Effects of Unconventional Work and Shift Work on the Human Gut Microbiota and the Potential of Probiotics to Restore Dysbiosis

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
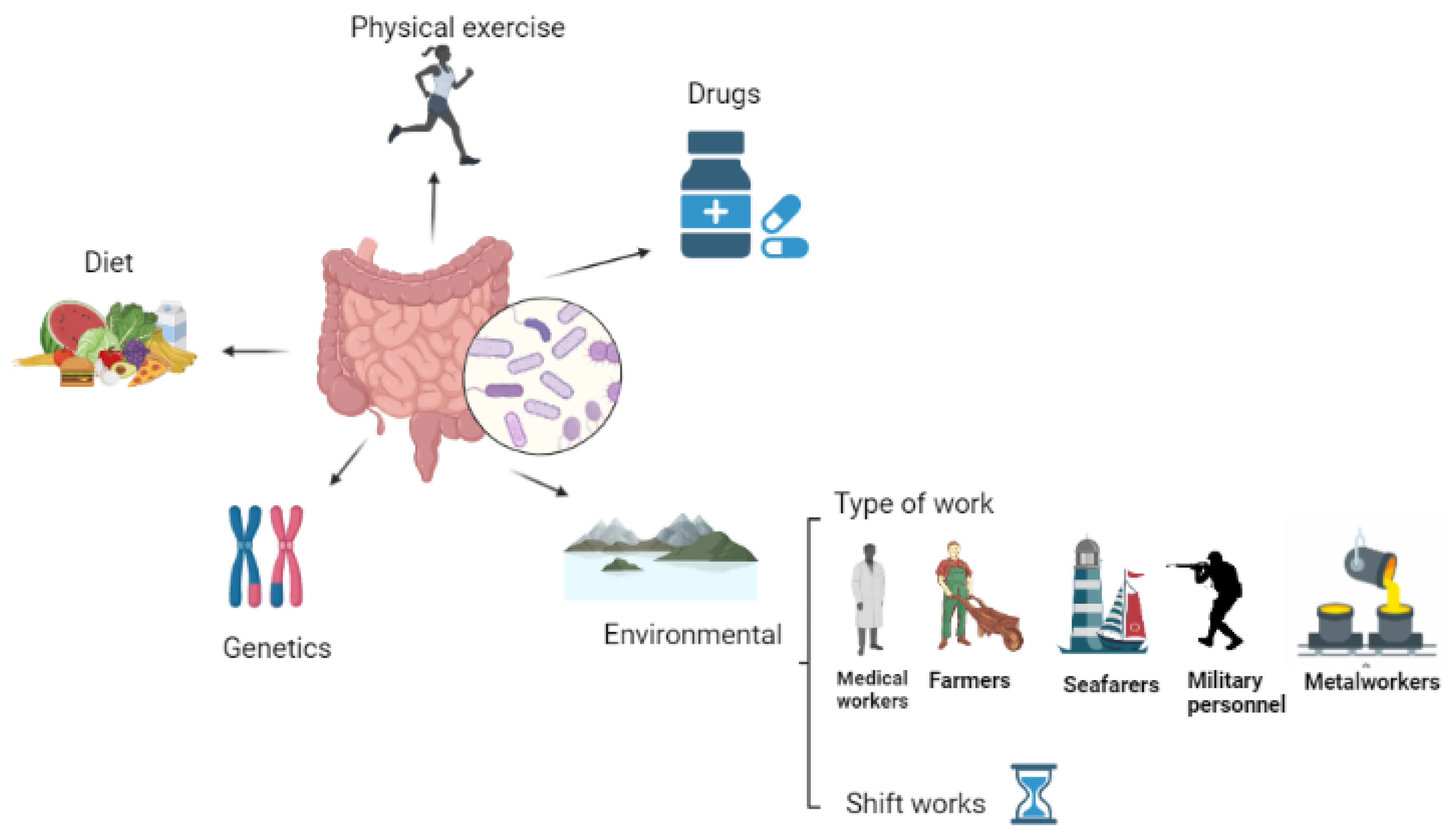
2. Effects of Work in Unconventional Schedules on Human Health
2.1. Changes in Dietary Pattern
2.2. Hormonal Changes
2.3. Sleep Disruption
2.4. Work Environment
3. Effects of Different Types of Unconventional Work on the Human Gut Microbiota
3.1. Healthcare Workers
3.2. Farm Workers
3.3. Military Personnel
3.4. Long-Travel Seamen
3.5. Metal and Tunnel-Workers
4. Use of Probiotics to Restore Dysbiosis in Workers

{kind=link}
| Type of trial | Subjects | Dosage and Time of Exposition | Effects on Gut Microbiota | Other Health Effects | Reference |
|---|---|---|---|---|---|
| Double-blind parallel-group trial | 94 shift workers | Lactobacillus acidophilus, Bifidobacterium animalis spp. lactis or placebo; 1 × 1010 colony count units (cfu) for 14 days | Not investigated | Probiotic supplementation decreased serum markers such as cortisol, pentraxin, or interleukin-1ra, related to sleep quality. | West et al. [22] |
| Randomized controlled trial | 961 women healthcare workers | 1.12 × 109 cfu or more of Lactobacillus bulgaricus OLL1073R-1 and strain of Streptococcus thermophilus daily for 16 weeks | Not investigated | A significant increase in interferon-γ production was found with a daily intake of OLL1073R-1 yogurt. | Kinoshita et al. [86] |
| A randomized, double-blind, placebo-controlled trial | 70 petrochemical workers | 100 g/day probiotic yogurt which contained two strains of Lactobacillus acidophilus LA5 and Bifidobacterium lactis BB12 with a total of min 1 × 107 cfu for 6 weeks | Not investigated | The consumption of probiotic yogurt had beneficial effects on mental health parameters in petrochemical workers. | Mohammadi et al. [87] |
| A double-blind, randomized, placebo-controlled experiment | 41 female healthcare workers employed on a rotating shift schedule | 4 g/day of freeze-dried powder of the multistrain probiotic mixture (2.5 × 109 cfu/g) for 6 weeks. The prebiotic mixture contained Bifidobacterium bifidum W23, Bifidobacterium lactis W51, Bifidobacterium lactis W52, Lactobacillus acidophilus W37, Lactobacillus brevis W63, Lactobacillus casei W56, Lactobacillus salivarius W24, and Lactococcus lactis (W19 and W58) | Not investigated | Results indicate a potential protective effect of probiotics against fat mass gain. Probiotics may alleviate anxiety and fatigue in shift-working females. | Smith-Ryan et al. [88] |
| A 30-day longitudinal experiment | 82 sailors during a sea voyage | 2 g package containing mixed probiotics including 9.70 Log cfu of Lactobacillus casei, 9.70 Log cfu of Lactobacillus plantarum P-8, 9.70 Log10 cfu of Lactobacillus rhamnosus M9, 9.88 Log10 cfu of Bifidobacterium lactis V9, and 9.88 Log10 cfu of Bifidobacterium lactis M8 once daily for 30 days | The compositions of the intestinal microbiota of the two groups (placebo and probiotic) were highly distinct at the end of the sea voyage, which confirmed the positive impacts of the probiotics consumed. | Probiotics maintained intestinal microbiome homeostasis and further prevented anxiety during the long sea voyage. | Zhang et al. [89] |
| A randomized, double-blind placebo-controlled study | 262 employees (day workers and three shift workers) | Daily dose of 108 cfu of L. reuteri for 80 days | Not investigated | Among the 53 shift workers, 33% in the placebo group reported being sick during the study period compared to none in the L. reuteri group. | Tubelius et al. [90] |
| Open-label single-arm study | 90 highly stressed information technology specialists | 300 mg of lyophilized L. plantarum PS128TM powder, which is equivalent to 10 billion cfu for 8 weeks | Not investigated | Significant improvements in self-perceived stress, overall job stress, job burden, cortisol level, general or psychological health, anxiety, depression, sleep disturbances, quality of life, and both positive and negative emotions | Wu et al. [73] |
