
Bioactive Molecules from Marine Diatoms and Their Value for the Nutraceutical Industry






Abstract
:1. Introduction
1.1. Nutraceuticals from the Sea
1.2. Bioactive Compounds from Microalgae
2. Primary Metabolites
2.1. Proteins and Peptides

{kind=link}
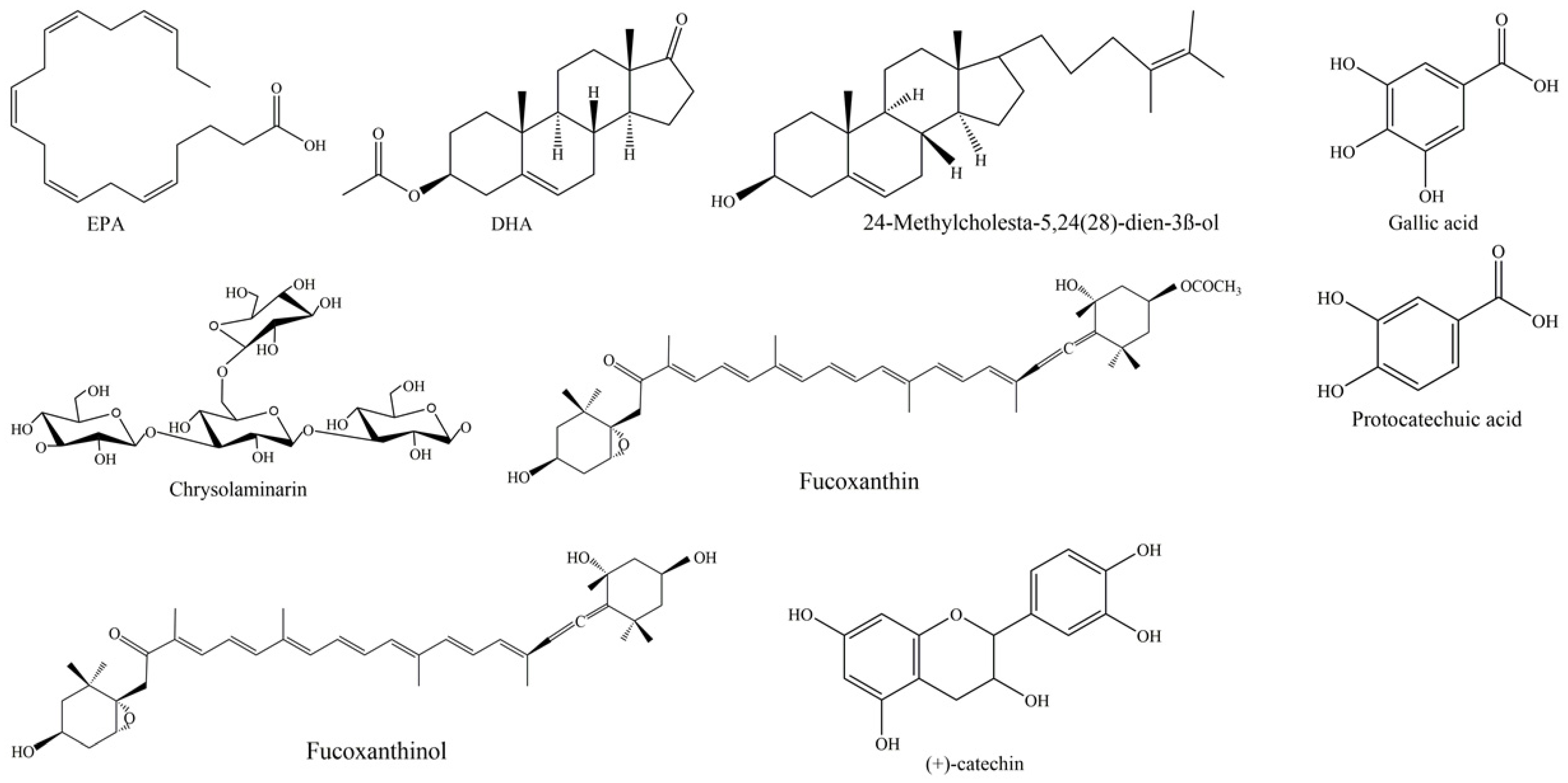{kind=link}
{kind=link}
| Diatom Species | Compound | Biological Activities | References | |
|---|---|---|---|---|
| Proteins/Peptides | ||||
| B. malleus, Bellerochea, Nitzschia | Protein hydrolysates | ACE-inhibitory and antioxidant | [59] | |
| One isolate of B. malleus, four isolates of the genus Bellerochea and one unidentified species of the genus Nitzschia | Papain hydrolysates | ACE-inhibitory and antioxidant | [56] | |
| Navicula sp. | Val-Glu-Val-Leu-Pro-Pro-Ala-Glu-Leu; Pro-Gly-Trp-Asn-Gln-Trp-Phe-Leu | Antioxidant | [54] | |
| Fatty acids | ||||
| P. borealis | EPA-rich biomass | Decreasing cholesterol and triacylglycerol and antioxidant | [60] | |
| Anomoenois sp. and Rhopalodia sp. | Fatty acids | ACE-inhibitory and antioxidant | [61] | |
| P. tricornutum | EPA-rich biomass | Anti-aging (clinical trial) | [62] | |
| Carotenoids | ||||
| N. laevis | Fucoxanthinol | Anti-inflammatory, neuroprotective | [55] | |
| P. tricornutum | Fucoxanthin | Antiproliferative Antioxidant | [59] | |
| P. tricornutum | Fucoxanthin | Anti-inflammatory | [63] | |
| Polysaccharides | ||||
| D. geminata | Crude polysaccharides | Antioxidant and anti-inflammatory | [64] | |
| Halamphora sp. | Sulphated polysaccharide PK3 | Immunostimulant (enhanced phagocytosis of macrophages) | [65] | |
| H. ostrearia | Water soluble fraction | Antiviral (HSV-1) | [66] | |
| N. directa | Naviculan | Antiviral (HSV1 and 2, IFV-A and HIV) | [67] | |
| Navicula sp. | Sulphated polysaccharides | Antioxidant | [68] | |
| O. aurita | Crysolaminarin | Antioxidant | [69] | |
| P. tricornutum | Crude polysaccharides | Anti-inflammatory and immunostimulant | [70] | |
| P. tricornutum | Chrysolaminarin-rich extract | Antioxidant and immunostimulant | [71] | |
| P. tricornutum | P. tricornutum chrysolaminarin-rich biomass | Positive effects on gut microbiota | [72] | |
| S. acus | Chrysolaminarin compound | Cytotoxic vs colon cancer cells | [73] | |
| Polyphenols | ||||
| N. palea | Phenolic extract | Antioxidant Anti-hemolytic Anti-inflammatory (in vitro and in vivo models) | [40] | |
| N. laevis | Phenolic extract | Antioxidant | [74] | |
| P. tricornutum | Gallic acid, protocatecuic acid, catechin, vanillic acid, epicatechin, syringic acid, chlorogenic acid, gentisic acid, caffeic acid, coumaric acid, ferulic acid, rutin, myricetin and quercetin | Antioxidant | [75] | |
| S. marinoi | Phenolic extract | Antioxidant Photoprotective | [76] |
2.2. Fatty Acids
2.3. Sterols
2.4. Polysaccharides
3. Secondary Metabolites
3.1. Carotenoids
3.2. Polyphenols
4. Innovative Cultivation System and Extraction Methods to Meet the Challenging Demands of Nutraceutical Industries
4.1. Culture Systems
