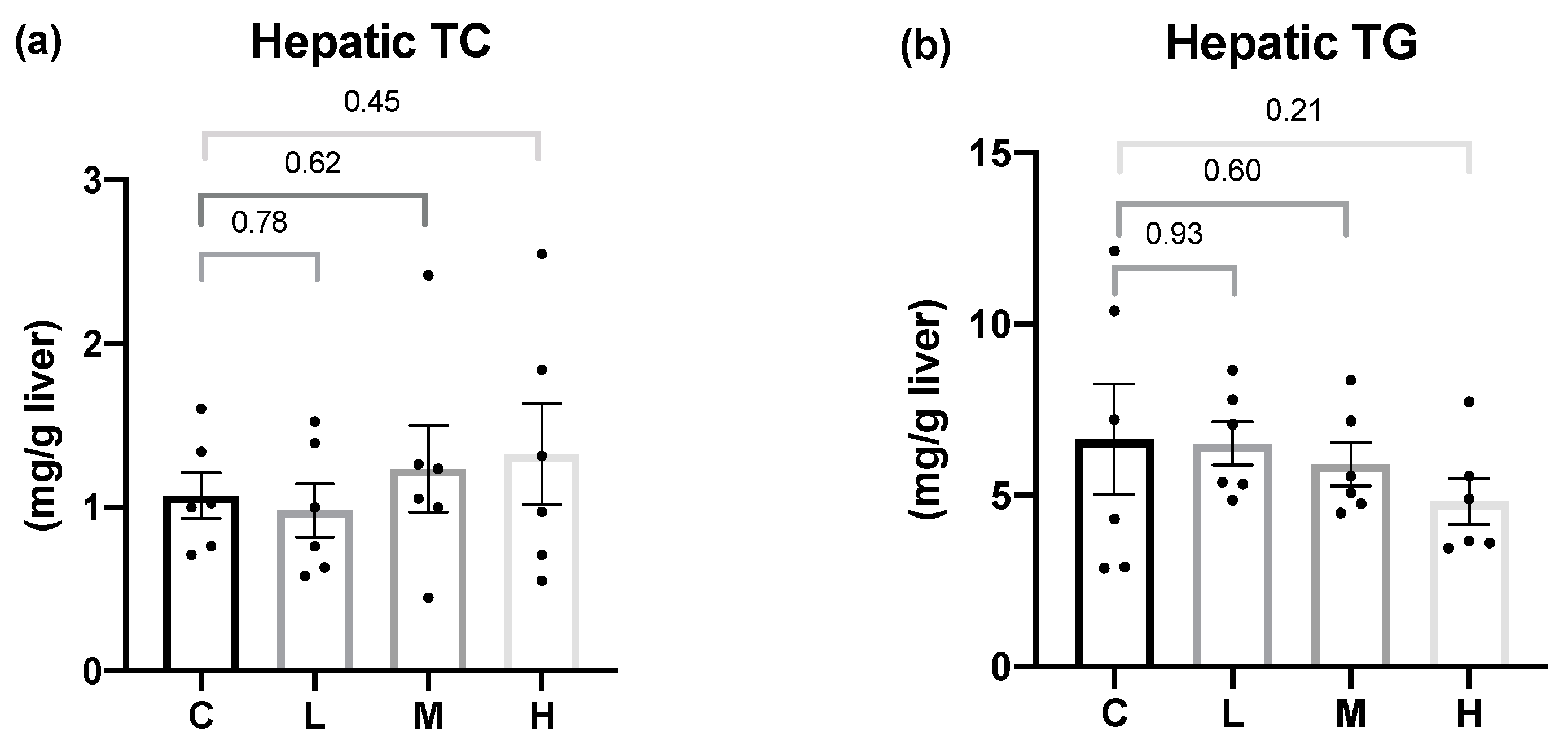
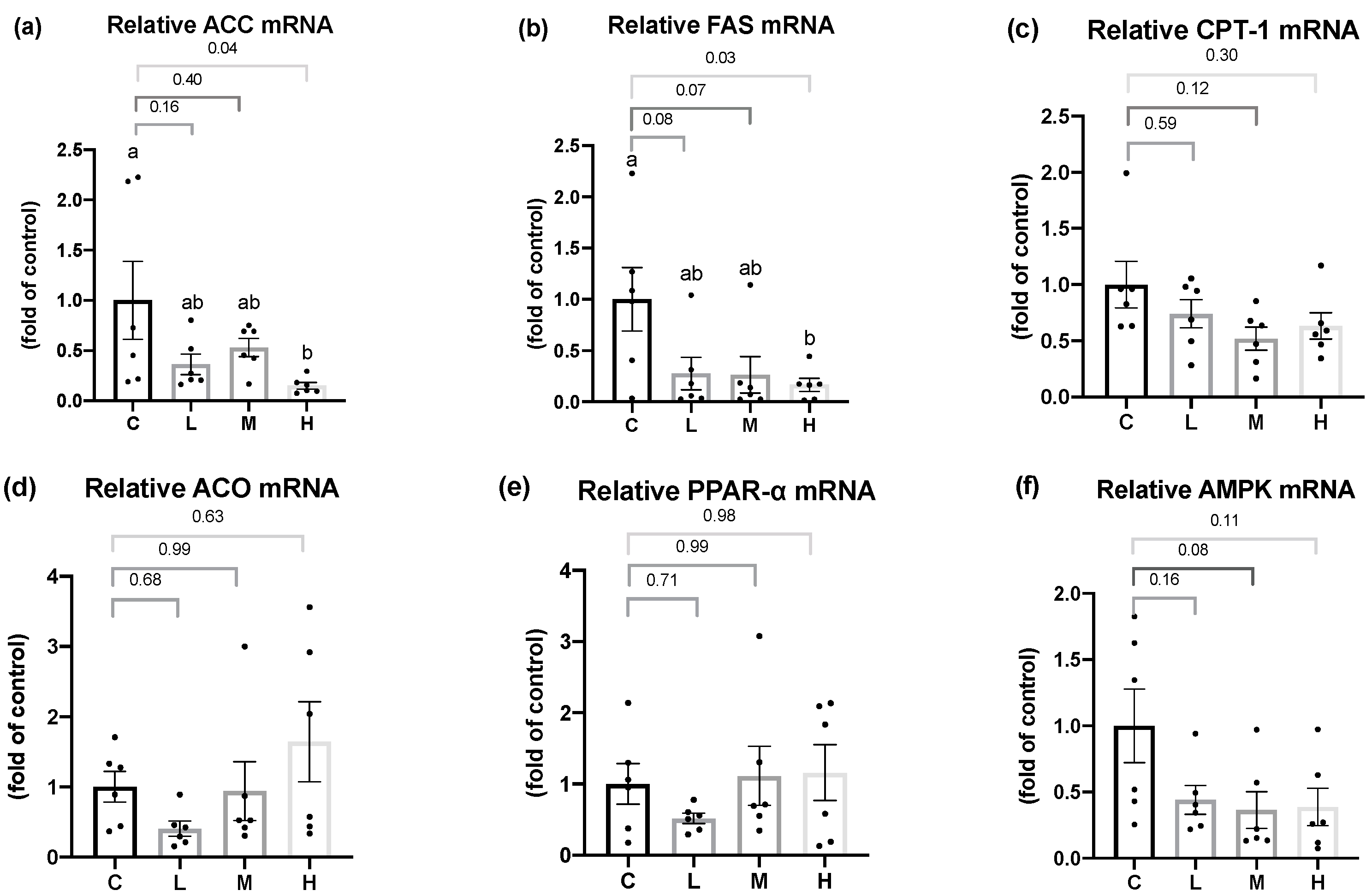
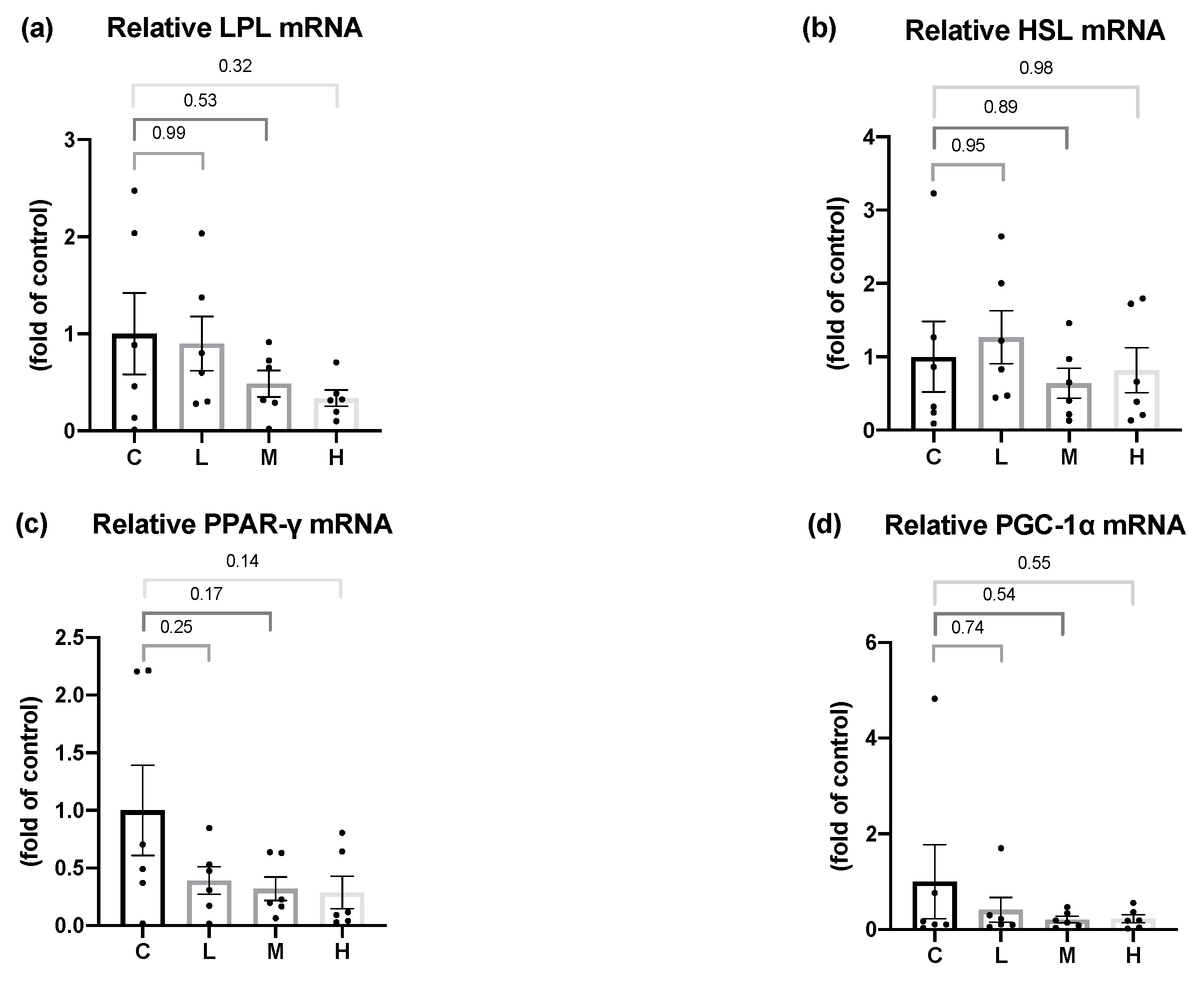
In terms of hepatic lipid metabolism, this study demonstrated that the mRNA levels of the hepatic fatty acid synthesis enzymes, ACC and FAS, were significantly lower in the OVX rats treated with 50 mg/kg BW Mel for 8 weeks, but no difference was found in the hepatic fatty acid oxidation enzymes. Past studies found that Mel could regulate the hepatic lipid metabolism. Ou et al. administered 10, 20, and 50 mg/kg BW of Mel to hamsters with HFD-induced hyperlipidemia, and found that Mel treatment decreased the activities of the ACC and FAS lipogenic enzymes, and increased the mRNA levels of the CPT-1 lipolysis enzymes, while reducing the hepatic TC and TG contents, thereby, decreasing serum TC, TG, and low-density lipoprotein cholesterol (LDL-C) levels [
