
Milk-Fat-Globule-Membrane-Enriched Dairy Milk Compared with a Soy-Lecithin-Enriched Beverage Did Not Adversely Affect Endotoxemia or Biomarkers of Gut Barrier Function and Cardiometabolic Risk in Adults with Metabolic Syndrome: A Randomized Controlled Crossover Trial

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participant Eligibility
2.2. Study Design
2.3. Test Beverage Formulation and Compliance to Test Beverages
2.4. Dietary Control and Assessment
2.5. Metabolic Health
2.6. Endotoxemia
2.7. Systemic and Intestinal Inflammation
2.8. Intestinal Permeability
2.9. Short-Chain Fatty Acids
2.10. Statistical Analysis
3. Results
3.1. Participants
3.2. Test Beverage and Diet Compliance
3.3. Anthropometrics, Blood Pressure, and Fasting Cardiometabolic Markers
3.4. Postprandial Excursions in Cardiometabolic Markers
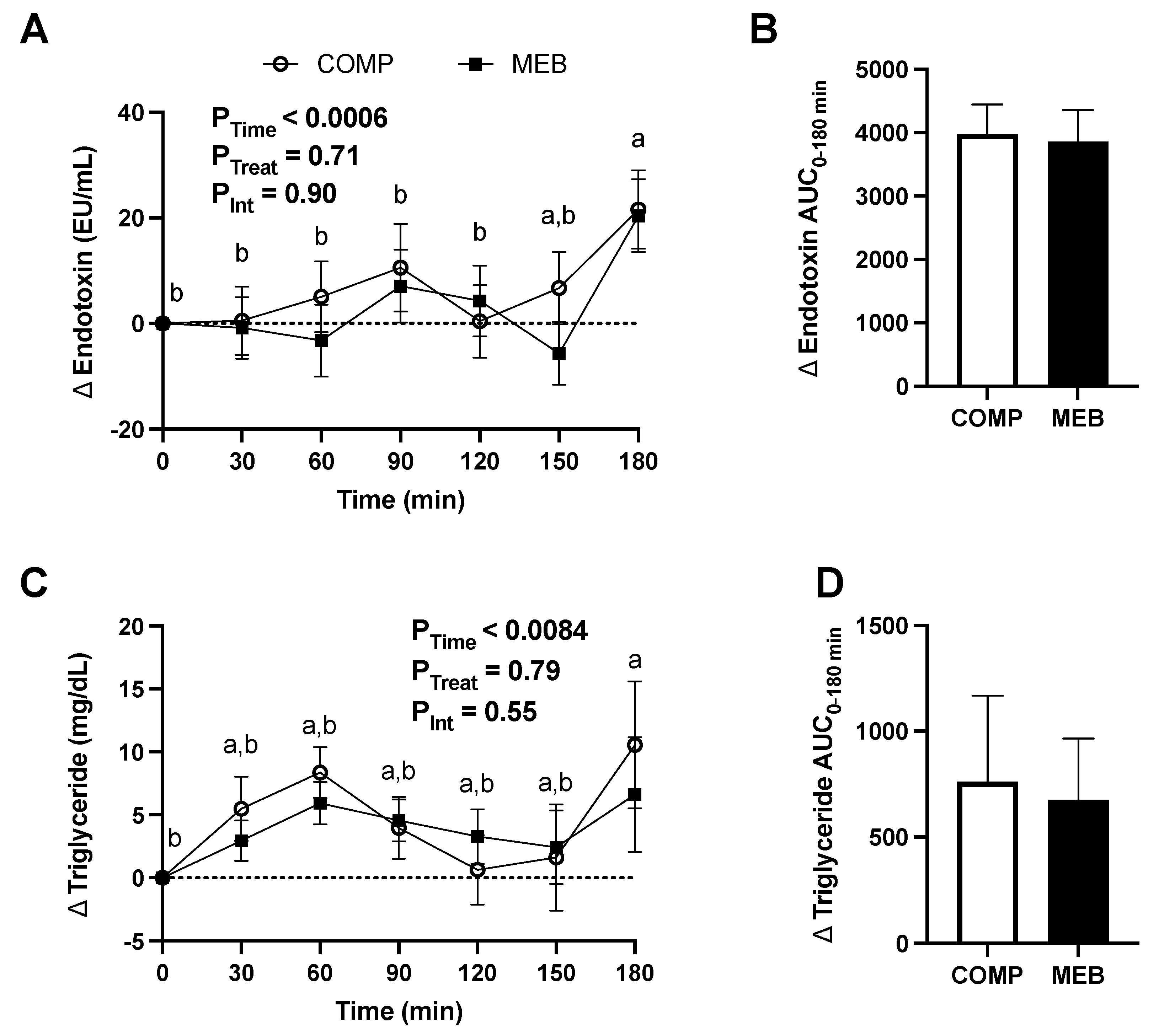
3.5. Gut Barrier Function and Inflammation
3.6. Systemic Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguilar, M.; Bhuket, T.; Torres, S.; Liu, B.; Wong, R.J. Prevalence of the metabolic syndrome in the United States, 2003–2012. JAMA 2015, 313, 1973–1974. [Google Scholar] [CrossRef] [Green Version]
- Ho, J.S.; Cannaday, J.J.; Barlow, C.E.; Mitchell, T.L.; Cooper, K.H.; FitzGerald, S.J. Relation of the number of metabolic syndrome risk factors with all-cause and cardiovascular mortality. Am. J. Cardiol. 2008, 102, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Traber, M.G.; Buettner, G.R.; Bruno, R.S. The relationship between vitamin C status, the gut-liver axis, and metabolic syndrome. Redox Biol. 2019, 21, 101091. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Creely, S.J.; McTernan, P.G.; Kusminski, C.M.; Fisher, F.; Da Silva, N.F.; Khanolkar, M.; Evans, M.; Harte, A.L.; Kumar, S. Lipopolysaccharide activates an innate immune system response in human adipose tissue in obesity and type 2 diabetes. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E740–E747. [Google Scholar] [CrossRef] [Green Version]
- Martel, J.; Chang, S.-H.; Ko, Y.-F.; Hwang, T.-L.; Young, J.D.; Ojcius, D.M. Gut barrier disruption and chronic disease. Trends Endocrinol. Metab. 2022, 33, 247–265. [Google Scholar] [CrossRef]
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2020 Update: A Report From the American Heart Association. Circulation 2020, 141, e139–e596. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Levy, M.; Grosheva, I.; Zheng, D.; Soffer, E.; Blacher, E.; Braverman, S.; Tengeler, A.C.; Barak, O.; Elazar, M.; et al. Hyperglycemia drives intestinal barrier dysfunction and risk for enteric infection. Science 2018, 359, 1376–1383. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.C.; Szeto, I.M.; Chen, L.H.; Han, S.F.; Li, Y.J.; van Hekezen, R.; Qin, L.Q. Dairy products consumption and metabolic syndrome in adults: Systematic review and meta-analysis of observational studies. Sci. Rep. 2015, 5, 14606. [Google Scholar] [CrossRef]