4.2. Methods of Extraction of the Active Compounds
5. Conclusions and Future Perspectives and Challenges of Diatoms as Nutraceuticals
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrow, C.; Shahidi, F. Marine Nutraceuticals and Functional Foods; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Andlauer, W.; Furst, P. Nutraceuticals: A piece of history, present status and outlook. Food Res. Int. 2002, 35, 171–176. [Google Scholar] [CrossRef]
- Zeisel, S.H. Regulation of “nutraceuticals”. Science 1999, 285, 1853–1855. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Saklani, S.; Kumar, P.; Kim, B.; Coutinho, H.D.M. Nutraceuticals: Pharmacologically active potent dietary supplements. Biomed Res. Int. 2022, 2022, 2051017. [Google Scholar] [CrossRef] [PubMed]
- Brower, V. Nutraceuticals: Poised for a healthy slice of the healthcare market? Nat. Biotechnol. 1998, 16, 728–731. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Han, Z.; Granato, D. Polyphenols in Foods: Classification, Methods of Identification, and Nutritional Aspects in Human Health, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2021; Volume 98, ISBN 9780128225066. [Google Scholar]
- Jaramillo-Madrid, A.C.; Ashworth, J.; Fabris, M.; Ralph, P.J. Phytosterol biosynthesis and production by diatoms (Bacillariophyceae). Phytochemistry 2019, 163, 46–57. [Google Scholar] [CrossRef]
- Kim, S.-K.; Wijesekara, I. Development and biological activities of marine-derived bioactive peptides: A review. J. Funct. Foods 2010, 2, 1–9. [Google Scholar] [CrossRef]
- Wijesekara, I.; Yoon, N.Y.; Kim, S.K. Phlorotannins from Ecklonia cava (Phaeophyceae): Biological activities and potential health benefits. BioFactors 2010, 36, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Wijesekara, I.; Pangestuti, R.; Kim, S.K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar] [CrossRef]
- Ngo, D.H.; Wijesekara, I.; Vo, T.S.; Van Ta, Q.; Kim, S.K. Marine food-derived functional ingredients as potential antioxidants in the food industry: An overview. Food Res. Int. 2011, 44, 523–529. [Google Scholar] [CrossRef]
- Kratzer, R.; Murkovic, M. Food ingredients and nutraceuticals from microalgae: Main product classes and biotechnological production. Foods 2021, 10, 1626. [Google Scholar] [CrossRef]
- Tsvetanova, F.; Yankov, D. Bioactive compounds from red microalgae with therapeutic and nutritional value. Microorganisms 2022, 10, 2290. [Google Scholar] [CrossRef] [PubMed]
- Darwazeh, A.M.G.; Lamey, P.J.; Samaranayake, L.P.; Macfarlane, T.W.; Fisher, B.M.; Macrury, S.M.; Maccuish, A.C. The relationship between colonisation, secretor status and in-vitro adhesion of Candida albicans to buccal epithelial cells from diabetics. J. Med. Microbiol. 1990, 33, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S.; Sasikala, S.L.; Aldous, V.H.J. Isolation and identification of MDR-Mycobacterium tuberculosis and screening of partially characterised antimycobacterial compounds from chosen marine micro algae. Asian Pac. J. Trop. Med. 2010, 3, 655–661. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Yeon, S.L.; Sang, H.J.; Sam, S.K.; Kuk, H.S. Anti-oxidant activities of fucosterol from the marine algae Pelvetia siliquosa. Arch. Pharm. Res. 2003, 26, 719–722. [Google Scholar] [CrossRef] [PubMed]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Kristine, W.; Sigurður, B. Developing diatoms for value-added products: Challenges and opportunities. New Biotechnol. 2015, 32, 547–551. [Google Scholar] [CrossRef]
- Ferrazzano, G.F.; Papa, C.; Pollio, A.; Ingenito, A.; Sangianantoni, G.; Cantile, T. Cyanobacteria and microalgae as sources of functional foods to improve human general and oral health. Molecules 2020, 25, 5164. [Google Scholar] [CrossRef]
- Buono, S.; Langellotti, A.L.; Martello, A.; Rinna, F.; Fogliano, V. Functional ingredients from microalgae. Food Funct. 2014, 5, 1669–1685. [Google Scholar] [CrossRef]
- Gouveia, L.; Raymundo, A.; Batista, A.P.; Sousa, I.; Empis, J. Chlorella vulgaris and Haematococcus pluvialis biomass as colouring and antioxidant in food emulsions. Eur. Food Res. Technol. 2006, 222, 362–367. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Liu, B.; Wei, H.; Cheng, K.W.; Chen, F. Extract of the microalga Nitzschia laevis prevents high-fat-diet-induced obesity in mice by modulating the composition of gut microbiota. Mol. Nutr. Food Res. 2019, 63, 1800808. [Google Scholar] [CrossRef]
- Tan, C.K.; Johns, M.R. Screening of diatoms for heterotrophic eicosapentaenoic acid production. J. Appl. Phycol. 1996, 8, 59–64. [Google Scholar] [CrossRef]
- Raniello, R.; Iannicelli, M.M.; Nappo, M.; Avila, C.; Zupo, V. Production of Cocconeis neothumensis (Bacillariophyceae) biomass in batch cultures and bioreactors for biotechnological applications: Light and nutrient requirements. J. Appl. Phycol. 2007, 19, 383–391. [Google Scholar] [CrossRef]
- Zupo, V.; Patalano, C.; Messina, P. Culture conditions influence the growth dynamics and the production of Cocconeis scutellum (bacillariophyta). J. Phycol. 2011, 47, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Yool, A.; Tyrrell, T. Role of diatoms in regulating the ocean’s silicon cycle. Glob. Biogeochem. Cycles 2003, 17, 1–21. [Google Scholar] [CrossRef]
- Kociolek, J.P. Diatoms: Unique eukaryotic extremophiles providing insights into planetary change. Instrum. Methods Mission. Astrobiol. 2007, 6694, 242–256. [Google Scholar] [CrossRef]
- Durkin, C.A.; Van Mooy, B.A.S.; Dyhrman, S.T.; Buesseler, K.O. Sinking phytoplankton associated with carbon flux in the Atlantic Ocean. Limnol. Oceanogr. 2016, 61, 1172–1187. [Google Scholar] [CrossRef] [Green Version]
- Maibam, C.; Fink, P.; Romano, G.; Buia, M.C.; Butera, E.; Zupo, V. Centropages typicus (Crustacea, Copepoda) reacts to volatile compounds produced by planktonic algae. Mar. Ecol. 2015, 36, 819–834. [Google Scholar] [CrossRef]
