
Tartrazine Modifies the Activity of DNMT and HDAC Genes—Is This a Link between Cancer and Neurological Disorders?

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Treatment of NMRI Mice
2.3. Sample Collection and mRNA Isolation
2.4. qRT-PCR
2.5. Statistical Analysis
3. Results
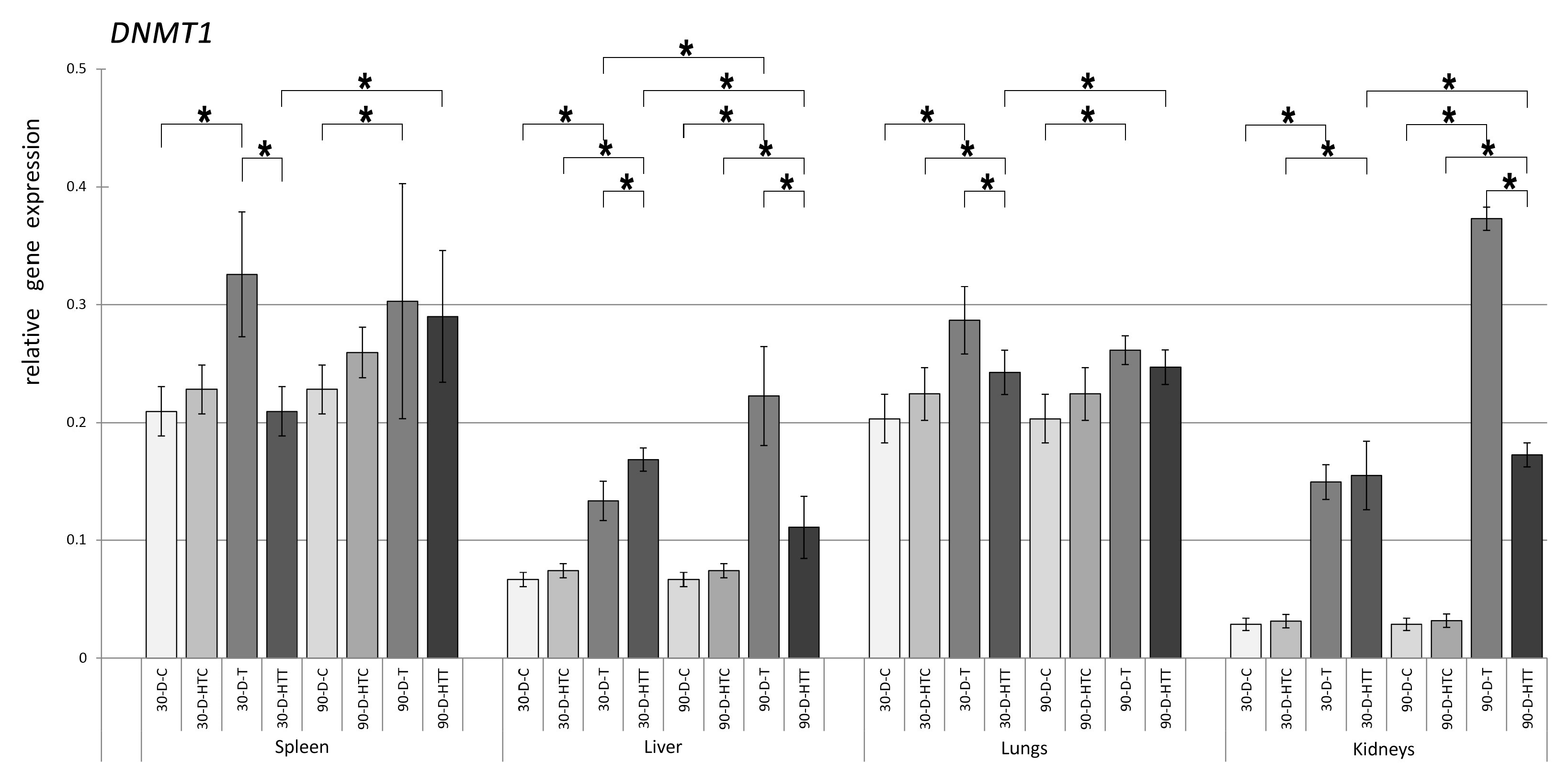
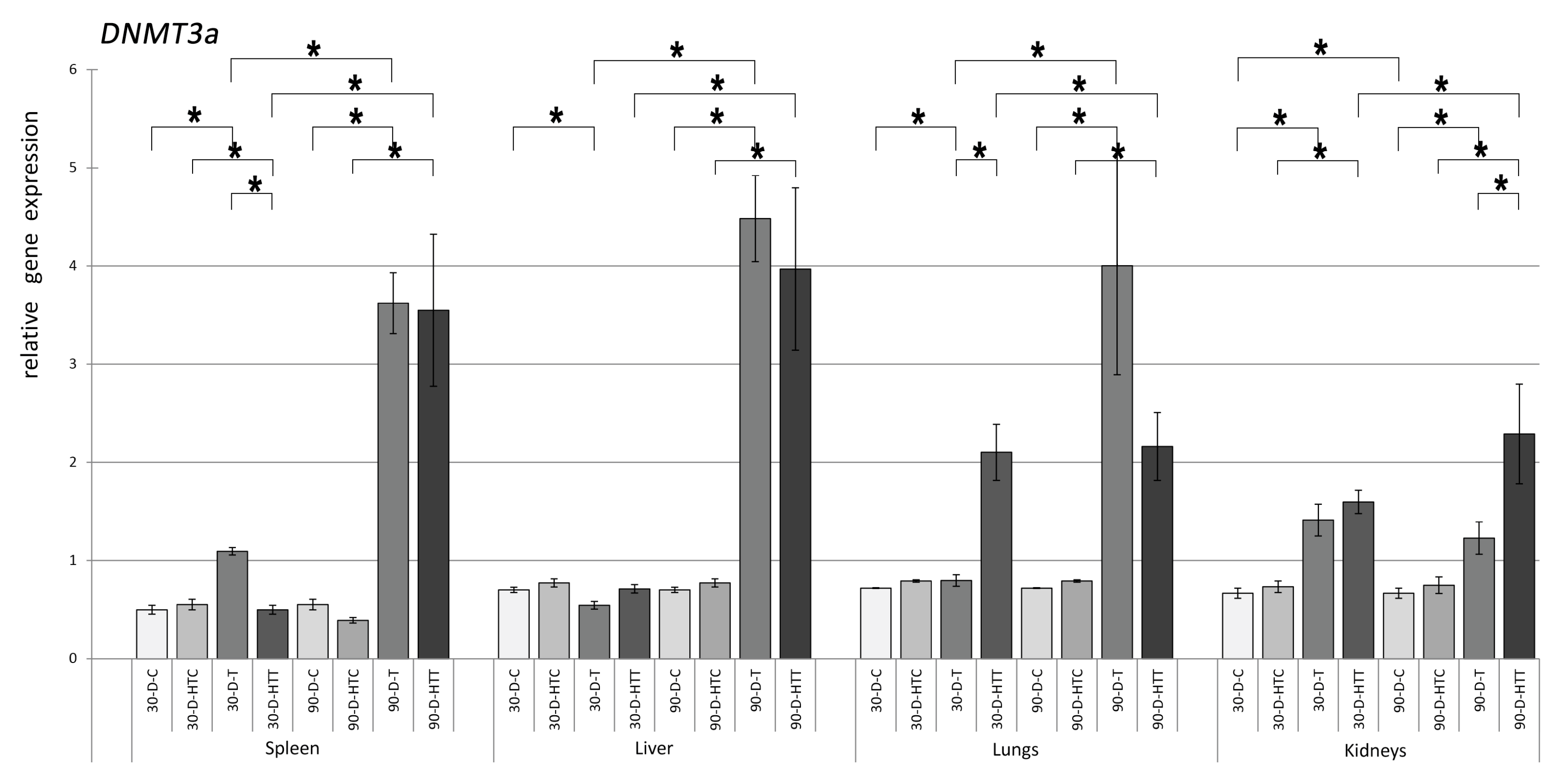
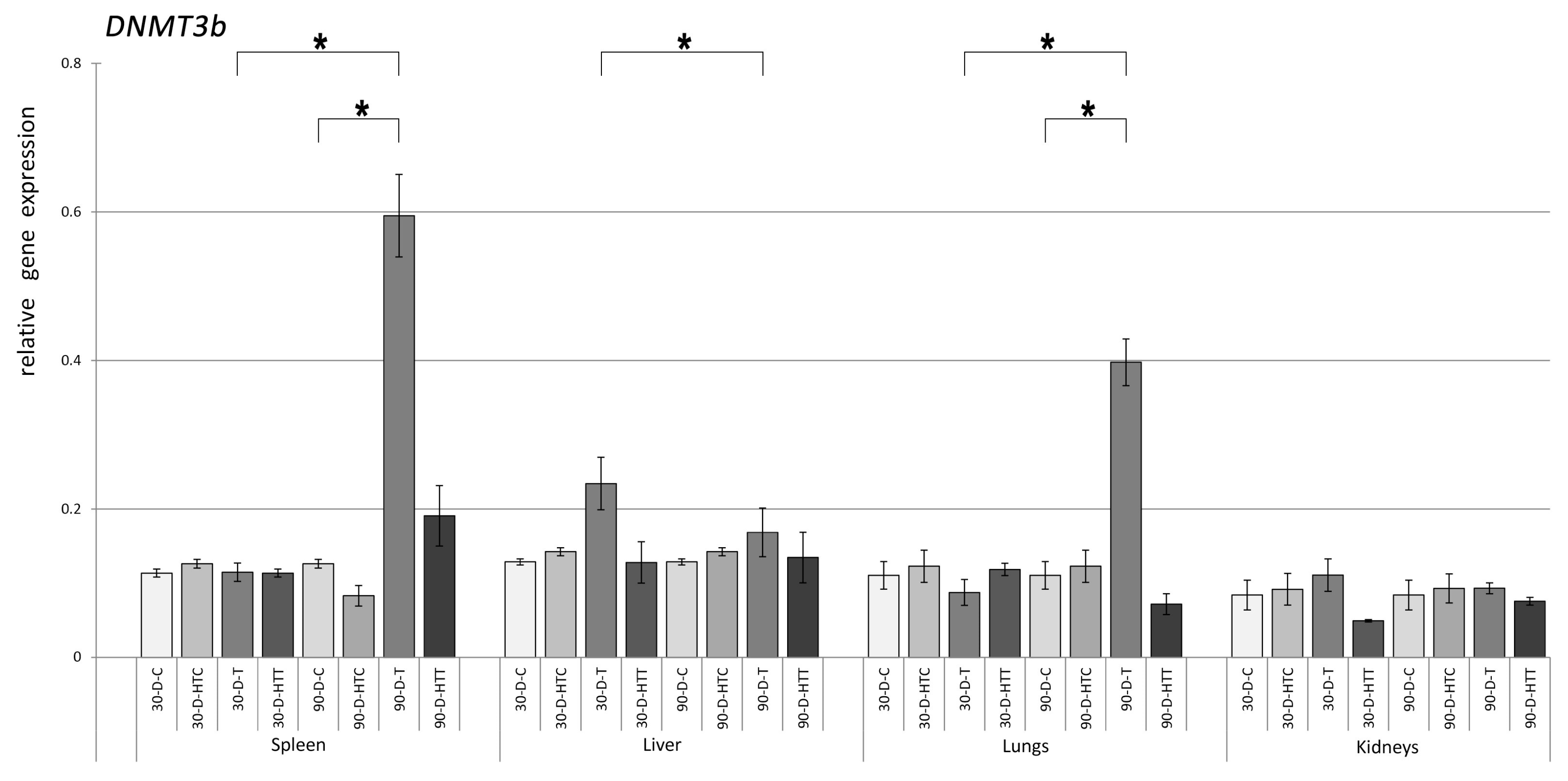
3.1. Expression of DNMT Genes
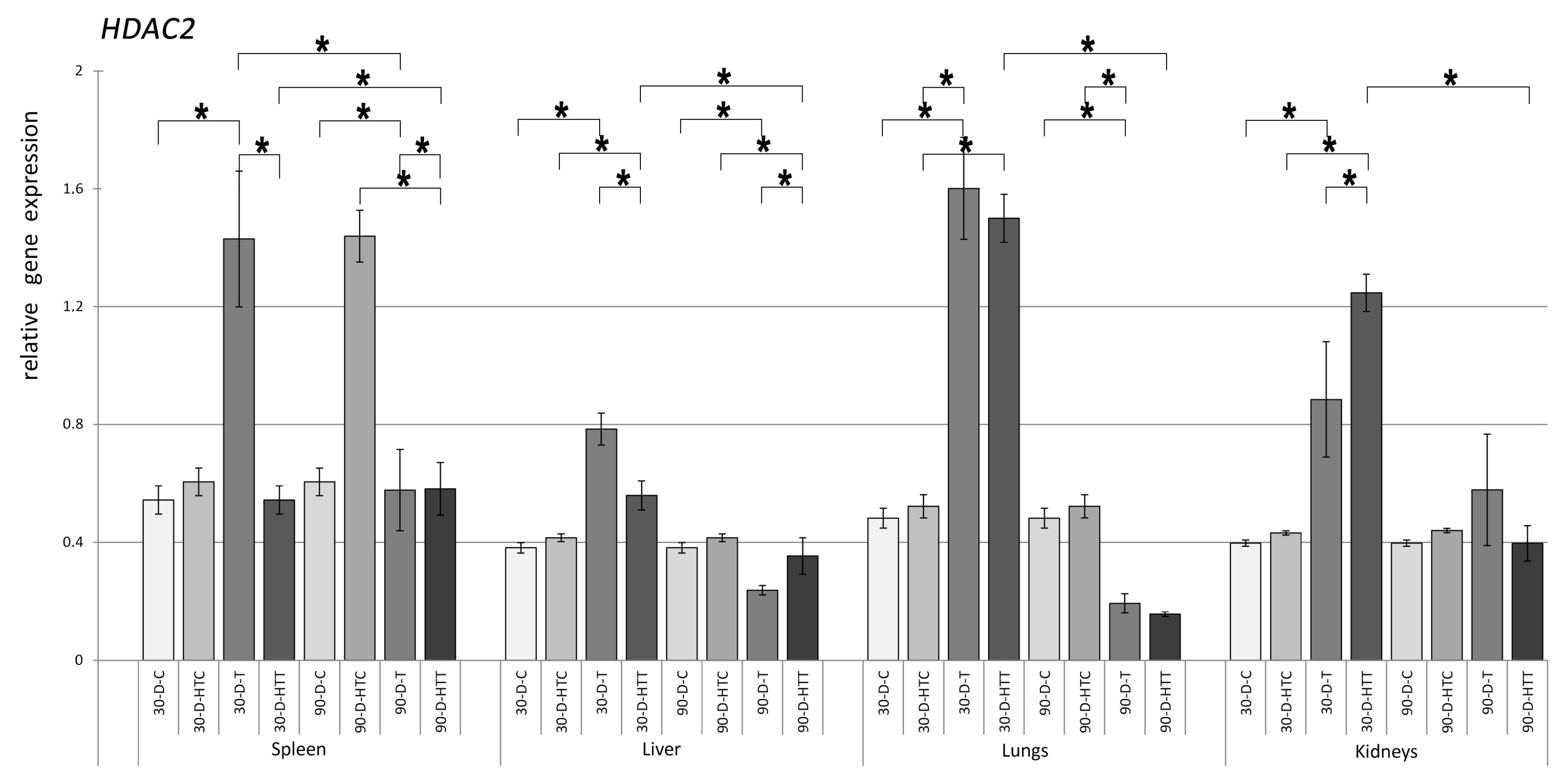
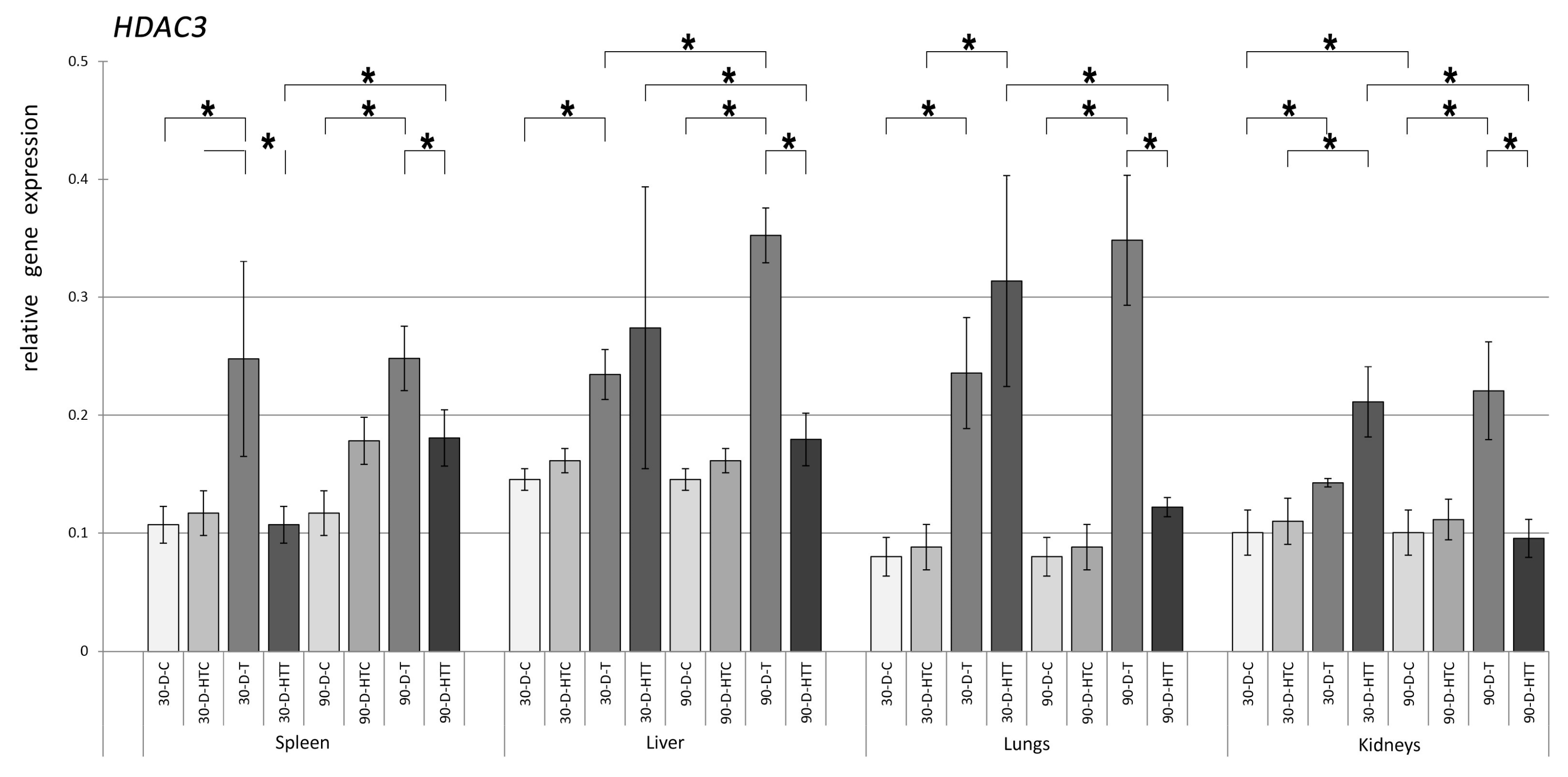
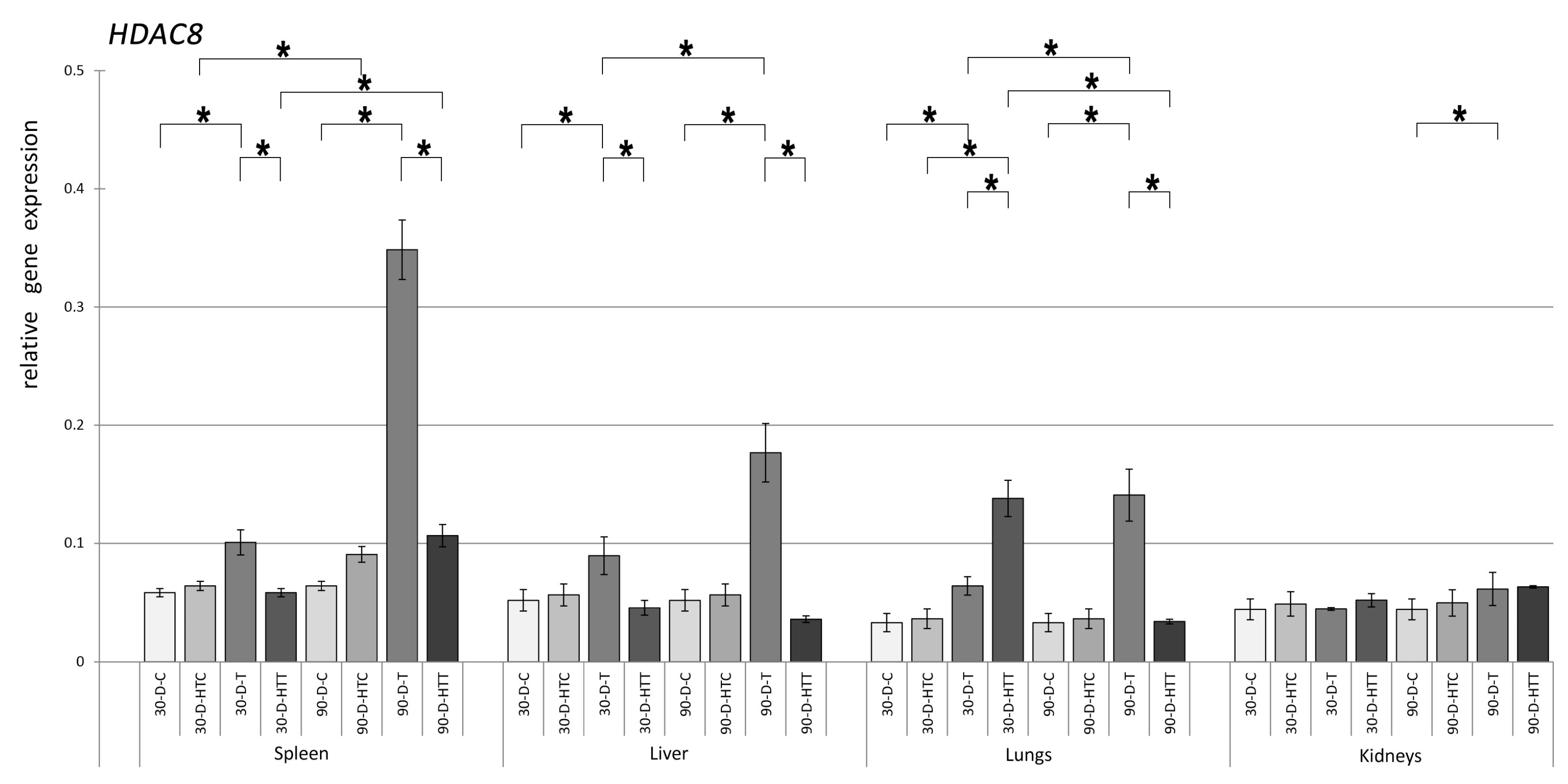