- Kratz, M.; Baars, T.; Guyenet, S. The relationship between high-fat dairy consumption and obesity, cardiovascular, and metabolic disease. Eur. J. Nutr. 2013, 52, 1–24. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Grieger, J.A.; Hilpert, K.F.; West, S.G. Milk products, dietary patterns and blood pressure management. J. Am. Coll. Nutr. 2009, 28, 103S–119S. [Google Scholar] [CrossRef]
- Ballard, K.D.; Bruno, R.S. Protective role of dairy and its constituents on vascular function independent of blood pressure-lowering activities. Nutr. Rev. 2015, 73, 36–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, J.D.; Mah, E.; Dey, P.; Olmstead, B.D.; Sasaki, G.Y.; Villamena, F.A.; Bruno, R.S. Dairy milk, regardless of fat content, protects against postprandial hyperglycemia-mediated impairments in vascular endothelial function in adults with prediabetes by limiting oxidative stress responses that reduce nitric oxide bioavailability. J. Nutr. Biochem. 2019, 63, 129–139. [Google Scholar] [CrossRef]
- Bhavadharini, B.; Dehghan, M.; Mente, A.; Rangarajan, S.; Sheridan, P.; Mohan, V.; Iqbal, R.; Gupta, R.; Lear, S.; Wentzel-Viljoen, E.; et al. Association of dairy consumption with metabolic syndrome, hypertension and diabetes in 147,812 individuals from 21 countries. BMJ Open. Diabetes Res. Care 2020, 8, e000826. [Google Scholar] [CrossRef]
- Bruno, R.S.; Pokala, A.; Torres-Gonzalez, M.; Blesso, C.N. Cardiometabolic health benefits of dairy-milk polar lipids. Nutr. Rev. 2021, 79, 16–35. [Google Scholar] [CrossRef] [PubMed]
- Timby, N.; Domellöf, M.; Lönnerdal, B.; Hernell, O. Supplementation of infant formula with bovine milk fat globule membranes. Adv. Nutr. 2017, 8, 351–355. [Google Scholar] [CrossRef] [Green Version]
- Norris, G.H.; Jiang, C.; Ryan, J.; Porter, C.M.; Blesso, C.N. Milk sphingomyelin improves lipid metabolism and alters gut microbiota in high fat diet-fed mice. J. Nutr. Biochem. 2016, 30, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Palmano, K.P.; MacGibbon, A.K.H.; Gunn, C.A.; Schollum, L.M. In vitro and in vivo anti-inflammatory activity of bovine milkfat globule (MFGM)-derived complex lipid fractions. Nutrients 2020, 12, 2089. [Google Scholar] [CrossRef] [PubMed]
- Snow, D.R.; Ward, R.E.; Olsen, A.; Jimenez-Flores, R.; Hintze, K.J. Membrane-rich milk fat diet provides protection against gastrointestinal leakiness in mice treated with lipopolysaccharide. J. Dairy. Sci. 2011, 94, 2201–2212. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Zhang, X.; Han, D.; Pi, Y.; Tao, S.; Zhang, S.; Wang, S.; Zhao, J.; Chen, L.; Wang, J. Early life administration of milk fat globule membrane promoted SCFA-producing bacteria colonization, intestinal barriers and growth performance of neonatal piglets. Anim. Nutr. 2021, 7, 346–355. [Google Scholar] [CrossRef]
- Quarles, W.R.; Pokala, A.; Shaw, E.L.; Ortega-Anaya, J.; Hillmann, L.; Jimenez-Flores, R.; Bruno, R.S. Alleviation of Metabolic Endotoxemia by Milk Fat Globule Membrane: Rationale, Design, and Methods of a Double-Blind, Randomized, Controlled, Crossover Dietary Intervention in Adults with Metabolic Syndrome. Curr. Dev. Nutr. 2020, 4, nzaa130. [Google Scholar] [CrossRef]