56]. Liu et al. reported that a daily subcutaneous injection of 10 mg/kg BW Mel activated the liver AMPKα/PPARα/CPT-1 pathway in guinea pigs with glucose and lipid metabolism disorders caused by persistent artificial light exposure and an HFD. Generally, AMPKα can regulate PPARα expression. After its activation by AMPKα, PPARα reduces TG synthesis in the liver by enhancing the CPT-1 expression [
57]. It was speculated that the therapeutic effect of Mel on hepatic fatty acid oxidation was inconsistent with past studies, and possible reasons were as follows: Firstly, compared to an HFD-induced obesity animal model, OVX rats had a milder degree of obesity and liver lipid accumulation, so the effect of Mel regulating the fatty acid oxidation pathway was less significant, resulting in no change in the hepatic lipid contents. This explanation can be verified from the hepatic histopathology results, as steatotic and necrotic lesions were not observed in any of the groups. Secondly, it might be related to the method of the Mel intervention. Compared to a subcutaneous injection of Mel, the absorption rate of oral Mel is lower, and the bioavailability is only 9%~33% [
58], thus, the designed dosage in this study might not have been sufficient to affect the hepatic lipid metabolism pathways. On the other hand, Heo et al. reported that the administration of 50 or 100 mg/kg BW of Mel for 10 weeks to rats with HFD-induced obesity resulted in hepatic histopathological images showing that 100 mg/kg BW Mel significantly reduced the hepatic steatosis scores, but 50 mg/kg BW Mel showed a downward trend, and the differences did not reach statistical significance [
59]. As a result, by increasing the dosage of Mel or the intervention time, one may be able to observe more-significant effects of Mel on hepatic lipid metabolism.
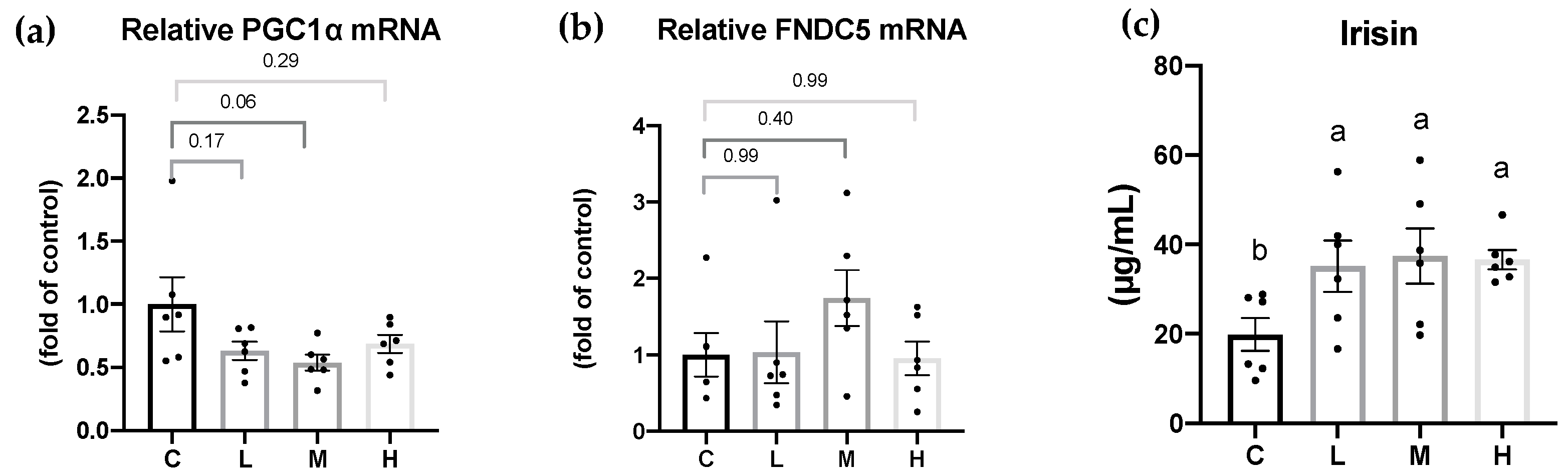
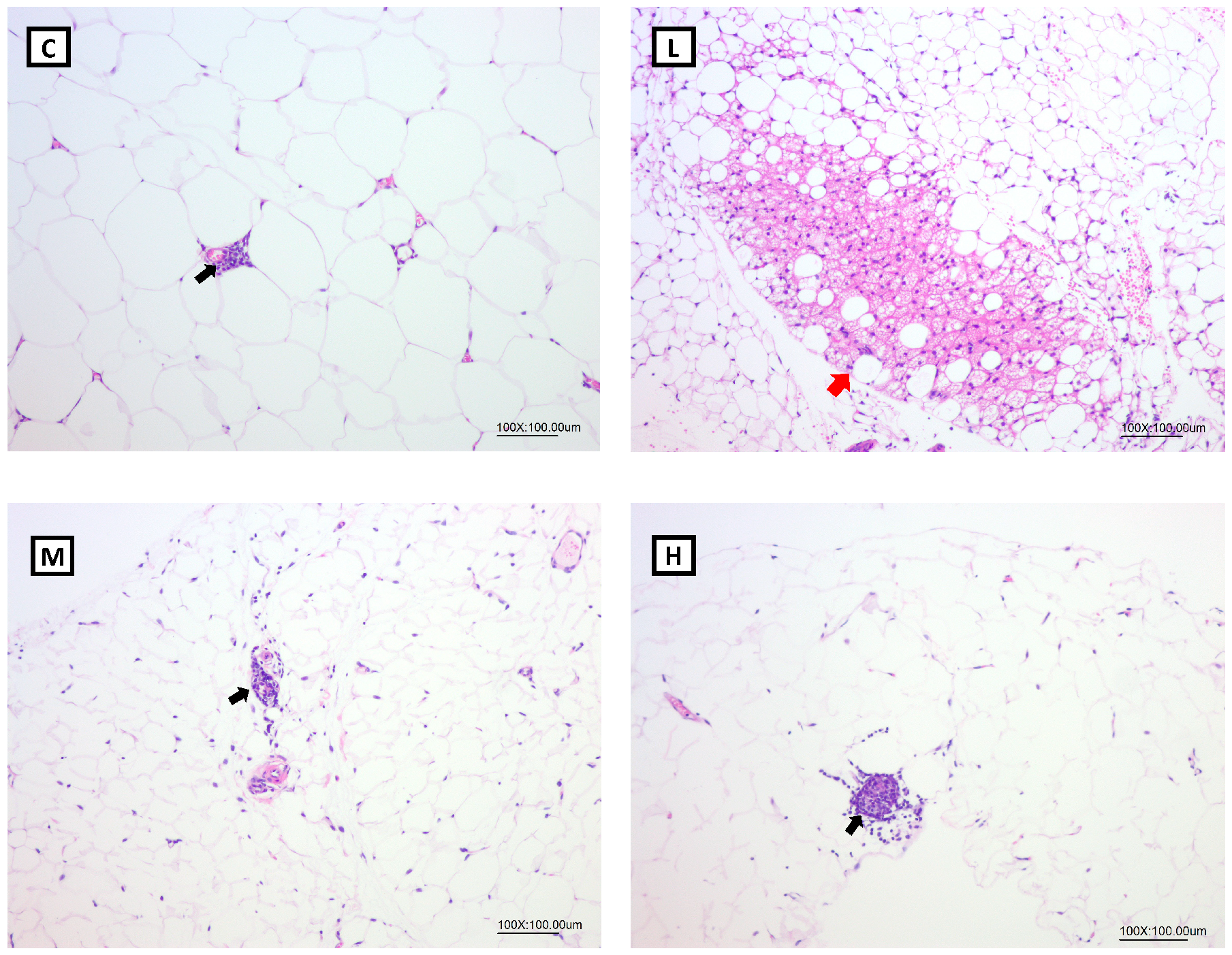
Previous studies suggested that Mel had an anti-obesity effect by the browning of WATs, thereby increasing energy expenditure [
51,
60]. Obese Zücker diabetic fatty (ZDF) rats were supplemented with 10 mg/kg Mel in drinking water for 6 weeks, and the number of brite/beige adipocytes in inguinal WATs increased, and PGC-1α and UCP-1 levels were significantly higher and promoted non-shivering thermogenesis [
