1. Introduction
Nutritional strategies to enhance exercise performance and recovery are of current scientific interest to individuals who regularly undertake physical activity, competitive athletes, military workers, as well as the general population. Recent approaches which have gained popularity in an attempt to attenuate exercise-induced muscle damage (EIMD) and oxidative stress include the supplementation of naturally occurring phytochemicals (i.e., polyphenols) from sources such as pomegranate, cocoa, or cherries [
1,
2,
3]. The average adult consumption of polyphenols is suggested to be ~1 g·d
−1 [
4], with primary sources from fruits, vegetables, beverages such as tea and coffee, wine, and chocolate [
5]. With antioxidant properties [
6], nutritional polyphenols may act as radical scavengers and metal chelators, regulating metabolism, body mass, chronic disease, and cell proliferation [
7]. Free radicals and reactive oxygen and nitrogen species (RONS) are the primary oxidizing agents produced in cellular biochemical reactions for aerobic energy production [
5]. Aerobic exercise is characterized by increased total energy expenditure [
8], where the availability of endogenous substrates and aerobic metabolism are crucial for overall performance [
9,
10]. The increased oxygen (O
2) demand by skeletal muscles during exercise results in greater free radical production and an increase in RONS [
11]. Whilst viewed as detrimental to the cell for many years, recent evidence shows that RONS are crucial physiological activators and regulators of various intracellular signaling pathways in response to stress, enhancing defense, improving cell adaptation, and upregulating the expression of endogenous antioxidant enzymes [
12,
13].
Furthermore, exercise adaptations are dependent, at least partially, on an acute oxidative stress response. When exercise intensity is matched, individuals expressing lower levels of RONS have demonstrated inferior training adaptations compared to those with moderate or higher levels of RONS [
14]. However, during excessive and demanding exercise, an imbalance between RONS and endogenous antioxidants induces oxidative damage, potentially impacting at a mitochondrial or DNA level [
15], reducing vasodilatory capacity [
16] and contractile force within the muscle through impaired calcium sensitivity [
17]. This can have inferences for repetitive training sessions or longer-term adaptations and may, therefore, impair exercise performance and/or the recovery process. In sports where arterial blood flow and maximum cardiac output are determinants of performance (i.e., endurance and team-based sports), acute ingestion (<3 h before competition) or chronic supplementation of polyphenols (~7-days) could improve time to exhaustion at 70% maximum oxygen uptake (
O
2max) by +9.7% [
18] and intermittent high-intensity running distance by +10% [
19].
The mechanisms by which polyphenols may facilitate ergogenic effects reportedly occur via nitric oxide synthase production [
20] as well as the activation of sirtuin 1 (SIRT1) [
21,
22]. SIRT1 deacetylates several transcription factors such as forkhead (FOXO) proteins and peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) [
23]. This can facilitate mitochondrial biogenesis, endothelial function, cell proliferation and differentiation, metabolic efficiency, resistance to stress, and improve inflammatory and immune function [
24,
25,
26]. The supplementation of phenolic compounds, and gut-derived metabolites, may therefore provide adjunct or indirect ergogenic effects on physical performance by way of potentially reducing the O
2 cost of exercise (i.e., economy), enhancing
O
2max or exercise tolerance, and/or improving substrate utilization efficiency. Previous findings have highlighted that polyphenol nutrients (e.g., resveratrol) may support mitochondrial function [
27] and may therefore modulate essential biological functions (including thermogenesis, mitochondrial biogenesis and adenosine triphosphate production) [
28]. These functions are pivotal for trained, recreationally active and untrained exercising individuals, ensuring that substrate supply kinetics and waste product removal match the requirements of the specific exercise bout [
29].
Furthermore, it could be inferred that due to the anti-inflammatory and immuno-modulatory effects of phenolic compounds, an increase in polyphenol consumption (from food or supplementation) may be pertinent to exercise recovery. A reduction in physiological stressors that negatively impact exercise training [
30] may support fast and slow phases of recovery, influencing performance in both prolonged or repeated bouts of exercise. Evidence for enhanced functional recovery from foods/supplements high in polyphenol compounds (e.g., Montmorency cherries > 5-days) have been exhibited in both trained and untrained individuals in a multitude of general exercise settings [
31,
32,
33]. However, further research is warranted to investigate other polyphenols or novel food products, to assess markers of exercise recovery and identify the potential impact of phenolic compounds in specific exercise settings.