- Alberti, K.G.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.; Loria, C.M.; Smith, S.C., Jr. Harmonizing the metabolic syndrome: A joint interim statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Troy, L.M.; Rogers, G.T.; Fox, C.S.; McKeown, N.M.; Meigs, J.B.; Jacques, P.F. Longitudinal association between dairy consumption and changes of body weight and waist circumference: The Framingham Heart Study. Int. J. Obes. 2014, 38, 299–305. [Google Scholar] [CrossRef] [Green Version]
- Boutagy, N.E.; McMillan, R.P.; Frisard, M.I.; Hulver, M.W. Metabolic endotoxemia with obesity: Is it real and is it relevant? Biochimie 2016, 124, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Erridge, C.; Attina, T.; Spickett, C.M.; Webb, D.J. A high-fat meal induces low-grade endotoxemia: Evidence of a novel mechanism of postprandial inflammation. Am. J. Clin. Nutr. 2007, 86, 1286–1292. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.J.; Heaton, K.W. Stool form scale as a useful guide to intestinal transit time. Scand. J. Gastroenterol. 1997, 32, 920–924. [Google Scholar] [CrossRef] [PubMed]
- Rombaut, R.; Dewettinck, K.; Van Camp, J. Phospho- and sphingolipid content of selected dairy products as determined by HPLC coupled to an evaporative light scattering detector (HPLC–ELSD). J. Food Compost. Anal. 2007, 20, 308–312. [Google Scholar] [CrossRef]
- Cohn, J.S.; Kamili, A.; Wat, E.; Chung, R.W.; Tandy, S. Dietary phospholipids and intestinal cholesterol absorption. Nutrients 2010, 2, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Volek, J.S.; Volk, B.M.; Gómez, A.L.; Kunces, L.J.; Kupchak, B.R.; Freidenreich, D.J.; Aristizabal, J.C.; Saenz, C.; Dunn-Lewis, C.; Ballard, K.D.; et al. Whey protein supplementation during resistance training augments lean body mass. J. Am. Coll. Nutr. 2013, 32, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Bingham, S.; Cummings, J.H. The use of 4-aminobenzoic acid as a marker to validate the completeness of 24 h urine collections in man. Clin. Sci. 1983, 64, 629–635. [Google Scholar] [CrossRef]
- Harris, J.A.; Benedict, F.G. A Biometric Study of Human Basal Metabolism. Proc. Natl. Acad. Sci. USA 1918, 4, 370–373. [Google Scholar] [CrossRef]
- King, D.E.; Mainous, A.G., 3rd; Lambourne, C.A. Trends in dietary fiber intake in the United States, 1999–2008. J. Acad. Nutr. Diet. 2012, 112, 642–648. [Google Scholar] [CrossRef]
- US Department of Agriculture and US Department of Health and Human Services. Dietary Guidelines for Americans, 2020–2025, 9th ed.; US Department of Agriculture and US Department of Health and Human Services: Washington, DC, USA, 2020. [Google Scholar]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, M.; DeFronzo, R.A. Insulin sensitivity indices obtained from oral glucose tolerance testing: Comparison with the euglycemic insulin clamp. Diabetes Care 1999, 22, 1462–1470. [Google Scholar] [CrossRef]