60]. In another study, Tung et al. treated obesity rats with 10, 20, and 50 mg/kg BW Mel for 8 weeks, and found that Mel reduced BW gain and increased the circulating irisin concentrations; an increased number of brite/beige adipocytes was observed in the inguinal WATs [
51]. In the present study, Mel supplementation elevated the serum irisin levels. In addition, 10 and 50 mg/kg BW Mel induced brite/beige adipocytes in the gonadal WATs, but no brite/beige adipocytes were observed in the M group. The possible reasons were as follows: The white adipose tissues for morphological analysis were randomly collected at the time of sacrifice. Additionally, the number of samples for morphological analysis was small (
n = 3). Thus, the presence of brite/beige adipocytes was not observed in the M group. Moreover, compared to the study by Jiménez-Aranda et al., where they dissected the adipose tissue after undergoing acute cold exposure [
60], this experimental design was more accurate and easier to find browning in the WATs. In future studies, acute cold exposure before sacrifice should be designed and it may be able to observe more-significant effects of melatonin on WAT browning. Previous studies suggested that Mel improved obesity possibly by being implicated in increased physical activity [
30,
33]. Exercise induces the skeletal muscles to cleave FNDC5, and irisin is produced. Once irisin is released into the circulation, it acts on white adipocytes to induce the browning response and subsequently activates non-shivering thermogenesis. Therefore, it was speculated that Mel supplementation could promote irisin secretion, which might be related to physical activity. Although the FNDC5 mRNA levels in muscle tissues only exhibited an upward trend in the melatonin treatment group, the serum irisin concentration was significantly increased in the melatonin treatment group. In addition, the FNDC5 gene expression in the adipose tissue was lower than in the muscle tissue but also correlated with the serum irisin levels [
61]. Thus, the expressions of FNDC5 proteins in the muscle or FNDC5 mRNA levels in the adipose tissue must be measured in future studies to ensure the explicit mechanism of irisin increase. However, some studies showed that Mel did not affect physical activity [
62,
63]. Jiménez-Aranda et al. indicated that Mel treatment did not affect circulating irisin concentrations or physical activity [
60]. Whether Mel increases physical activity is still inconclusive. Although many studies found that Mel or irisin can promote the browning of adipose tissues, few studies had explored the relationship between Mel and irisin. Therefore, elucidating the exact mechanism by which Mel increases FNDC5/Irisin requires further animal and cell studies. However, the results of our study still showed that Mel may be beneficial for regulating the irisin levels and browning of WATs in OVX rats.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}