This is the first study to undertake an investigation into a commercially available polyphenol-rich olive fruit water, OliPhenolia
® (OliP), which has not been assessed in an exercise domain. Originating during the olive picking season, this polyphenol-rich drink is extracted via concentration, reverse osmosis, and ceramic membrane technology at the aqueous part of the olive fruit. Whilst OliP contains a variety of phenolic compounds, it is particularly rich in hydroxytyrosol (HT). Abundant in olives in the form of pure HT, HT glycosides and oleuropein, HT is an effective antioxidant, with studies highlighting protection against oxidative stress in vascular tissue [
34,
35], low-density lipoprotein oxidation [
36,
37,
38], and a reduction in oxidative damage in intestinal epithelial cells [
39], hepatocytes, and erythrocytes [
40]. However, OliP has yet to be considered within an exercise and/or recovery domain and thus, requires investigation. Therefore, this study investigated the effect of OliP on submaximal and exhaustive exercise, as well as respiratory markers of acute recovery, in recreationally active volunteers. Understanding the efficacy of OliP may inform future nutritional strategies pertinent to exercise training and recovery.
4. Discussion
To the authors’ knowledge, this is the first study to undertake research focusing on OliP in an exercise domain and aligns with concurrent research pertinent to olive-derived phytonutrients [
45]. The key findings from this study demonstrate that 16 consecutive days consumption of OliP resulted in positive effects on several key markers of running performance. Of particular interest, OliP consumption significantly improved respiratory parameters at the onset of exercise within condition (i.e., τ), and oxygen consumption and running economy at LT1 (particularly when expressed as relative change in comparison to PL). Whilst respiratory parameters at LT2 were largely unaffected by OliP, perceived exertion was improved with the phytocomplex beverage. Acute recovery (T50%) following incremental exercise was also notably improved with OliP. Whilst maximal effort and TTE measures were not different between OliP and PL, an elevated
CO
2max was reported for PL only. Furthermore, it was noted that both groups improved TTE following the intervention. Importantly, no adverse effects were reported throughout the intervention.
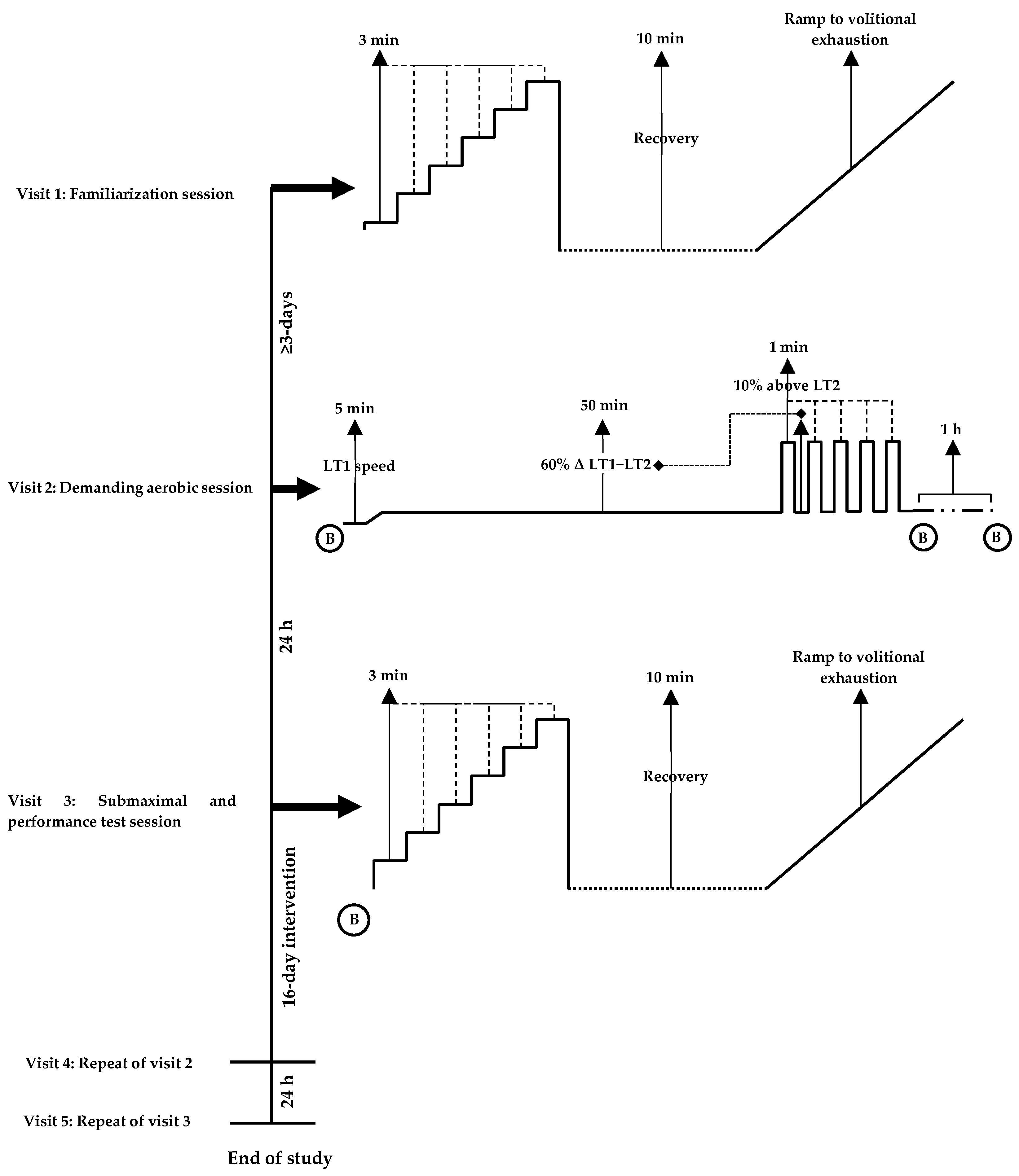