- Gonzalez-Quintela, A.; Alonso, M.; Campos, J.; Vizcaino, L.; Loidi, L.; Gude, F. Determinants of serum concentrations of lipopolysaccharide-binding protein (LBP) in the adult population: The role of obesity. PLoS ONE 2013, 8, e54600. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Shaw, E.L.; Quarles, W.R.; Sasaki, G.Y.; Dey, P.; Hodges, J.K.; Pokala, A.; Zeng, M.; Bruno, R.S. Daily Inclusion of Resistant Starch-Containing Potatoes in a Dietary Guidelines for Americans Dietary Pattern Does Not Adversely Affect Cardiometabolic Risk or Intestinal Permeability in Adults with Metabolic Syndrome: A Randomized Controlled Trial. Nutrients 2022, 14, 1545. [Google Scholar] [CrossRef] [PubMed]
- Vojdani, A. For the assessment of intestinal permeability, size matters. Altern. Ther. Health Med. 2013, 19, 12–24. [Google Scholar]
- Salminen, S.; Salminen, E. Lactulose, lactic acid bacteria, intestinal microecology and mucosal protection. Scand. J. Gastroenterol. Suppl. 1997, 222, 45–48. [Google Scholar] [CrossRef] [PubMed]
- van Wijck, K.; Verlinden, T.J.; van Eijk, H.M.; Dekker, J.; Buurman, W.A.; Dejong, C.H.; Lenaerts, K. Novel multi-sugar assay for site-specific gastrointestinal permeability analysis: A randomized controlled crossover trial. Clin. Nutr. 2013, 32, 245–251. [Google Scholar] [CrossRef]
- Bakdash, J.Z.; Marusich, L.R. Repeated Measures Correlation. Front. Psychol. 2017, 8, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Yu, Z.; Ye, X.; Zou, S.; Li, H.; Yu, D.; Wu, H.; Chen, Y.; Dore, J.; Clément, K.; et al. A marker of endotoxemia Is associated with obesity and related metabolic disorders in apparently healthy chinese. Diabetes Care 2010, 33, 1925–1932. [Google Scholar] [CrossRef] [Green Version]
- Bowser, S.M.; McMillan, R.P.; Boutagy, N.E.; Tarpey, M.D.; Smithson, A.T.; Osterberg, K.L.; Neilson, A.P.; Davy, B.M.; Davy, K.P.; Hulver, M.W. Serum endotoxin, gut permeability and skeletal muscle metabolic adaptations following a short term high fat diet in humans. Metabolism 2020, 103, 154041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemente-Postigo, M.; Queipo-Ortuño, M.I.; Murri, M.; Boto-Ordoñez, M.; Perez-Martinez, P.; Andres-Lacueva, C.; Cardona, F.; Tinahones, F.J. Endotoxin increase after fat overload is related to postprandial hypertriglyceridemia in morbidly obese patients. J. Lipid Res. 2012, 53, 973–978. [Google Scholar] [CrossRef] [Green Version]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; Gonzalez, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)-mediated gut rpithelial and immune regulation and Its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Millar, C.L.; Jiang, C.; Norris, G.H.; Garcia, C.; Seibel, S.; Anto, L.; Lee, J.Y.; Blesso, C.N. Cow’s milk polar lipids reduce atherogenic lipoprotein cholesterol, modulate gut microbiota and attenuate atherosclerosis development in LDL-receptor knockout mice fed a Western-type diet. J. Nutr. Biochem. 2020, 79, 108351. [Google Scholar] [CrossRef]
- Poppitt, S.D. Cow’s milk and dairy consumption: Is there now consensus for cardiometabolic health? Front. Nutr. 2020, 7, 277. [Google Scholar] [CrossRef]