| A controlled, parallel, randomized, and double-blind clinical trial | 65 military | Symbiotic ice cream containing: 2 × 108 cfu/g for L. acidophilus LA-5 and 2.7 × 109 cfu/g for B. animalis BB-12 and 2.3 g of inulin in the 60 g of ice cream for 30 days | No significant differences in α diversity between groups. No significant differences in the proportions of each phyla comparing the two groups. | This supplementation improved tenseness and sleepiness in healthy young military. | Valle et al. [91] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, Z.H.; Liu, Y.W.; Ji, Z.H.; Fu, T.; Yan, M.; Shao, Z.J.; Lona, Y. Alterations in the intestinal microbiome and mental health status of workers in an underground tunnel environment. BMC Microbiol. 2021, 21, 7. [Google Scholar] [CrossRef]
- Rogers, A.; Hu, Y.-J.; Yue, Y.; Wissel, E.F.; Petit, R.A., III; Jarrett, S.; Christie, J.; Read, T.D. Shiftwork, functional bowel symptoms, and the microbiome. Peer J. 2021, 9, e11406. [Google Scholar] [CrossRef]
- Bijnens, S.; Depoortere, I. Controlled light exposure and intermittent fasting as treatment strategies for metabolic syndrome and gut microbiome dysregulation in night shift works. Physiol. Behav. 2023, 263, 114103. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.J.; Bello, L.; Sánchez, A.; Serra, L. La turnicidad laboral y su impacto en la alimentación. In Alimentación y Trabajo; Editorial Médica Panamericana: Madrid, España, 2012; pp. 53–67. [Google Scholar]
- Lowden, A.; Moreno, C.; Holmback, U.; Lennernas, M.; Tucker, P. Eating and shift work—Effects on habits, metabolism and performance. Scand. J. Work Environ. Health 2010, 36, 150–162. [Google Scholar] [CrossRef]
- Parnell, J.A.; Reimer, R.A. Prebiotic fiber modulation of the gut microbiota improves risk factors for obesity and the metabolic syndrome. Gut Microbes 2012, 3, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tada, Y.; Kawano, Y.; Maeda, I.; Yoshizaki, T.; Sunami, A.; Yokoyama, Y.; Matsumoto, H.; Hida, A.; Komatsu, T.; Togo, F. Association of body mass index with lifestyle and rotating shift working Japanese female nurses. Obesity 2014, 22, 2489–2493. [Google Scholar] [CrossRef]
- Kim, H.I.; Choi, J.Y.; Kim, S.-E.; Jung, H.-K.; Shim, K.-N.; Yoo, K. Impact of shift work on irritable bowel syndrome and functional dyspepsia. Korean J. Med. Educ. 2013, 28, 431–437. [Google Scholar]
- Akere, A.; Akande, K.O. Association between irritable bowel syndrome and shift work: Prevalence and associated factors among nurses. J. Gastroenterol. Hepatol. 2014, 3, 1328–1331. [Google Scholar] [CrossRef] [Green Version]
- Kostic, A.D.; Gevers, D.; Siljander, H.; Vatanen, T.; Hyötyläinen, T.; Hämäläinen, A.-M.; Peet, A.; Tillmann, V.; Pöhö, P.; Mattila, I.; et al. The dynamics of the human infant gut microbiome in development and in progression towards type 1 diabetes. Cell Host Microbe 2015, 17, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson’s disease. Cell 2016, 167, 1469–1480. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, Z.; Lu, T.; Chen, W.; Yan, W.; Yuan, K.; Shi, L.; Liu, X.; Zhou, X.; Shi, J.; et al. The microbiota-gut-brain axis in sleep disorders. Sleep Med. Rev. 2022, 65, 101691. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Yuan, S.; Zhang, J. The interplay between sleep and gut microbiota. Brain Res. Bull. 2022, 180, 131–146. [Google Scholar] [CrossRef]
- Konturek, P.C.; Brzozowski, T.; Konturek, S.J. Gut clock: Implication of circadian rhythms in the gastrointestinal tract. J. Physiol. Pharmacol. 2011, 62, 139–150. [Google Scholar] [PubMed]
- Paulose, J.K.; Wright, J.M.; Patel, A.G.; Cassone, V.M. Human gut bacteria are sensitive to melatonin and express endogenous circadian rhythmicity. PLoS ONE 2016, 11, e0146643. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kuang, Z.; Yu, X.; Ruhn, K.A.; Kubo, M.; Hooper, L.V. The intestinal microbiota regulates body composition through NFIL3 and the circadian clock. Science 2017, 357, 912–916. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Yang, W.; Chen, G.; Men, C.; Gu, Y.; Song, X.; Zhang, R.; Wang, L.; Zhang, X. An important link between the gut microbiota and the circadian rhythm: Imply for treatments of circadian rhythm sleep disorder. Food Sci. Biotechnol. 2022, 31, 155–164. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Golubeva, A.V.; Crampton, S.; Desbonnet, L.; Edge, D.; O’Sullivan, O.; Lomasney, K.W.; Zhdanov, A.V.; Crispie, F.; Moloney, R.D.; Borre, Y.E.; et al. Prenatal stress-induced alterations in major physiological systems correlate with gut microbiota composition in adulthood. Psychoneuroendocrinology 2015, 60, 58–74. [Google Scholar] [CrossRef]