Regarding methodological approaches to the demanding aerobic session, steady-state moderate intensity exercise (60–70%
O
2max) for 30–60 min followed by arduous (90%
O
2max) [
57] or performance efforts [
58] have been shown to provoke a heightened oxidative stress response and elicit peripheral fatigue. Accordingly, the demanding aerobic sessions employed in the current study resulted in an intensity of ~75%
O
2max, with no differences within or between groups. It can therefore be assumed that an equal degree of physiological strain was achieved between cohorts prior to the main performance tests. As dietary and exercise habits were maintained across the intervention, it is feasible that physiological adaptations observed, may therefore be partly attributed to the phenolic compounds within OliP. As a naturally derived phytocomplex, OliP is notably rich in HT, which is a key polyphenol of interest and may support endogenous antioxidant mechanisms pertinent to mitochondrial respiratory capacity and/or efficiency, such as upregulation of PGC-1α [
28,
59,
60,
61,
62].
Consumption of OliP may therefore be of relevance to individuals who engage in regular aerobic exercise, considering the negligible dietary HT content in both the pre-visit control period and habitual diet assessments for both cohorts. Plasma HT concentrations were not detected at baseline (pre-supplementation), or post PL, but were significantly elevated in response to the OliP intervention. Therefore, any impact on aerobic exercise may be associated with increased systemic HT concentrations, or gut-derived metabolites. At present, however, there is a paucity of scientific research surrounding HT and exercise performance. Additionally, there does not appear to be any existing research evidencing the effects of HT on aerobic running performance in humans. Plant-based polyphenols have peaked scientific interest in recent years [
29,
63], in particular HT, due to its potential to impact multiple physiological pathways. In an exercise domain, recent animal studies have demonstrated the ability of HT to enhance endurance capacity [
59], prevent exercise induced fatigue, muscle damage and immunosuppression [
64,
65] and improve mitochondrial function in both trained and sedentary rodents [
65]. However, these findings need to be corroborated in human models as well as within an exercise domain to ascertain the efficacy of HT-rich supplements.