- Zupo, V.; Messina, P.; Buttino, I.; Sagi, A.; Avila, C.; Nappo, M.; Bastida, J.; Codina, C.; Zupo, S. Do benthic and planktonic diatoms produce equivalent effects in crustaceans? Mar. Freshw. Behav. Physiol. 2007, 40, 169–181. [Google Scholar] [CrossRef]
- Maibam, C.; Romano, G.; Fink, P.; Buia, M.C.; Scipione, M.B.; Patti, F.P.; Lorenti, M.; Zupo, V. Dual role of wound-activated compounds produced by diatoms, as both allelochemicals and infochemicals for benthich invertebrates. In Proceedings of the ASLO Summer Meeting, Otsu, Japan, 8–13 July 2012. [Google Scholar]
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Russo, G.L.; Buttino, I.; Mazzarella, G.; Laablr, M.; Cabrini, M.; et al. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar] [CrossRef]
- Jüttner, F.; Messina, P.; Patalano, C.; Zupo, V. Odour compounds of the diatom Cocconeis scutellum: Effects on benthic herbivores living on Posidonia oceanica. Mar. Ecol. Prog. Ser. 2010, 400, 63–73. [Google Scholar] [CrossRef]
- Welker, C.; Nichetto, P. The influence of mucous aggregates on the microphytobenthic community in the Northern Adriatic Sea. Mar. Ecol. 1996, 17, 473–489. [Google Scholar] [CrossRef]
- Maibam, C.; Fink, P.; Romano, G.; Buia, M.C.; Gambi, M.C.; Scipione, M.B.; Patti, F.P.; Lorenti, M.; Butera, E.; Zupo, V. Relevance of wound-activated compounds produced by diatoms as toxins and infochemicals for benthic invertebrates. Mar. Biol. 2014, 161, 1639–1652. [Google Scholar] [CrossRef]
- Zupo, V.; Maibam, C.; Buia, M.C.; Gambi, M.C.; Patti, F.P.; Scipione, M.B.; Lorenti, M.; Fink, P. Chemoreception of the seagrass Posidonia Oceanica by benthic invertebrates is altered by seawater acidification. J. Chem. Ecol. 2015, 41, 766–779. [Google Scholar] [CrossRef] [PubMed]
- Nappo, M.; Berkov, S.; Massucco, C.; Di Maria, V.; Bastida, J.; Codina, C.; Avila, C.; Messina, P.; Zupo, V.; Zupo, S. Apoptotic activity of the marine diatom Cocconeis scutellum and eicosapentaenoic acid in BT20 cells. Pharm. Biol. 2012, 50, 529–535. [Google Scholar] [CrossRef]
- Stock, W.; Blommaert, L.; De Troch, M.; Mangelinckx, S.; Willems, A.; Vyverman, W.; Sabbe, K. Host specificity in diatom-bacteria interactions alleviates antagonistic effects. FEMS Microbiol. Ecol. 2019, 95, fiz171. [Google Scholar] [CrossRef]
- Sudarshan, B.L.; Maheshwar, P.K.; Priya, P.S.; Sanjay, K.R. Volatile and phenolic compounds in freshwater diatom Nitzschia palea as a potential oxidative damage protective and anti-inflammatory source. Pharmacogn. Mag. 2019, 15, 228–236. [Google Scholar] [CrossRef]
- Lakshmegowda, S.B.; Rajesh, S.K.; Kandikattu, H.K.; Nallamuthu, I.; Khanum, F. In Vitro and in vivo studies on hexane fraction of Nitzschia palea, a freshwater diatom for oxidative damage protective and anti-inflammatory response. Rev. Bras. Farmacogn. 2020, 30, 189–201. [Google Scholar] [CrossRef]
- Jeong, K.S.; Jeong, K.Y.; Hong, Y.S.; Kim, D.K.; Oh, H.J.; Chang, K.H. Application of nuclear magnetic resonance for analyzing metabolic characteristics of winter diatom blooms. J. Plankton Res. 2020, 42, 31–39. [Google Scholar] [CrossRef]