- Gu, F.; Han, J.; Laden, F.; Pan, A.; Caporaso, N.E.; Stampfer, M.J.; Kawachi, I.; Rexrode, K.M.; Willett, W.C.; Hankinson, S.E.; et al. Total and cause- specific mortality of U.S., nurses working rotating night shifts. Am. J. Prev. Med. 2015, 48, 241–252. [Google Scholar] [CrossRef] [Green Version]
- Myers, J.A.; Haney, M.F.; Griffiths, R.F.; Pierse, N.F.; Powell, D.M.C. Fatigue in air medical clinicians undertaking high-acuity patient transports. Prehosp. Emerg. Care 2015, 19, 36–43. [Google Scholar] [CrossRef]
- West, N.P.; Hughes, L.; Ramsey, R.; Zhang, P.; Martoni, C.J.; Leyer, G.J.; Cripps, A.W.; Cox, A.J. Probiotics, anticipation stress, and the acute immune response to night shift. Front. Immunol. 2021, 11, 599547. [Google Scholar] [CrossRef] [PubMed]
- Wells, M.M.; Roth, L.; Chande, N. Sleep disruption secondary to overnight call shifts is associated with irritable bowel syndrome in residents: A cross sectional study. Am. J. Gastroenterol. 2012, 107, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Zeevi, D.; Levy, M.; Zilberman-Shapira, G.; Suez, J.; Tengler, A.C.; Abramson, L.; Katz, M.N.; Korem, N.; Kuperman, Y.; et al. Transkingdom control of microbiota diurnal oscillations promotes metabolic homeostais. Cell 2014, 159, 514–529. [Google Scholar] [CrossRef] [Green Version]
- Nojkov, B.; Rubenstein, J.H.; Chey, W.D.; Hoogerwerf, W.A. The impact of rotating shift work on the prevalence of irritable bowel syndrome in nurses. Am. J. Gastroenterol. 2010, 105, 842–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattarai, Y.M.; Pedrogo, D.A.; Kashyap, P.C. Irritable bowel syndrome: A gut microbiota-related disorder. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, 52–62. [Google Scholar] [CrossRef]
- Wang, M.L.; Lin, P.L.; Huang, C.H.; Huang, H.H. Decreased parasympathetic activity of heart rate variability during anticipation of night duty in anesthesiology residents. Anesth. Analg. 2018, 126, 1013–1018. [Google Scholar] [CrossRef]
- Morris, S.B.; Daisley, R.L.; Wheeler, M.; Boyer, P. A meta-analysis of the relationship between individual assessments and job performance. J. App. Psychol. 2015, 100, 5–20. [Google Scholar] [CrossRef]
- Geda, N.R.; Feng, C.X.; Yu, Y. Examining the association between work stress, Life stress and obesity among working adult population in Canada: Findings from a Nationally Representative Data. Arch. Public Health 2022, 80, 97. [Google Scholar] [CrossRef]
- Magnet, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Briançon-Marjollet, A.; Weiszenstein, M.; Henri, M.; Thomas, A.; Godin-Ribuot, D.; Polak, J. The impact of sleep disorders on glucose metabolism: Endocrine and molecular mechanisms. Diabetol. Metab. Syndr. 2015, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Young, Y.H.; Kim, H.C.; Lim, H.C.; Park, J.J.; Kim, J.H.; Park, H. Long-term clinical course of post-infectious irritable bowel syndrome after shifellosis: A 10- year follow up study. J. Neurogastroenterol. Motil. 2016, 22, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Burek, K.; Rabstein, S.; Kantermann, T.; Vetter, C.; Rotter, M.; Wang-Sattler, R.; Lehnert, M.; Pallapies, D.; Jöckel, K.-H.; Brüning, T.; et al. Night work, chronotype and cortisol at awakening in female hospital employees. Sci. Rep. 2022, 12, 6525. [Google Scholar] [CrossRef] [PubMed]
- Besedovsky, L.; Lange, T.; Haack, M. The sleep-immune crosstalk in health and disease. Physiol. Rev. 2019, 99, 1325–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, M.P. A societal sleep prescription. Neuron 2019, 103, 559–562. [Google Scholar] [CrossRef] [PubMed]
- Szentirmai, É.; Kapás, L. Brown adipose tissue plays a central role in systemic inflammation-induced sleep responses. PLoS ONE 2018, 13, e0197409. [Google Scholar] [CrossRef] [Green Version]