It is also important to outline the current debate surrounding the efficacy of antioxidant and polyphenol supplementation as an exercise or training aid. Adaptations from exercise are dependent, at least partially, on individual oxidative stress responses [
66]. One perspective highlights the potential inhibition of natural training adaptations through limiting the upregulation of endogenous antioxidant enzymes, and therefore diminishing the hormetic response to moderate exercise [
13]. However, the counterargument highlights that the subsequent reduction in oxidative stress following antioxidant and/or polyphenol supplementation may positively influence recovery kinetics, development in contractile force, calcium handling, and therefore the ability to exercise and/or recover more ‘economically’. This may facilitate adaptations to exercise training and/or athletic performance [
17].
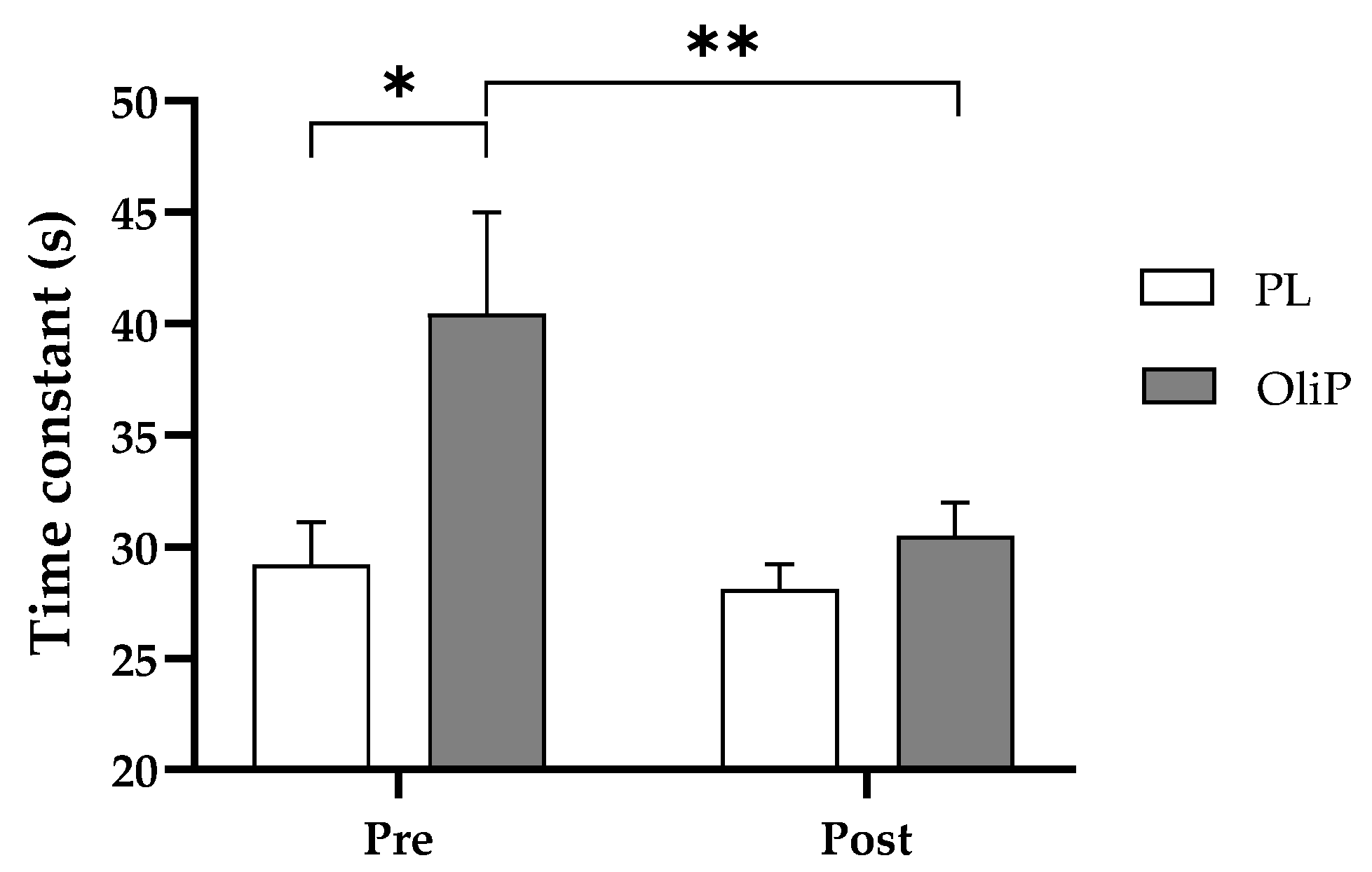
Findings from this study demonstrated a ~17% improvement in τ at the onset of exercise for OliP. τ reflects the speed at which the steady-state is achieved [
53], and in turn the size of the O
2 deficit [
67]. However, these results were only significant within condition and should therefore be interpreted with caution. In addition, it was noted that non-significant differences were observed between conditions prior to the nutritional intervention based on random participant allocation, which may in part impact the observed findings. Contrary to these findings, Breese et al. [
68] reported no differences in
O
2 phase II time constant, from unloaded to moderate exercise after 6-days supplementation with beetroot juice (BTJ; ~8 mmol nitrate (NO
3−)). Although mitochondrial respiratory capacity was not assessed, it is known that the speed of the O
2 uptake response during the onset of moderate exercise intensity is associated with the respiratory capacity of mitochondrial complex II and the capacity of the mitochondrial electron transport system [
69,
70]. As HT has been shown to improve the expression of mitochondrial complex I/II/IV, this is of particular interest in an exercise domain as complex I is recognized to be the primary complex for the electron transport chain [
71]. Moreover, HT has been reported to promote the congregation of complex I (CI) into supercomplexes (SCs) [