- Ding, T.D.; Yang, M.T.; Zhang, J.M.; Yang, B.; Lin, K.D.; Li, J.Y.; Gan, J. Toxicity, degradation and metabolic fate of ibuprofen on freshwater diatom Navicula sp. J. Hazard. Mater. 2017, 330, 127–134. [Google Scholar] [CrossRef]
- Goncalves, S.; Kahlert, M.; Almeida, S.F.P.; Figueira, E. Assessing Cu impacts on freshwater diatoms: Biochemical and metabolomic responses of Tabellaria flocculosa (Roth) Kutzing. Sci. Total Environ. 2018, 625, 1234–1246. [Google Scholar] [CrossRef]
- Meyer, N.; Rettner, J.; Werner, M.; Werz, O.; Pohnert, G. Algal oxylipins mediate the resistance of diatoms against algicidial bacteria. Mar. Drugs 2018, 16, 486. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.G.; Ansari, M.A.; Jamal, Q.M.S.; Almatroudi, A.; Alzohairy, M.A.; Alomary, M.N.; Rehman, S.; Mahadevamurthy, M.; Jalal, M.; Khan, H.M.; et al. Butea monosperma seed extract mediated biosynthesis of ZnO NPs and their antibacterial, antibiofilm and anti-quorum sensing potentialities. Arab. J. Chem. 2021, 14, 103044. [Google Scholar] [CrossRef]
- Alkhathlan, A.H.; Al-abdulkarim, H.A.; Khan, M.; Khan, M.; Alkhathlan, H.Z.; Siddiqui, M.R.H. Evaluation of the anticancer activity of phytomolecules conjugated gold nanoparticles synthesized by aqueous extracts of Zingiber officinale (ginger) and Nigella sativa L. seeds (black cumin). Materials 2021, 14, 3368. [Google Scholar] [CrossRef] [PubMed]
- Lebeau, T.; Gaudin, P.; Moan, R.; Robert, J.M. A new photobioreactor for continuous marennin production with a marine diatom: Influence of the light intensity and the immobilised-cell matrix (alginate beads or agar layer). Appl. Microbiol. Biotechnol. 2002, 59, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Kato, I.; Sudo, Y.; O’Shima, E. Extraction method of fatty acids from cells of Phaeodaclylum tricornutum. Nippon. Nogeikagaku Kaishi 1991, 65, 1489–1491. [Google Scholar] [CrossRef]
- Mutalipassi, M.; Mazzella, V.; Schott, M.; Fink, P.; Glaviano, F.; Porzio, L.; Lorenti, M.; Buia, M.C.; von Elert, E.; Zupo, V. Ocean acidification affects volatile infochemicals production and perception in fauna and flora associated with Posidonia oceanica (L.) Delile. Front. Mar. Sci. 2022, 9, 809702. [Google Scholar] [CrossRef]
- Sdrigotti, E.; Talarico, L. Excretion of exudates by a diatom under culture. Arch. Della Ric. Di Trieste 1994, 47–61. [Google Scholar]
- Wasternack, C.; Feussner, I. The oxylipin pathways: Biochemistry and function. Annu. Rev. Plant Biol. 2018, 69, 363–386. [Google Scholar] [CrossRef]
- Zupo, V. Co-evolution of the shrimp Hippolyte inermis and the diatoms Cocconeis spp. in Posidonia oceanica: Sexual adaptations explained by ecological fitting. Ref. Ser. Phytochem. 2020, 7, 135–147. [Google Scholar] [CrossRef]
- Nijjar, M.S.; Grimmelt, B.; Brown, J. Purification of domoic acid from toxic blue mussels (Mytilus edulis) and phytoplankton. J. Chromatogr. B Biomed. Sci. Appl. 1991, 568, 393–406. [Google Scholar] [CrossRef]