- Vgontzas, A.N.; Papanicolaou, D.A.; Bixler, E.O.; Lotsikas, A.; Zachman, K.; Kales, A.; Prolo, P.; Wong, M.L.; Licinio, J.; Gold, P.W.; et al. Circadian interleukin-6 secretion and quantity and depth of sleep. J. Clin. Endocrinol. Metab. 1999, 84, 2603–2607. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.C.; Dorrian, J.; Liu, P.Y.; Van Dongen, H.P.; Wittert, G.A.; Harmer, L.J.; Banks, S. Impact of five nights of sleep restriction on glucose metabolism, leptin and testosterone in young adult men. PLoS ONE 2012, 7, e41218. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, K.; Tasali, E.; Penev, P.; Van Cauter, E. Brief communication: Sleep curtailment in healthy young men is associated with decreased leptin levels, elevated ghrelin levels, and increased hunger and appetite. Ann. Int. Med. 2004, 141, 846–850. [Google Scholar] [CrossRef]
- Shearer, W.T.; Reuben, J.M.; Mullington, J.M.; Price, N.J.; Lee, B.N.; Smith, E.O.; Szuba, M.P.; Van Dongen, H.P.; Dinges, D.F. Soluble TNF alpha receptor 1 and IL-6 plasma levels in humans subjected to the sleep deprivation model of spaceflight. J. Allergy Clin. Immunol. 2001, 107, 165–170. [Google Scholar] [CrossRef]
- Litichevskiy, L.; Thaiss, C.A. The Oscillating Gut Microbiome and Its Effects on Host Circadian Biology. Ann. Rrev. Nutr. 2022, 42, 145–164. [Google Scholar] [CrossRef]
- Wollmuth, E.M.; Angert, E.R. Microbial circadian clocks: Host-microbe interplay in diel cycles. BMC Microbiol. 2023, 23, 124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.I.; Bai, L.; Goel, N.; Bailey, A.; Jang, C.J.; Bushman, F.D.; Meerlo, P.; Dinges, D.F.; Sehgal, A. Human and rat gut microbiome composition is maintained following sleep restriction. Proc. Natl. Acad. Sci. USA 2017, 114, E1564–E1571. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.; Vogel, H.; Jonas, W.; Woting, A.; Blaut, M.; Schurmann, A.; Cedernaes, J. Gut microbiota and glucometabolic alterations in response to recurrent particle sleep deprivation in normal-weight young individuals. Mol. Metab. 2016, 5, 1175–1186. [Google Scholar] [CrossRef]
- Voigt, R.M.; Forsyth, C.B.; Green, S.J.; Mutlu, E.; Engen, P.; Vitaterna, M.H.; Turek, F.W.; Keshavarizian, A. Circadian disorganization alters intestinal microbiota. PLoS ONE 2014, 9, e97500. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Fang, D.; Wang, Z.; Liu, Y. Sleep Deprivation and Gut Microbiota Dysbiosis: Current Understandings and Implications. Int. J. Mol. Sci. 2023, 24, 9603. [Google Scholar] [CrossRef]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liao, X.P.; D’Souza, A.W.; Boolchandani, M.; Li, S.-H.; Cheng, K.; Luis Martínez, J.; Li, L.; Feng, Y.J.; Fang, L.X.; et al. Environmental remodeling of human gut microbiota and antibiotic resistome in livestock farms. Nat. Commun. 2020, 11, 1427. [Google Scholar] [CrossRef] [Green Version]
- Poza, M.; Gayoso, C.; Gómez, M.J.; Rumbo-Feal, S.; Tomás, M.; Aranda, J.; Fernández, A.; Bou, G. Exploring bacterial diversity in hospital environments by GS-FLX Titanium pyrosequencing. PLoS ONE 2012, 7, e44105. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Tu, C.C.; Kuo, H.Y.; Zeng, R.F.; Yu, C.S.; Lu, H.H.S.; Liou, M.L. Dynamic change of surface microbiota with different environmental cleaning methods between two wards in a hospital. Appl. Microbiol. Biotechnol. 2016, 101, 771–781. [Google Scholar] [CrossRef]
- Brooks, B.; Olm, M.R.; Firek, B.A.; Baker, R.; Thomas, B.C.; Morowitz, M.J.; Banfield, J.F. Strain- resolved analysis of hospital rooms and infants reveals overlap between the human and room microbiome. Nat. Commun. 2017, 8, 1814. [Google Scholar] [CrossRef] [Green Version]
- Lai, P.S.; Allen, J.G.; Hutchinson, D.S.; Ajami, N.J.; Petrosino, J.F.; Winters, T.; Hug, C.; Wartenberg, G.R.; Vallarino, J.; Christiani, D.C. Impact of environmental microbiota on human microbiota of workers in academic mouse research facilities: An observational study. PLoS ONE 2017, 12, e0180969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, N.D.; Pollard, J.; Stupart, D.; Knight, D.R.; Khajehnoori, M.; Davey, E.K.; Parry, L.; Riley, T.V. Prevalence of Clostridium difficile colonization among healthcare workers. BMC Infect. Dis. 2013, 13, 459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huttunen, R.; Syrjanen, J. Healthcare workers as vectors of infectious diseases. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1477–1488. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Huang, T.; Chen, C.; Cao, T.T.; Cheng, K.; Liao, X.P.; Liu, Y.H. Comparison of fecal microbial composition and antibiotic resi stance genes from swine, farm workers and the surrounding villagers. Sci. Rep. 2017, 7, 4965. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Guo, R.; Ma, Q.; Li, Y.; Wang, W.; Fan, Y.; Ju, Y.; Zhao, B.; Gao, Y.; Qian, L.; et al. Stressful events induce long-term gut microbiota dysbiosis and associated post-traumatic stress symptoms in healthcare workers fighting against COVID-19. J. Affect. Disord. 2022, 303, 187–195. [Google Scholar] [CrossRef]