65], therefore decreasing the diffusion distance for transfer of electrons between complexes, and improving the efficiency of the mitochondrial electron transfer between complexes [
59,
72]. More research is required to ascertain the above stated mechanisms in humans, particularly in relation to OliP consumption.
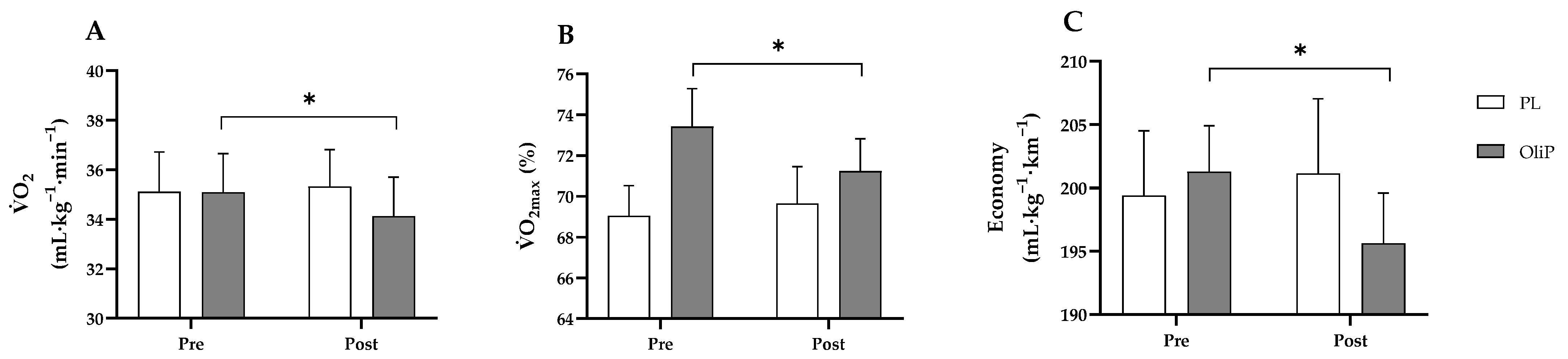
Consumption of OliP in the dose provided also resulted a significant decrease (−2.7%) in
O
2 consumption at LT1 compared with PL. This aligns with existing research into both high [
73]- and low [
74]-dose BTJ supplementation whereby a ~5% reduction
O
2 consumption was reported with no changes in
E, RER or HR [
74]. This modest change could be partially attributed to the increase in mitochondrial function and increased expression of PGC-1α following supplementation of OliP. In vitro, HT administration has been shown to upregulate nuclear respiratory factors 1 and 2, mitochondrial transcription factor A, and peroxisome proliferator active receptor γ (PPAR γ) in response to increased phosphorylation of adenosine monophosphate kinase (AMPK) [
61]. The role HT may play in enhancing mitochondrial respiratory capacity could also provide a rationale for the reduced oxygen consumption observed during sub-maximal exercise at low to moderate intensities (LT1). In vitro, HT has been shown to improve mitochondrial biogenesis, O
2 and fatty acid utilization in adipocyte cells [
61,
75,
76]. Although not measured in this study, this may support the proposed benefits of OliP in a submaximal exercise domain, however, more research is required in humans to confirm such mechanisms. It is also viable that other phenolic compounds [
38] (i.e., oleuropein aglycone) and HT derivatives (i.e., HT glucosides) found in OliP may also support antioxidant pathways that may influence aerobic performance [
59,
77]. Indeed, olive-derived phenolic compounds are not entirely absorbed during digestion and are extensively transformed into different metabolites by the gut microbiota [
78]. For instance, whilst oleuropein transformation by gut bacteria can increase HT yield [
79], HT is further transformed into homovanillin derivatives [
80,
81] and glutathione conjugates [
79] which may have pertinent antioxidant properties [
82]. These metabolites may exert further physiological effects [