- Keys, A.; Menotti, A.; Aravanis, C.; Blackburn, H.; Djordevic, B.S.; Buzina, R.; Dontas, A.S.; Fidanza, F.; Karvonen, M.J.; Kimura, N.; et al. The seven countries study: 2,289 deaths in 15 years. Prev. Med. 1984, 13, 141–154. [Google Scholar] [CrossRef]
- Thompson, F.E.; Subar, A.F. Chapter 1—Dietary assessment methodology. In Nutrition in the Prevention and Treatment of Disease, 4th ed.; Coulston, A.M., Boushey, C.J., Ferruzzi, M.G., Delahanty, L.M., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 5–48. [Google Scholar] [CrossRef]
- Louie, J.C.Y.; Flood, V.M.; Hector, D.J.; Rangan, A.M.; Gill, T.P. Dairy consumption and overweight and obesity: A systematic review of prospective cohort studies. Obes. Rev. 2011, 12, e582–e592. [Google Scholar] [CrossRef]
- Chen, M.; Pan, A.; Malik, V.S.; Hu, F.B. Effects of dairy intake on body weight and fat: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2012, 96, 735–747. [Google Scholar] [CrossRef] [Green Version]
- Schwingshackl, L.; Hoffmann, G.; Schwedhelm, C.; Kalle-Uhlmann, T.; Missbach, B.; Knüppel, S.; Boeing, H. Consumption of dairy products in relation to changes in anthropometric variables in adult populations: A systematic review and meta-analysis of cohort studies. PLoS ONE 2016, 11, e0157461. [Google Scholar] [CrossRef]
- Vien, S.; Fabek, H.; Yamagishi, Y.; Lee, Y.T.; Luhovyy, B.L.; Anderson, G.H. Role of single serving form of dairy on satiety and postprandial glycaemia in young and older healthy adults. Appl. Physiol. Nutr. Metab. 2019, 44, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Pei, R.; DiMarco, D.M.; Putt, K.K.; Martin, D.A.; Gu, Q.; Chitchumroonchokchai, C.; White, H.M.; Scarlett, C.O.; Bruno, R.S.; Bolling, B.W. Low-fat yogurt consumption reduces biomarkers of chronic inflammation and inhibits markers of endotoxin exposure in healthy premenopausal women: A randomised controlled trial. Br. J. Nutr. 2017, 118, 1043–1051. [Google Scholar] [CrossRef]
- Bhinder, G.; Allaire, J.M.; Garcia, C.; Lau, J.T.; Chan, J.M.; Ryz, N.R.; Bosman, E.S.; Graef, F.A.; Crowley, S.M.; Celiberto, L.S.; et al. Milk fat globule membrane supplementation in formula modulates the Neonatal Gut microbiome and normalizes intestinal development. Sci. Rep. 2017, 7, 45274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, R.; Du, X.; Brink, L.; Lönnerdal, B. The role of orally ingested milk fat globule membrane on intestinal barrier functions evaluated with a suckling rat pup supplementation model and a human enterocyte model. J. Nutr. Biochem. 2022, 108, 109084. [Google Scholar] [CrossRef]
- Demmer, E.; Van Loan, M.D.; Rivera, N.; Rogers, T.S.; Gertz, E.R.; German, J.B.; Smilowitz, J.T.; Zivkovic, A.M. Addition of a dairy fraction rich in milk fat globule membrane to a high-saturated fat meal reduces the postprandial insulinaemic and inflammatory response in overweight and obese adults. J. Nutr. Sci. 2016, 5, e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadav, H.; Lee, J.H.; Lloyd, J.; Walter, P.; Rane, S.G. Beneficial metabolic effects of a probiotic via butyrate-induced GLP-1 hormone secretion. J. Biol. Chem. 2013, 288, 25088–25097. [Google Scholar] [CrossRef] [Green Version]