- Zheng, N.; Li, S.H.; Dong, B.; Sun, W.; Li, H.R.; Zhang, Y.L.; Li, P.; Fang, Z.W.; Chen, C.M.; Han, X.Y.; et al. Comparison of the gut microbiota of short-term and long-term medical workers and non-medical controls: A cross-sectional analysis. Clin. Microbiol. Infect. 2021, 27, 1285–1292. [Google Scholar] [CrossRef]
- Mortaş, H.; Bilici, S.; Karakan, T. The circadian disruption of night work alters gut microbiota consistent with elevated risk for future metabolic and gastrointestinal pathology. Chronobiol. Internat. 2020, 37, 1067–1081. [Google Scholar] [CrossRef]
- Cerf-Bensussan, N.; Gaboriau-Routhiau, V. The immune system and the gut microbiota: Friends or foes? Nat. Rev. Immunol. 2010, 10, 735–744. [Google Scholar] [CrossRef]
- Tahara, Y.; Yamazaki, M.; Sukigara, H.; Motohashi, H.; Sasaki, H.; Miyakawa, H. Gut microbiota-derived short chain fatty acids induce circadian clock entrainment in mouse peripheral tissue. Sci. Rep. 2018, 8, 1395. [Google Scholar] [CrossRef] [Green Version]
- Stanaway, I.B.; Wallace, J.C.; Shojaie, A.; Griffith, W.C.; Hong, S.; Wilder, C.S.; Green, F.H.; Tsai, J.; Knight, M.; Workman, T.; et al. Human oral buccal microbiomes are associated with farmworker status and azinphos-methyl agricultural pesticide exposure. Appl. Environ. Microbiol. 2017, 83, e02149-16. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Lin, H.; Meng, Y.; Zhu, J.; Zhang, Y.; Zhang, L.; Li, G. Analysis and determinants of Chinese navy personnel health status: A cross-sectional study. Health Qual. Life Outcomes 2018, 16, 138. [Google Scholar] [CrossRef] [Green Version]
- Walters, W.A.; Reyes, F.; Soto, G.M.; Reynolds, N.D.; Fraser, J.A.; Aviles, R.; Tribble, D.R.; Irvin, A.P.; Kelley-Loughnane, N.; Gutierrez, R.L.; et al. Epidemiology and associated microbiota changes in deployed military personnel at high risk of traveler’s Diarrhea. PLoS ONE 2020, 15, e0236703. [Google Scholar] [CrossRef] [PubMed]
- Rydstedt, L.W.; Lundh, M. An ocean of stress? The relationship between psychosocial workload and mental strain among engine officers in the Swedish merchant fleet. Int. Marit. Health 2010, 62, 168–175. [Google Scholar] [PubMed]
- O’Halloran, C.L.; Silver, M.W.; Colford, J.M., Jr. Acute stress symptoms among US ocean lifeguards. Wilderness Environ. Med. 2015, 26, 442–443. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Price, J.; Mahurkar, A.; Rahnavard, G.; Crabtree, J.; Orvis, J.; Hall, A.B.; Brady, A.; Creasy, H.H.; McCracken, C.; Giglio, M.G.; et al. Strains, functions and dynamics in the expanded Human Microbiome Project. Nature 2017, 550, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Zhang, Z.; Liu, C.; Qiao, Y.; Zhou, D.; Qu, J.; An, H.; Xiong, M.; Zhu, Z.; Zhao, X. Metagenomic sequencing reveals altered metabolic pathways in the oral microbiota of sailors during a long sea voyage. Sci Rep. 2015, 5, 9131. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zhao, J.; Jin, H.; Jin, H.; Lv, R.; Shi, H.; De, G.; Yang, B.; Sun, Z.; Zhang, H. Probiotics maintain the intestinal microbiome homeostasis of the sailors during a long sea voyage. Gut Microbes 2020, 11, 930–943. [Google Scholar] [CrossRef]
- Kriss, M.; Hazleton, K.Z.; Nusbacher, N.M.; Martin, C.G.; Lozupone, C.A. Low diversity gut microbiota dysbiosis: Drivers, functional implications and recovery. Curr. Opin. Microbiol. 2018, 44, 4–40. [Google Scholar] [CrossRef]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 65–983. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Chen, L.; Sun, G.; Li, Y.; Huang, R. Alterations in the gut microbiota of patients with silica-induced pulmonary fibrosis. J. Occup. Med. Toxicol. 2019, 14, 5. [Google Scholar] [CrossRef]