83] potentially explaining findings from the current study. Furthermore, HT-derived metabolite variability and quantity are also dependent on the phenolic composition of the product consumed [
81]. Such complexities should also be considered when determining the physiological impact of combined polyphenols.
A relevant parameter of aerobic performance is the efficiency of movement, i.e., exercise economy [
84]. This reflects the amount of O
2 required to generate a constant submaximal running speed and therefore, is directly associated with the efficiency of aerobic fuel metabolism and the sparing of glycogen reserves [
85]. Mitochondria are crucial for aerobic energy generation in exercise [
86]. Improvement in mitochondrial respiratory capacity and functional efficiency following HT supplementation in animal studies has been established [
87] and is associated with the constitution of supramolecular entities, the mitochondrial SCs, including respiratory complex I, III, and IV [
88]. Administration of HT for 10-weeks in rodents (20 vs. 300 mg·kg
−1·d
−1) and exercise (up to 65 min a day at 75% of maximal velocity) compared to exercise alone improved mitochondrial function and antioxidant capacity induced by exercise [
65]. However, when the HT dose was increased to 300 mg·kg
−1·d
−1, pro-oxidant effects were evident [
65], which appeared to negatively influence SCs assembly, aligning with existing published literature [
64]. Collectively, these results indicate that whilst exercise induces the formation of mitochondrial SCs [
89], low-dose HT consumption may support or enhance this process [
65] whilst a high dose of HT may provoke pro-oxidant mechanisms, disrupting the mitochondria and potentially limiting or diminishing SC adaptation [
64]. In the current study, a relatively low HT dose was employed as part of the olive-derived phytocomplex (~0.8 mg·kg
−1·d
−1) in healthy volunteers. Whilst mitochondrial function was not directly assessed, it is feasible that HT and related gut-derived phenolic metabolites may have supported SC assembly and facilitated improved oxygen cost responses observed at the onset of exercise and during low to moderate exercise (LT1). Furthermore, the low HT dose employed in the current study may also explain why exercise performance (TTE) was not significantly different between cohorts in line with previous research [
64].