3.2. Expression of HDAC Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mpountoukas, P.; Pantazaki, A.; Kostareli, E.; Christodoulou, P.; Kareli, D.; Poliliou, S.; Mourelatos, C.; Lambropoulou, V.; Lialiaris, T. Cytogenetic Evaluation and DNA Interaction Studies of the Food Colorants Amaranth, Erythrosine and Tartrazine. Food Chem. Toxicol. 2010, 48, 2934–2944. [Google Scholar] [CrossRef]
- Merinas-Amo, R.; Martínez-Jurado, M.; Jurado-Güeto, S.; Alonso-Moraga, Á.; Merinas-Amo, T. Biological Effects of Food Coloring in In Vivo and In Vitro Model Systems. Foods 2019, 8, 176. [Google Scholar] [CrossRef] [Green Version]
- Dixit, S.; Purshottam, S.K.; Khanna, S.K.; Das, M. Usage Pattern of Synthetic Food Colours in Different States of India and Exposure Assessment through Commodities Preferentially Consumed by Children. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2011, 28, 996–1005. [Google Scholar] [CrossRef]
- Husain, A.; Sawaya, W.; Al-Omair, A.; Al-Zenki, S.; Al-Amiri, H.; Ahmed, N.; Al-Sinan, M. Estimates of Dietary Exposure of Children to Artificial Food Colours in Kuwait. Food Addit. Contam. 2006, 23, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Revankar, M.S.; Lele, S.S. Synthetic Dye Decolorization by White Rot Fungus, Ganoderma Sp. WR-1. Bioresour. Technol. 2007, 98, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.T. Azo Dyes and Human Health: A Review. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2016, 34, 233–261. [Google Scholar] [CrossRef]
- Mittal, A.; Kurup, L.; Mittal, J. Freundlich and Langmuir Adsorption Isotherms and Kinetics for the Removal of Tartrazine from Aqueous Solutions Using Hen Feathers. J. Hazard. Mater. 2007, 146, 243–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joint FAO/WHO Expert Committee on Food Additives. 44th Meeting, Rome, Ialy, 1995; International Program on Chemical Safety. In Toxicological Evaluation of Certain Food Additives and Contaminants in Food; Joint FAO/WHO Expert Committee on Food Additives: Geneva, Switzerland, 1996; p. 465. [Google Scholar]
- Specifications for the Identity and Purity of Food Additives and Their Toxicological Evaluation: Food Colours and Some Antimicrobials and Antioxidants, Eighth Report of the Joint FAO/WHO Expert Committee on Food Additives, Geneva, Switzerland, 8–17 December 1964. Available online: https://apps.who.int/iris/handle/10665/40627 (accessed on 10 June 2023).