- Ahmed, N.; Mahmoud, N.F.; Solyman, S.; Hanora, A. Human nasal microbiome as characterized by metagenomics differs markedly between rural and industrial communities in Egypt. Omics J. Integr. Biol. 2019, 23, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.G.; Kapoor, B.; Cummings, K.J.; Stanton, M.L.; Nett, R.J.; Kreiss, K.; Abraham, J.L.; Colby, T.V.; Franko, A.D.; Green, F.H.; et al. Evidence for environmental–human microbiota transfer at a manufacturing facility with novel work-related respiratory disease. Am. J. Respir. Crit. Care Med. 2020, 202, 1678–1688. [Google Scholar] [CrossRef] [PubMed]
- Cummings, K.J.; Stanton, M.L.; Nett, R.J.; Segal, L.N.; Kreiss, K.; Abraham, J.L.; Colby, T.V.; Franko, A.D.; Green, F.H.Y.; Sanyal, S.; et al. Severe lung disease characterized by lymphocytic bronchiolitis, alveolar ductitis, and emphysema (BADE) in industrial machine-manufacturing workers. Am. J. Ind. Med. 2019, 62, 927–937. [Google Scholar] [CrossRef]
- Le Guern, R.; Stabler, S.; Gosset, P.; Pichavant, M.; Grandjean, T.; Faure, E.; Karaca, Y.; Faure, K.; Kipnis, E.; Dessein, R. Colonization resistance against multi-drug-resistant bacteria: A narrative review. J. Hosp. Infect. 2021, 118, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Derrien, M.; van Hylckama Vlieg, J.E. Fate, activity, and impact of ingested bacteria within the human gut microbiota. Trends Microbiol. 2015, 23, 354–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, B.; Delgado, S.; Blanco-Miguez, A.; Lourenco, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.H.; Nam, M.H.; Lee, H.; Lee, J.S.; Kim, H.; Chung, M.J.; Seo, J.G. Amelioration of obesity-related characteristics by a probiotic formulation in a high-fat diet-induced obese rat model. Eur. J. Nutr. 2017, 57, 2081–2090. [Google Scholar] [CrossRef]
- Xu, H.; Huang, W.; Hou, Q.; Kwok, L.Y.; Laga, W.; Wang, Y.; Ma, H.; Sun, Z.; Zhang, H. Oral administration of com- pound probiotics improved canine feed intake, weight gain, immunity and intestinal microbiota. Front Immunol. 2019, 10, 666. [Google Scholar] [CrossRef]
- Huang, L.; Yu, Q.; Peng, H.; Zhen, Z. Alterations of gut microbiome and effects of probiotic therapy in patients with liver cirrhosis: A systematic review and meta-analysis. Medicine 2022, 101, e32335. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.X.; Niu, H.T.; Zhang, S.Y. Intestinal microbiota metabolism and atherosclerosis. Chin. Med. J. 2015, 128, 2805–2811. [Google Scholar] [CrossRef] [PubMed]
- Nallu, A.; Sharma, S.; Ramezani, A.; Muralidharan, J.; Raj, D. Gut microbiome in chronic kidney disease: Challenges and opportunities. Transl. Res. 2017, 179, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Wlodarska, M.; Kostic, A.D.; Xavier, R.J. An integrative view of microbiome-host interactions in inflammatory bowel diseases. Cell Host Microbe 2015, 17, 577–591. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Santamarina, A.; Gonzalez, E.G.; Lamas, A.; Mondragon, A.C.; Regal, P.; Miranda, J.M. Probiotics as a possible strategy for the prevention and treatment of allergies. A narrative review. Foods 2021, 10, 701. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Maruyama, K.; Suyama, K.; Nishijima, M.; Akamatsu, K.; Jogamoto, A.; Katakami, K.; Saito, I. The effects of OLL1073R-1 yogurt intake on influenza incidence and immunological markers among women healthcare workers: A randomized controlled trial. Food Funct. 2019, 10, 8129–8136. [Google Scholar] [CrossRef]
- Mohammadi, A.A.; Jazayeri, S.; Khosravi-Darani, K.; Solati, Z.; Mohammadpour, N.; Asemi, Z.; Adab, Z.; Djalali, M.; Tehrani-Doost, M.; Hosseini, M.; et al. The effects of probiotics on mental health and hypothalamic–pituitary–adrenal axis: A randomized, double-blind, placebo-controlled trial in petrochemical workers. Nutr. Neuro 2015, 19, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Smith-Ryan, A.E.; Mock, M.G.; Trexler, E.T.; Hirsch, K.R.; Blue, M.N.M. Influence of a multistrain probiotic on body composition and mood in female occupational shift workers. Appl. Physiol. Nutr. Metab. 2019, 44, 765–773. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, Z.; Jiang, S.; Bai, X.; Ma, C.; Peng, Q.; Chen, K.; Chang, H.; Fang, T.; Zhang, H. Probiotic Bifidobacterium lactis V9 regulates the secretion of sex hormones in polycystic ovary syndrome patients through the gut-brain axis. mSystems 2019, 4, e00017-19. [Google Scholar] [CrossRef] [Green Version]