Despite OliP presenting no significant impact on respiratory mechanisms at LT2, a poignant finding was the observed significant reduction in RPE at this intensity. Mechanisms for this are unclear, however it is feasible that there may be a link to a reduction in brain oxygenation that is present during intensive exercise and directly associated with an increase of fatigue (subjectively quantified as perceived exertion) [
90]. Alternatively, mechanisms potentially occurring at a mitochondrial level and the effect upon SCI and SCII, may indicate that beneficial responses to OliP are more likely to be present at lower intensities only. Further research is required to accurately ascertain potential mechanisms involved in subjective measures associated with exercise.
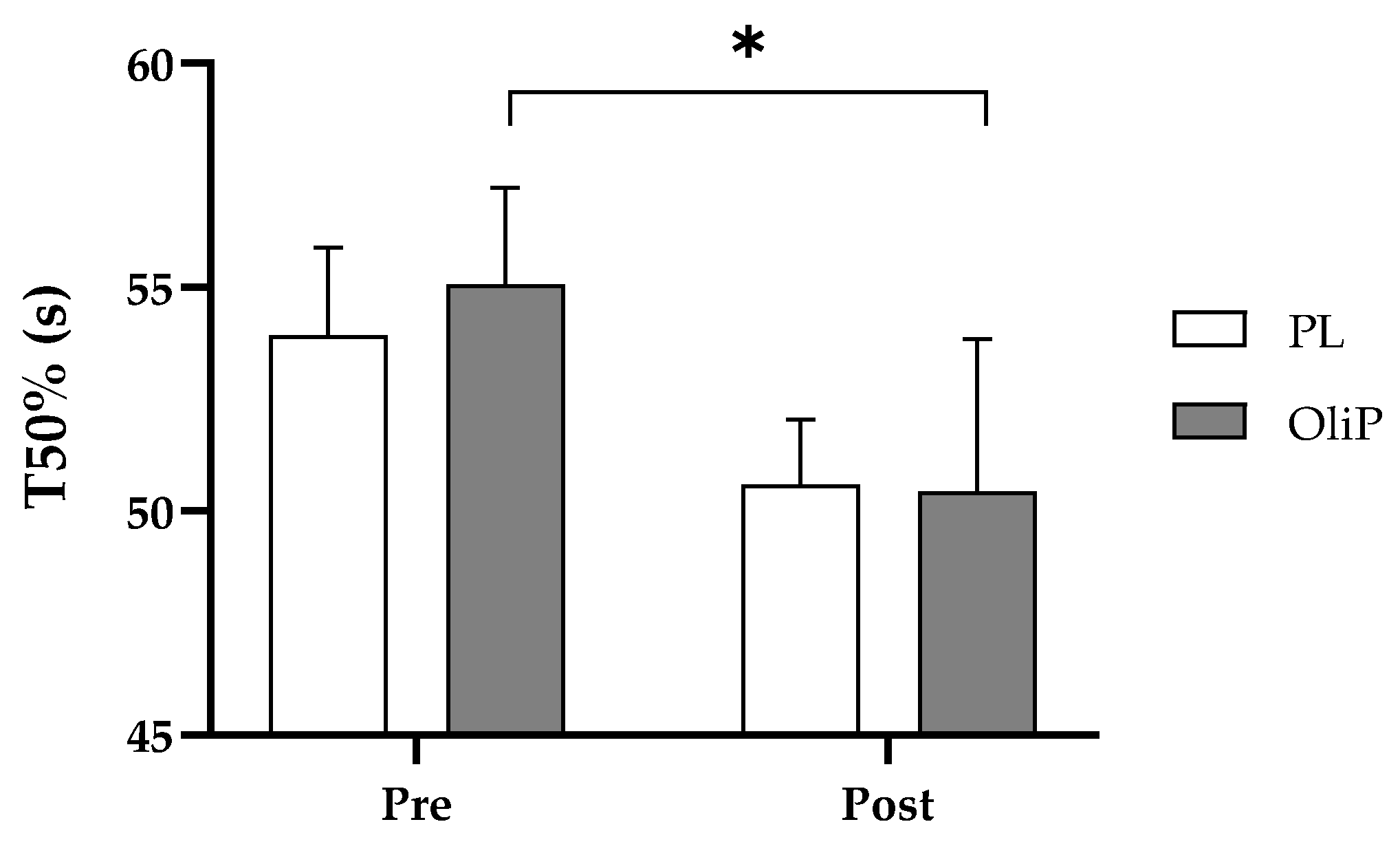
Similarly, recovery was largely unaffected based upon off-kinetic modeling; however, current findings did present a −9.4% decrease in T50% for OliP compared with a −5.6% decrease in PL, during the initial recovery period from sub-maximal exercise. This in itself warrants further investigation considering that previous findings utilizing a similar exercise intensity (70% maximum aerobic power) did not find a benefit to
O
2 half-recovery time following the supplementation of mixed polyphenols (250 mg Vinitrox
™ for 7-days [
19]). In the current study it is feasible that the HT content in OliP (and related gut-dervied metabolites) may be influencing recovery indirectly, and may therefore have applications following repeated bouts of exercise. However, results should be interpreted with caution and further research should be undertaken to corroborate findings. Finally, although improvements in TTE were evident in both groups (+4.1% OliP and +5.8% PL), the overall change in exercise performance was not different between OliP and PL. In the current study, exercise performance was based upon physical tolerance to sustained near-maximal exercise. Based upon findings at LT1 intensity, it could be prudent to assess whether OliP is more effective when determining performance employing other measures such as an extended time trial (i.e., 5 km run time) or total work completed in a fixed time period as opposed to an acute near-maximal TTE bout.
Study Limitations and Future Directions
It is important to note that there were several limitations to the current study. Firstly, improvements were found in specific, but not all parameters assessed. As example, change in time constant at the onset of exercise was noted within-group only for OliP and therefore should be interpreted with caution. Likewise, during acute recovery, whilst improvements were observed for T50%, other parameters using respiratory off-kinetics were not deemed significant, and again results should be interpreted carefully. However, where main interaction effects were identified (including relative changes in oxygen consumption and running economy at LT1 compared with PL) important adaptations following the inclusion of dietary OliP may be evident. It should, however, be noted that differences were not observed post-intervention between conditions which should be taken into consideration.