- Vors, C.; Joumard-Cubizolles, L.; Lecomte, M.; Combe, E.; Ouchchane, L.; Drai, J.; Raynal, K.; Joffre, F.; Meiller, L.; Le Barz, M.; et al. Milk polar lipids reduce lipid cardiovascular risk factors in overweight postmenopausal women: Towards a gut sphingomyelin-cholesterol interplay. Gut 2020, 69, 487–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira Otto, M.C.; Mozaffarian, D.; Kromhout, D.; Bertoni, A.G.; Sibley, C.T.; Jacobs, D.R., Jr.; Nettleton, J.A. Dietary intake of saturated fat by food source and incident cardiovascular disease: The multi-ethnic study of atherosclerosis. Am. J. Clin. Nutr. 2012, 96, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Yakoob, M.Y.; Shi, P.; Willett, W.C.; Rexrode, K.M.; Campos, H.; Orav, E.J.; Hu, F.B.; Mozaffarian, D. Circulating biomarkers of dairy fat and risk of incident diabetes mellitus among men and women in the United States in two large prospective cohorts. Circulation 2016, 133, 1645–1654. [Google Scholar] [CrossRef] [Green Version]
- Rautiainen, S.; Wang, L.; Lee, I.M.; Manson, J.E.; Buring, J.E.; Sesso, H.D. Dairy consumption in association with weight change and risk of becoming overweight or obese in middle-aged and older women: A prospective cohort study1, 2, 3. Am. J. Clin. Nutr. 2016, 103, 979–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, S.; Moreira, P.; Moreira, C.; Mota, J.; Moreira-Silva, I.; Santos, P.C.; Santos, R. Intake of milk, but not total dairy, yogurt, or cheese, is negatively associated with the clustering of cardiometabolic risk factors in adolescents. Nutr. Res. 2014, 34, 48–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norris, G.H.; Porter, C.M.; Jiang, C.; Millar, C.L.; Blesso, C.N. Dietary sphingomyelin attenuates hepatic steatosis and adipose tissue inflammation in high-fat-diet-induced obese mice. J. Nutr. Biochem. 2017, 40, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Norris, G.H.; Milard, M.; Michalski, M.-C.; Blesso, C.N. Protective properties of milk sphingomyelin against dysfunctional lipid metabolism, gut dysbiosis, and inflammation. J. Nutr. Biochem. 2019, 73, 108224. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | COMP (250 mL) | MEB (250 mL) |
|---|---|---|
| Energy (kcal) | 170 | 162 |
| Total carbohydrates (g) | 14.8 | 13.2 |
| Total protein (g) | 8.4 | 8.4 |
| Total fat (g) | 7.4 | 7.1 |
| Saturated (g) | 5.3 | 4.4 |
| Monounsaturated (g) | 1.8 | 2.1 |
| Polyunsaturated (g) | 0.3 | 0.6 |
| Trans (mg) | 0.0 | 4.6 |
| Total phospholipids (mg) | 770.0 | 772.5 |
| Sphingomyelin (mg) | 0.0 | 125.9 |
| Phosphatidylserine (mg) | 0.0 | 108.7 |
| Phosphatidylcholine (mg) | 185.2 | 171.7 |
| Phosphatidylethanolamine (mg) | 206.9 | 260.2 |
| Phosphatidylinositol (mg) | 175.4 | 63.7 |
| Other phospholipids (mg) 2 | 202.3 | 42.3 |
| C4:0 (mg) | 0.0 | 205.5 |
| C6:0 (mg) | 0.0 | 143.7 |
| C8:0 (mg) | 112.7 | 99.2 |
| C10:0 (mg) | 112.0 | 216.3 |
| C12:0 (mg) | 939.8 | 261.0 |
| C14:0 (mg) | 490.8 | 779.6 |
| C16:0 (mg) | 2537.4 | 2048.6 |