- Joint FAO World Health Organization, & WHO Expert Committee on Food Additives. Evaluation of Certain Food Additives: Eighty-Second Report of the Joint FAO. World Health Organization. 2016. Available online: http://apps.who.int/iris/bitstream/handle/10665/250277/9789241210003-eng.pdf?sequence=1#page=75%22%3E (accessed on 10 June 2023).
- McCann, D.; Barrett, A.; Cooper, A.; Crumpler, D.; Dalen, L.; Grimshaw, K.; Kitchin, E.; Lok, K.; Porteous, L.; Prince, E.; et al. Food Additives and Hyperactive Behaviour in 3-Year-Old and 8/9-Year-Old Children in the Community: A Randomised, Double-Blinded, Placebo-Controlled Trial. Lancet 2007, 370, 1560–1567. [Google Scholar] [CrossRef]
- Amin, K.A.; Abdel Hameid, H.; Abd Elsttar, A.H. Effect of Food Azo Dyes Tartrazine and Carmoisine on Biochemical Parameters Related to Renal, Hepatic Function and Oxidative Stress Biomarkers in Young Male Rats. Food Chem. Toxicol. 2010, 48, 2994–2999. [Google Scholar] [CrossRef]
- Maekawa, A.; Matsuoka, C.; Onodera, H.; Tanigawa, H.; Furuta, K.; Kanno, J.; Jang, J.J.; Hayashi, Y.; Ogiu, T. Lack of Carcinogenicity of Tartrazine (FD & C Yellow No. 5) in the F344 Rat. Food Chem. Toxicol. 1987, 25, 891–896. [Google Scholar] [CrossRef]
- Chung, K.T.; Stevens, S.E.; Cerniglia, C.E. The Reduction of Azo Dyes by the Intestinal Microflora. Crit. Rev. Microbiol. 1992, 18, 175–190. [Google Scholar] [CrossRef]
- Moutinho, I.L.D.; Bertges, L.C.; Assis, R.V.C. Prolonged Use of the Food Dye Tartrazine (FD&C Yellow N° 5) and Its Effects on the Gastric Mucosa of Wistar Rats. Braz. J. Biol. 2007, 67, 141–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ameur, F.Z.; Mehedi, N.; Kheroua, O.; Saïdi, D.; Salido, G.M.; Gonzalez, A. Sulfanilic Acid Increases Intracellular Free-Calcium Concentration, Induces Reactive Oxygen Species Production and Impairs Trypsin Secretion in Pancreatic AR42J Cells. Food Chem. Toxicol. 2018, 120, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Mekkawy, H.A.; Ali, M.O.; El-Zawahry, A.M. Toxic Effect of Synthetic and Natural Food Dyes on Renal and Hepatic Functions in Rats. Toxicol. Lett. 1998, 95, 155. [Google Scholar] [CrossRef]
- Sarikaya, R.; Selvi, M.; Erkoç, F. Evaluation of Potential Genotoxicity of Five Food Dyes Using the Somatic Mutation and Recombination Test. Chemosphere 2012, 88, 974–979. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya, R.; Çakir, Ş. Genotoxicity Testing of Four Food Preservatives and Their Combinations in the Drosophila Wing Spot Test. Environ. Toxicol. Pharmacol. 2005, 20, 424–430. [Google Scholar] [CrossRef]
- Poul, M.; Jarry, G.; Elhkim, M.O.; Poul, J.M. Lack of Genotoxic Effect of Food Dyes Amaranth, Sunset Yellow and Tartrazine and Their Metabolites in the Gut Micronucleus Assay in Mice. Food Chem. Toxicol. 2009, 47, 443–448. [Google Scholar] [CrossRef]
- Hassan, G.M. Effects of Some Synthetic Coloring Additives on DNA Damage and Chromosomal Aberrations of Rats. Arab. J. Biotechnol. 2010, 13, 13–24. [Google Scholar]
- Yurchenko, V.V.; Ingel, F.I.; Akhaltseva, L.V.; Konyashkina, M.A.; Yurtseva, N.A.; Nikitina, T.A.; Krivtsova, E.K. Genotoxic Safety of Synthetic Food Colours. Review. Ecol. Genet. 2021, 19, 323–341. [Google Scholar] [CrossRef]
- An, Y.; Jiang, L.; Cao, J.; Geng, C.; Zhong, L. Sudan I Induces Genotoxic Effects and Oxidative DNA Damage in HepG2 Cells. Mutat. Res. 2007, 627, 164–170. [Google Scholar] [CrossRef]
- Atlı Şekeroğlu, Z.; Güneş, B.; Kontaş Yedier, S.; Şekeroğlu, V.; Aydın, B. Effects of Tartrazine on Proliferation and Genetic Damage in Human Lymphocytes. Toxicol. Mech. Method. 2017, 27, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Albasher, G.; Maashi, N.; Alfarraj, S.; Almeer, R.; Albrahim, T.; Alotibi, F.; Bin-Jumah, M.; Mahmoud, A.M. Perinatal Exposure to Tartrazine Triggers Oxidative Stress and Neurobehavioral Alterations in Mice Offspring. Antioxidants 2020, 9, 53. [Google Scholar] [CrossRef] [Green Version]