Although improvements were observed particularly at LT1 with healthy, recreationally active volunteers, we did not specifically distinguish whether such effects were pertinent to gender, training status, or the type/intensity of habitual exercise. Further research may therefore be relevant to determine the potential applications of OliP in various cohorts. Additionally, the protocol used in the current study was designed to standardize the demanding aerobic run prior to the following day exercise performance session for all participants [
45]. It is important to recognize the translation of controlled laboratory findings to real-world exercise applications [
91], and future research should investigate the adjunct use of OliP in applied and field-based settings (e.g., single exercise sessions, events that require repeated bouts, or multi-day events). It should also be noted that existing literature has outlined the potential variability in polyphenol products [
92]. Whilst 16-days of OliP consumption (Batch 14) positively influenced aerobic exercise parameters and acute recovery, results may differ between batches and additional investigation is needed to corroborate current findings. Indeed, as previously noted, the intestinal microbiota plays an important role influencing gut-derived phenolic metabolites, which are additionally dependent on the phenolic composition of dietary products.
As this was the first study to assess the use of OliP in an exercise domain, a parallel cohort design was employed to ascertain the influence of a single course of the phytocomplex (16 consecutive days) whilst minimizing potential for longer term training effects. Further research should investigate whether time course (>16-days), dose-response (>56 mL·d
−1) and/or dose-frequency (>2 serves·d
−1) can influence sustained exercise training adaptations or accumulated recovery, i.e., during marathon training. Additional exploration into alternate recovery periods (i.e., respiratory measures up to 1 h post exercise, and inflammatory or muscle soreness measures 1, 12, 24, and 48 h+ following exhaustive exercise) is also warranted. Finally, based upon current findings, including effects of OliP on exercise-induced oxidative stress presented elsewhere [
45], it would be beneficial to assess the potential impact of this olive-derived phytocomplex on inflammatory markers associated with EIMD (particularly within other populations, e.g., trained athletes), or within clinical applications where functional movement may be impacted (e.g., arthritis, fibromyalgia).


 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}