| C18:0 (mg) | 402.6 | 800.4 |
| C20:0 (mg) | 2.5 | 3.2 |
| C14:1 (mg) | 0.00 | 26.1 |
| C16:1 (mg) | 0.72 | 115.2 |
| C18:1 (mg) | 2186.7 | 1596.0 |
| C18:2 (mg) | 650.0 | 223.2 |
| C18:3 (mg) | 3.6 | 35.8 |
| All (n = 24) | Men (n = 11) | Women (n = 13) | p | |
|---|---|---|---|---|
| Age (y) | 37.3 ± 2.1 | 33.6 ± 2.1 | 40.4 ± 3.2 | 0.11 |
| BMI (kg/m2) | 33.4 ± 1.1 | 32.9 ± 1.4 | 33.8 ± 1.7 | 0.70 |
| Waist circumference (cm) | 106.6 ± 2.1 | 108.2 ± 3.2 | 105.2 ± 2.9 | 0.50 |
| SBP (mmHg) | 120.4 ± 2.3 | 124.6 ± 2.7 | 116.9 ± 3.4 | 0.09 |
| DBP (mmHg) | 83.1 ± 2.1 | 87.0 ± 2.7 | 79.9 ± 3.0 | 0.10 |
| Plasma triglyceride (mg/dL) | 130.1 ± 16.0 | 149.8 ± 30.3 | 113.5 ± 14.4 | 0.27 |
| Plasma HDL-C (mg/dL) | 37.2 ± 1.7 | 34.2 ± 2.7 | 39.7 ± 2.1 | 0.12 |
| Plasma glucose (mg/dL) | 101.8 ± 1.8 | 103.7 ± 2.8 | 100.2 ± 2.4 | 0.34 |
| MetS criteria | ||||
| Three risk factors (%) | 79 | 64 | 92 | 0.14 |
| Four risk factors (%) | 21 | 36 | 8 | 0.14 |
| Five risk factors (%) | 0 | 0 | 0 | - |
| Waist circumference (%) | 88 | 73 | 100 | 0.08 |
| HDL-C (%) | 96 | 91 | 100 | 0.46 |
| Glucose (%) | 71 | 73 | 69 | 0.99 |
| Blood pressure (%) | 42 | 64 | 23 | 0.09 |
| Triglyceride (%) | 25 | 36 | 15 | 0.36 |
| COMP | MEB | p1 | |
|---|---|---|---|
| Prescribed energy (kcal) 2 | 2450 ± 64 | 2450 ± 64 | - |
| Consumed energy (kcal) 2 | 2178 ± 77 | 2212 ± 70 | 0.06 |
| Carbohydrate (% kcal) | 54.2 ± 0.31 | 54.3 ± 0.27 | 0.28 |
| Fat (% kcal) | 29.4 ± 0.19 | 29.8 ± 0.17 | 0.70 |
| Protein (% kcal) | 18.6 ± 0.17 | 18.4 ± 0.14 | 0.19 |
| Non-prescribed foods (% kcal) | 2.4 ± 0.006 | 3.2 ± 0.01 | 0.47 |
| Fiber (g) | 16.4 ± 0.29 | 16.9 ± 0.26 | 0.11 |
| HEI-2015 total score 3 | 62.2 ± 0.35 | 62.1 ± 0.36 | 0.83 |
| HEI-2015 adequacy components | |||
| Total fruits | 4.6 ± 0.06 | 4.6 ± 0.06 | 0.73 |
| Whole fruits | 4.6 ± 0.06 | 4.7 ± 0.06 | 0.41 |
| Total vegetables | 3.7 ± 0.01 | 3.7 ± 0 | 0.06 |
| Greens and beans | 0.3 ± 0.02 | 0.3 ± 0.02 | 0.66 |
| Whole grains | 1.6 ± 0.07 | 1.6 ± 0.07 | 0.33 |
| Dairy | 0 | 0 | - |
| Total protein foods | 4.4 ± 0.06 | 4.5 ± 0.05 | 0.67 |
| Seafood and plant proteins | 2.3 ± 0.13 | 2.2 ± 0.13 | 0.85 |
| Healthy fats | 9.8 ± 0.09 | 9.8 ± 0.09 | 0.47 |
| HEI-2015 moderation components | |||
| Refined grains | 3.9 ± 0.16 | 3.6 ± 0.16 | 0.21 |
| Sodium | 10 ± 0.02 | 9.8 ± 0.07 | 0.06 |
| Added sugars | 9.6 ± 0.05 | 9.6 ± 0.04 | 0.73 |
| Saturated fat | 9.9 ± 0.09 | 9.8 ± 0.08 | 0.47 |
| COMP | MEB | ||||||
|---|---|---|---|---|---|---|---|
| Parameter | Day 0 | Day 13 | Day 0 | Day 13 | PTime | PTreatment | PInteraction |
| BMI (kg/m2) | 33.4 ± 1.1 | 33.2 ± 1.1 | 33.7 ± 1.1 | 33.4 ± 1.1 | 0.0013 | 0.05 | 0.80 |
| Waist circumference (cm) | 107.5 ± 2.2 | 107.3 ± 2.2 | 108.0 ± 2.3 | 107.5 ± 2.5 | 0.25 | 0.81 | 0.87 |
| SBP (mmHg) | 119.9 ± 2.8 | 118.3 ± 2.0 | 121.4 ± 3.0 | 119.6 ± 2.6 | 0.64 | 0.65 | 0.76 |
| DBP (mmHg) | 81.8 ± 1.7 | 81.8 ± 1.7 | 84.9 ± 2.2 | 83.2 ± 2.1 | 0.73 | 0.07 | 0.45 |
| Glucose (mg/dL) | 101.2 ± 1.8 | 99.6 ± 2.5 | 101.6 ± 1.7 | 98.1 ± 1.6 | 0.10 | 0.71 | 0.42 |