- Tubelius, P.; Stan, V.; Zachrisson, A. Increasing work-place healthiness with the probiotic lactobacillus reuteri: A randomised, double-blind placebo-controlled study. Environm. Health 2005, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Valle, M.C.; Vieira, I.A.; Fino, L.C.; Gallina, D.A.; Esteves, A.M.; da Cunha, D.T.; Cabral, L.; Benatti, F.B.; Marostica, M.R., Jr.; Batista, Â.G.; et al. Immune status, well-being and gut microbiota in military supplemented with synbiotic ice cream and submitted to field training: A randomised clinical trial. Br. J. Nutr. 2021, 126, 1794–1808. [Google Scholar] [CrossRef]
- Vemuri, R.; Shinde, T.; Gundamaraju, R.; Gondalia, S.; Karpe, A.; Beale, D.; Martoni, C.; Eri, R. Lactobacillus acidophilus DDS-1 modulates the gut microbiota and improves metabolic profiles in aging mice. Nutrients 2018, 10, 1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martoni, C.J.; Srivastava, S.; Leyer, G.J. Lactobacillus acidophilus DDS-1 and Bifidobacterium lactis UABla-12 improve abdominal pain severity and symptomology in irritable bowel syndrome: Randomized controlled trial. Nutrients 2020, 12, 363. [Google Scholar] [CrossRef] [Green Version]
- Gerasimov, S.V.; Vasjuta, V.V.; Myhovych, O.O.; Bondarchuk, L.I. Probiotic supplement reduces atopic dermatitis in preschool children. Am. J. Clin. Dermatol. 2010, 11, 351–361. [Google Scholar] [CrossRef]
- Pakdaman, M.N.; Udani, J.K.; Molina, J.P.; Shahani, M. The effects of the DDS-1 strain of lactobacillus on symptomatic relief for lactose intolerance—A randomized, double-blind, placebo-controlled, crossover clinical trial. Nutr. J. 2015, 15, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irwin, C.; McCartney, D.; Desbrow, B.; Khalesi, S. Effects of probiotics and paraprobiotics on subjective and objective sleep metrics: A systematic review and meta-analysis. Eur. J. Clin. Nutr. 2020, 74, 1536–1549. [Google Scholar] [CrossRef]
- Gerasimov, S.V.; Ivantsiv, V.A.; Bobryk, L.M.; Tsitsura, O.O.; Dedyshin, L.P.; Guta, N.V.; Yandyo, B.V. Role of short-term use of L. acidophilus DDS-1 and B. lactis UABLA-12 in acute respiratory infections in children: A randomized controlled trial. Eur. J. Clin. Nutr. 2015, 70, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Labanki, A.; Langhorst, J.; Engler, H.; Elsenbruch, S. Stress and the brain-gut axis in functional and chronic-inflammatory gastrointestinal diseases: A transdisciplinary challenge. Psychoneuroendocrinology 2020, 111, 104501. [Google Scholar] [CrossRef] [PubMed]
- Casper, A.; Sonnentag, S. Feeling exhausted or vigorous in anticipation of high workload? The role of worry and planning during the evening. J. Occup. Org. Psychol. 2020, 93, 215–242. [Google Scholar] [CrossRef]
- Codoñer-Franch, P.; Gomberto, M.; Martínez-Raga, J.; Cenit, M.C. Circadian Disruption and Mental Health: The Chronotherapeutic Potential of Microbiome-Based and Dietary Strategies. Int. J. Mol. Sci. 2023, 24, 7579. [Google Scholar] [CrossRef]
- Kaczmarek, J.L.; Musaad, S.M.; Holster, H.D. Time of day and eating behaviors are associated with the composition and function of the human gastrointestinal microbiota. Am. J. Clin. Nutr. 2017, 106, 1220–1231. [Google Scholar] [CrossRef] [Green Version]
- Haarhuis, J.E.; Kardinaal, A.; Kortman, G.A.M. Probiotics, prebiotics and postbiotics for better sleep quality: A narrative review. Benef. Microbes 2022, 13, 169–182. [Google Scholar] [CrossRef] [PubMed]

| Workers | Subjects | Dosage and Time of Exposition | Effects of Gut Microbiota | Other Health Effects | Reference |
|---|---|---|---|---|---|
| Tunnel miners | 48 healthy men | Before and after 3 weeks of working in a tunnel 8 h/day | Decreased GM diversity; increased Actinobacteria and Bifidobacteriales, Corynebacteriales and Desulfovibrionales; increased Bifidobacterium, Romboutsia, Clostridium, and Leucobacter; decreasing Faecalibacterium and Roseburia | Decreased antioxidant efficacy, digestive and absorptive capacity; increased proinflammatory factors | Lu et al. [1] |
| Healthcare workers | 214 workers | Fecal samples were subjected to an enzyme immunoassay for toxins A and b and for glutamate dehydrogenase | Only 0.8% of healthcare workers were found positive for C. difficile toxins and antigen | The results found did not confirm the hypothesis that a long stay in the hospital is a risk of C. difficile infection | Friedman et al. [53] |