- Essawy, A.E.; Mohamed, A.I.; Ali, R.G.; Ali, A.M.; Abdou, H.M. Analysis of Melatonin-Modulating Effects Against Tartrazine-Induced Neurotoxicity in Male Rats: Biochemical, Pathological and Immunohistochemical Markers. Neurochem. Res. 2023, 48, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.W.; Turnbull, D.M. Mitochondrial DNA Mutations in Human Disease. Nat. Rev. Genet. 2005, 6, 389–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteller, M. Epigenetics in Cancer. N. Engl. J. Med. 2008, 358, 1148–1159. [Google Scholar] [CrossRef] [PubMed]
- Poh, W.J.; Wee, C.P.P.; Gao, Z. DNA Methyltransferase Activity Assays: Advances and Challenges. Theranostics 2016, 6, 369–391. [Google Scholar] [CrossRef]
- de Ruijter, A.J.M.; van Gennip, A.H.; Caron, H.N.; Kemp, S.; van Kuilenburg, A.B.P. Histone Deacetylases (HDACs): Characterization of the Classical HDAC Family. Biochem. J. 2003, 370, 737. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Kim, J.S. A Short Guide to Histone Deacetylases Including Recent Progress on Class II Enzymes. Exp. Mol. Med. 2020, 52, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Glozak, M.A.; Seto, E. Histone Deacetylases and Cancer. Oncogene 2007, 26, 5420–5432. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, S.; Bharat, R.B.; Lee, K.H.; Cho, H. Some of the Food Color Additives Are Potent Inhibitors of Human Protein Tyrosine Phosphatases. Bull. Korean Chem. Soc. 2006, 27, 1567–1571. [Google Scholar] [CrossRef] [Green Version]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose Translation from Animal to Human Studies Revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, E.A.; Chipman, J.K.; Forsythe, S.J. Evidence for Direct-Acting Oxidative Genotoxicity by Reduction Products of Azo Dyes. Environ. Health Perspect. 1994, 102 (Suppl. S6), 119–122. [Google Scholar] [CrossRef] [Green Version]
- Cerniglia, C.E.; Zhuo, Z.; Manning, B.W.; Federle, T.W.; Heflich, R.H. Mutagenic Activation of the Benzidine-Based Dye Direct Black 38 by Human Intestinal Microflora. Mutat. Res. Lett. 1986, 175, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Bansal, A.K.; Bansal, M.; Soni, G.; Bhatnagar, D. Modulation of N-Nitrosodiethylamine (NDEA) Induced Oxidative Stress by Vitamin E in Rat Erythrocytes. Hum. Exp. Toxicol. 2016, 24, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Abo-EL-Sooud, K.; Hashem, M.M.; Badr, Y.A.; Eleiwa, M.M.E.; Gab-Allaha, A.Q.; Abd-Elhakim, Y.M.; Bahy-EL-Dien, A. Assessment of Hepato-Renal Damage and Genotoxicity Induced by Long-Term Exposure to Five Permitted Food Additives in Rats. Environ. Sci. Pollut. Res. Int. 2018, 25, 26341–26350. [Google Scholar] [CrossRef]
- Islam, B.U.; Habib, S.; Ahmad, P.; Allarakha, S.; Moinuddin; Ali, A. Pathophysiological role of peroxynitrite induced DNA damage in human diseases: A special focus on poly (ADP-ribose) polymerase (PARP). Ind. J. Clin. Biochem. 2015, 30, 368–385. [Google Scholar] [CrossRef] [Green Version]
- Himri, I.; Souna, F.; Belmekki, F.; Aziz, M.; Bnouham, M.; Zoheir, J.; Berkia, Z.; Mekhfi, H.; Saalaoui, E. A 90-day oral toxicity study of tartrazine, a synthetic food dye, in Wistar rats. Int. J. Pharm. Pharm. Sci. 2011, 3, 159–169. [Google Scholar]
- Khayyat, L.; Essawy, A.; Sorour, J.; Soffar, A. Tartrazine Induces Structural and Functional Aberrations and Genotoxic Effects in Vivo. PeerJ 2017, 5, e3041. [Google Scholar] [CrossRef] [Green Version]
- Demirkol, O.; Zhang, X.; Ercal, N. Oxidative Effects of Tartrazine (CAS No. 1934-21-0) and New Coccin (CAS No. 2611-82-7) Azo Dyes on CHO Cells. J. Verbrauch. Lebensm. 2012, 7, 229–236. [Google Scholar] [CrossRef]