| Insulin (μIU/mL) | 17.2 ± 9.9 | 14.5 ± 6.0 | 16.4 ± 6.5 | 15.1 ± 6.5 | 0.10 | 0.96 | 0.41 |
| HOMA-IR | 4.2 ± 0.4 | 3.5 ± 0.3 | 4.0 ± 0.3 | 3.6 ± 0.3 | 0.10 | 0.86 | 0.59 |
| GIP (pg/mL) | 101.9 ± 13.6 a | 90.6 ± 13.2 a | 87.3 ± 15.5 b | 67.0 ± 6.9 b | 0.12 | 0.006 | 0.63 |
| GLP-1 (pmol/L) | 7.4 ± 2.4 | 6.7 ± 2.2 | 6.1 ± 1.6 | 6.0 ± 1.6 | 0.05 | 0.24 | 0.13 |
| Triglyceride (mg/dL) | 109.2 ± 8.9 | 103.3 ± 9.9 | 106.0 ± 9.8 | 101.7 ± 8.0 | 0.49 | 0.61 | 0.83 |
| Total cholesterol (mg/dL) | 183.9 ± 8.9 a | 161.8 ± 8.3 b | 186.5 ± 9.8 a | 187.0 ± 8.9 a | 0.09 | 0.0019 | 0.0064 |
| HDL-cholesterol (mg/dL) | 36.2 ± 1.8 a | 34.1 ± 1.4 b | 36.5 ± 1.7 a | 32.3 ± 1.3 b | 0.0004 | 0.23 | 0.07 |
| LDL-cholesterol (mg/dL) 2 | 125.8 ± 7.8 a | 107.0 ± 7.6 b | 128.9 ± 8.7 a | 134.4 ± 8.4 a | 0.23 | 0.002 | 0.003 |
| CD14 (ng/mL) | 1255.9 ± 38.0 | 1172.3 ± 30.8 | 1282.7 ± 42.6 | 1210.8 ± 33.9 | 0.0057 | 0.22 | 0.92 |
| LBP (ng/mL) | 5315.4 ± 275.9 | 5097.4 ± 201.6 | 5647.1 ± 309.7 | 5542.0 ± 346.8 | 0.42 | 0.03 | 0.78 |
| LBP:sCD14 | 4.2 ± 0.2 | 4.4 ± 0.2 | 4.4 ± 0.2 | 4.6 ± 0.2 | 0.27 | 0.10 | 0.94 |
| Endotoxin (EU/mL) | 32.3 ± 2.0 | 30.6 ± 2.1 | 36.3 ± 2.5 | 32.5 ± 2.3 | 0.13 | 0.27 | 0.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pokala, A.; Quarles, W.R.; Ortega-Anaya, J.; Jimenez-Flores, R.; Cao, S.; Zeng, M.; Hodges, J.K.; Bruno, R.S. Milk-Fat-Globule-Membrane-Enriched Dairy Milk Compared with a Soy-Lecithin-Enriched Beverage Did Not Adversely Affect Endotoxemia or Biomarkers of Gut Barrier Function and Cardiometabolic Risk in Adults with Metabolic Syndrome: A Randomized Controlled Crossover Trial. Nutrients 2023, 15, 3259. https://doi.org/10.3390/nu15143259
Pokala A, Quarles WR, Ortega-Anaya J, Jimenez-Flores R, Cao S, Zeng M, Hodges JK, Bruno RS. Milk-Fat-Globule-Membrane-Enriched Dairy Milk Compared with a Soy-Lecithin-Enriched Beverage Did Not Adversely Affect Endotoxemia or Biomarkers of Gut Barrier Function and Cardiometabolic Risk in Adults with Metabolic Syndrome: A Randomized Controlled Crossover Trial. Nutrients. 2023; 15(14):3259. https://doi.org/10.3390/nu15143259
Chicago/Turabian StylePokala, Avinash, William R. Quarles, Joana Ortega-Anaya, Rafael Jimenez-Flores, Sisi Cao, Min Zeng, Joanna K. Hodges, and Richard S. Bruno. 2023. "Milk-Fat-Globule-Membrane-Enriched Dairy Milk Compared with a Soy-Lecithin-Enriched Beverage Did Not Adversely Affect Endotoxemia or Biomarkers of Gut Barrier Function and Cardiometabolic Risk in Adults with Metabolic Syndrome: A Randomized Controlled Crossover Trial" Nutrients 15, no. 14: 3259. https://doi.org/10.3390/nu15143259
APA StylePokala, A., Quarles, W. R., Ortega-Anaya, J., Jimenez-Flores, R., Cao, S., Zeng, M., Hodges, J. K., & Bruno, R. S. (2023). Milk-Fat-Globule-Membrane-Enriched Dairy Milk Compared with a Soy-Lecithin-Enriched Beverage Did Not Adversely Affect Endotoxemia or Biomarkers of Gut Barrier Function and Cardiometabolic Risk in Adults with Metabolic Syndrome: A Randomized Controlled Crossover Trial. Nutrients, 15(14), 3259. https://doi.org/10.3390/nu15143259