| Farm workers | 6 swine farm workers and 6 local villagers | 16S rRNA gene sequencing of fecal samples | Workers had less species diversity compared to the local villagers, as well as higher amounts of Proteobacteria and Clostridiaceae | Analysis of antimicrobial resistance genes did not reveal significant differences among workers and villagers | Sun et al. [48] |
| Veterinary students in swine farms | 14 students who stayed 3 months on swine farms | 91 fecal samples investigated by 16S rRNA and whole metagenome shotgun | Moderate decrease in Bacteroidetes and an increase in Proteobacteria phyla | Antibiotic resistance genes were found in similar genetic contexts in student samples and farm environmental samples | Sun et al. [55] |
| Healthcare workers | 71 frontline healthcare workers fighting against COVID-19 | A longitudinal investigation at four time points: immediately after they finished treatment and left the isolation wards (Day 0), after a two-week quarantine in a hotel (Day 14), four weeks after their return to normal life (Day 45), and a half year after the frontline work (Day 180) | Microbes associated with mental health were mainly Faecalibacterium spp. and Eubacterium eligens group spp. Of note, the prediction model indicated that a low abundance of Eubacterium hallii group uncultured bacterium and a high abundance of Bacteroides eggerthii immediately after the two-month frontline work were significant determinants of the reappearance of post-traumatic stress symptoms. | Stressful events induced significant depression, anxiety, and stress | Gao et al. [56] |
| Medical workers | 175 healthy medical workers | Short-term (1–3 months) workers (n = 80) and long-term (>1 year) workers (n = 95) | Short-term workers: significantly higher abundances of Lactobacillus, Butyrivibrio, Clostridiaceae, Clostridium, Ruminococcus, Dialister, Bifidobacterium, Odoribacter, and Desulfovibrio, and lower abundances of Bacteroides and Blautia. Long-term workers: higher abundances of taxa such as Dialister, Veillonella, Clostridiaceae, Clostridium, Bilophila, Desulfovibrio, Pseudomonas, and Akkermansia, and lower abundances of Bacteroides and Coprococcus | Not investigated | Zheng et al. [57] |
| Nurses | 51 full-time staff nurses | Worked 12 h day or night shifts | No differences in the richness and diversity of species in samples from nurses working day and night shifts | Not investigated | Rogers et al. [2] |
| Security officers | 10 male security officers working rotational day/night shifts | After working the day shift (7:00 h–15: 00 h) for 4 weeks and after working the night shift (23:00 h–7:00 h) for 2 weeks. One-off day per week | The phylum with the highest abundance in the day shift was Firmicutes, followed by Bacteroidetes. In the night shift, the relative abundance of Bacteroidetes decreased; however, Actinobacteria and Firmicutes increased. Faecalibacterium was found to be a biomarker of day-shift work. | Not investigated | Mortaᶊ et al. [58] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez-Santamarina, A.; Mondragon, A.d.C.; Cardelle-Cobas, A.; Santos, E.M.; Porto-Arias, J.J.; Cepeda, A.; Miranda, J.M. Effects of Unconventional Work and Shift Work on the Human Gut Microbiota and the Potential of Probiotics to Restore Dysbiosis. Nutrients 2023, 15, 3070. https://doi.org/10.3390/nu15133070
Lopez-Santamarina A, Mondragon AdC, Cardelle-Cobas A, Santos EM, Porto-Arias JJ, Cepeda A, Miranda JM. Effects of Unconventional Work and Shift Work on the Human Gut Microbiota and the Potential of Probiotics to Restore Dysbiosis. Nutrients. 2023; 15(13):3070. https://doi.org/10.3390/nu15133070
Chicago/Turabian StyleLopez-Santamarina, Aroa, Alicia del Carmen Mondragon, Alejandra Cardelle-Cobas, Eva Maria Santos, Jose Julio Porto-Arias, Alberto Cepeda, and Jose Manuel Miranda. 2023. "Effects of Unconventional Work and Shift Work on the Human Gut Microbiota and the Potential of Probiotics to Restore Dysbiosis" Nutrients 15, no. 13: 3070. https://doi.org/10.3390/nu15133070
APA StyleLopez-Santamarina, A., Mondragon, A. d. C., Cardelle-Cobas, A., Santos, E. M., Porto-Arias, J. J., Cepeda, A., & Miranda, J. M. (2023). Effects of Unconventional Work and Shift Work on the Human Gut Microbiota and the Potential of Probiotics to Restore Dysbiosis. Nutrients, 15(13), 3070. https://doi.org/10.3390/nu15133070