- Kietzmann, T.; Petry, A.; Shvetsova, A.; Gerhold, J.M.; Görlach, A. The epigenetic landscape related to reactive oxygen species formation in the cardiovascular system. Brit. J. Pharmacol. 2017, 174, 1533–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakoor, S.; Ismail, A.; Redzwan Sabran, M.; Mohtarrudin, N. Effect of Food Colorants Supplementation on Reactive Oxygen Species, Antioxidant Vitamins Level and DNA Damage (Kesan Tambahan Pewarna Makanan Pada Spesies Oksigen Reaktif, Tahap Antioksidan Vitamin Dan Kerosakan DNA). Sains. Malays. 2021, 50, 1343–1356. [Google Scholar] [CrossRef]
- Prasad, R.; Katiyar, S.K. Prostaglandin E2 Promotes UV Radiation-Induced Immune Suppression through DNA Hypermethylation. Neoplasia 2013, 15, 795–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, R.B.; Resende, W.R.; Dal-Pont, G.C.; Gava, F.F.; Tye, S.J.; Quevedo, J.; Valvassori, S.S. HDAC Inhibitors Reverse Mania-like Behavior and Modulate Epigenetic Regulatory Enzymes in an Animal Model of Mania Induced by Ouabain. Pharmacol. Biochem. Behav. 2020, 193, 172917. [Google Scholar] [CrossRef]
- Dash, P.K.; Orsi, S.A.; Zhang, M.; Grill, R.J.; Pati, S.; Zhao, J.; Moore, A.N. Valproate Administered after Traumatic Brain Injury Provides Neuroprotection and Improves Cognitive Function in Rats. PLoS ONE 2010, 5, e11383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Tan, T.; Cao, Q.; Zhang, F.; Rein, B.; Duan, W.M.; Yan, Z. Histone Deacetylase Inhibition Restores Behavioral and Synaptic Function in a Mouse Model of 16p11.2 Deletion. Int. J. Neuropsychopharmacol. 2022, 25, 877–889. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Duration of Consumption | Temperature of Sample Preparation |
|---|---|---|
| Group 1 Control | 30 days | Room temperature |
| Group 2 Control | 90 days | Room temperature |
| Group 3 Control | 30 days | 160 °C |
| Group 4 Control | 90 days | 160 °C |
| Group 5 + TRZ | 30 days | Room temperature |
| Group 6 + TRZ | 90 days | Room temperature |
| Group 7 + TRZ | 30 days | 160 °C |
| Group 8 + TRZ | 90 days | 160 °C |
| Gene name | Forward Primer | Reverse Primer |
|---|---|---|
| DNA methyltransferase 1 (DNMT1) | AAGAATGGTGTTGTCTACCGAC | CATCCAGGTTGCTCCCCTTG |
| DNA methyltransferase 3A (DNMT3a) | GAGGGAACTGAGACCCCAC | CTGGAAGGTGAGTCTTGGCA |
| DNA methyltransferase 3B (DNMT3b) | AGCGGGTATGAGGAGTGCAT | GGGAGCATCCTTCGTGTCTG |
| Histone deacetylase 2 (HDAC2) | GGAGGAGGCTACACAATCCG | TCTGGAGTGTTCTGGTTTGTCA |
| Histone deacetylase 3 (HDAC3) | GCCAAGACCGTGGCGTATT | GTCCAGCTCCATAGTGGAAGT |
| Histone deacetylase 8 (HDAC8) | ACTATTGCCGGAGATCCAATGT | CCTCCTAAAATCAGAGTTGCCAG |
| Hypoxanthine phosphoribo-syltransferase 1 (HPRT1) | TCAGTCAACGGGGGACATAAA | GGGGCTGTACTGCTTAACCAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zand, A.; Enkhbilguun, S.; Macharia, J.M.; Budán, F.; Gyöngyi, Z.; Varjas, T. Tartrazine Modifies the Activity of DNMT and HDAC Genes—Is This a Link between Cancer and Neurological Disorders? Nutrients 2023, 15, 2946. https://doi.org/10.3390/nu15132946
Zand A, Enkhbilguun S, Macharia JM, Budán F, Gyöngyi Z, Varjas T. Tartrazine Modifies the Activity of DNMT and HDAC Genes—Is This a Link between Cancer and Neurological Disorders? Nutrients. 2023; 15(13):2946. https://doi.org/10.3390/nu15132946
Chicago/Turabian StyleZand, Afshin, Sodbuyan Enkhbilguun, John M. Macharia, Ferenc Budán, Zoltán Gyöngyi, and Timea Varjas. 2023. "Tartrazine Modifies the Activity of DNMT and HDAC Genes—Is This a Link between Cancer and Neurological Disorders?" Nutrients 15, no. 13: 2946. https://doi.org/10.3390/nu15132946
APA StyleZand, A., Enkhbilguun, S., Macharia, J. M., Budán, F., Gyöngyi, Z., & Varjas, T. (2023). Tartrazine Modifies the Activity of DNMT and HDAC Genes—Is This a Link between Cancer and Neurological Disorders? Nutrients, 15(13), 2946. https://doi.org/10.3390/nu15132946