- Wainman, B.C.; Leant, D.R.S. Methodological concerns in measuring the lipid fraction of carbon fixation. Hydrobiologia 1994, 273, 111–120. [Google Scholar] [CrossRef]
- Li, Y.; Liu, L.; Sun, P.; Zhang, Y.; Wu, T.; Sun, H.; Cheng, K.W.; Chen, F. Fucoxanthinol from the diatom Nitzschia laevis ameliorates neuroinflammatory responses in lipopolysaccharide-stimulated BV-2 microglia. Mar. Drugs 2020, 18, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkia, I.; Al-Haj, L.; Abdul Hamid, A.; Zakaria, M.; Saari, N.; Zadjali, F. Indigenous marine diatoms as novel sources of bioactive peptides with antihypertensive and antioxidant properties. Int. J. Food Sci. Technol. 2019, 54, 1514–1522. [Google Scholar] [CrossRef]
- Tiwari, A.; Melchor-Martínez, E.M.; Saxena, A.; Kapoor, N.; Singh, K.J.; Saldarriaga-Hernández, S.; Parra-Saldívar, R.; Iqbal, H.M.N. Therapeutic attributes and applied aspects of biological macromolecules (polypeptides, fucoxanthin, sterols, fatty acids, polysaccharides, and polyphenols) from diatoms—A review. Int. J. Biol. Macromol. 2021, 171, 398–413. [Google Scholar] [CrossRef]
- Alzahrani, M.A.J.; Perera, C.O.; Hemar, Y. Production of bioactive proteins and peptides from the diatom Nitzschia laevis and comparison of their in vitro antioxidant activities with those from Spirulina platensis and Chlorella vulgaris. Int. J. Food Sci. Technol. 2018, 53, 676–682. [Google Scholar] [CrossRef]
- Neumann, U.; Derwenskus, F.; Flister, V.F.; Schmid-Staiger, U.; Hirth, T.; Bischoff, S.C. Fucoxanthin, a carotenoid derived from Phaeodactylum tricornutum exerts antiproliferative and antioxidant activities in vitro. Antioxidants 2019, 8, 183. [Google Scholar] [CrossRef] [Green Version]
- Świderska-Kołacz, G.; Jefimow, M.; Klusek, J.; Rączka, N.; Zmorzyński, S.; Wojciechowska, A.; Stanisławska, I.; Łyp, M.; Czerwik-Marcinkowska, J. Influence of algae supplementation on the concentration of glutathione and the activity of glutathione enzymes in the mice liver and kidney. Nutrients 2021, 13, 1996. [Google Scholar] [CrossRef]
- Pekkoh, J.; Phinyo, K.; Thurakit, T.; Lomakool, S.; Duangjan, K.; Ruangrit, K.; Pumas, C.; Jiranusornkul, S.; Yooin, W.; Cheirsilp, B.; et al. Lipid profile, antioxidant and antihypertensive activity, and computational molecular docking of diatom fatty acids as ACE inhibitors. Antioxidants 2022, 11, 186. [Google Scholar] [CrossRef]
- Stiefvatter, L.; Neumann, U.; Rings, A.; Frick, K.; Schmid-Staiger, U.; Bischoff, S.C. The microalgae Phaeodactylum tricornutum is well suited as a food with positive effects on the intestinal microbiota and the generation of SCFA: Results from a pre-clinical study. Nutrients 2022, 14, 2504. [Google Scholar] [CrossRef]
- Lee, A.H.; Shin, H.Y.; Park, J.H.; Koo, S.Y.; Kim, S.M.; Yang, S.H. Fucoxanthin from microalgae Phaeodactylum tricornutum inhibits pro-inflammatory cytokines by regulating both NF-κB and NLRP3 inflammasome activation. Sci. Rep. 2021, 11, 543. [Google Scholar] [CrossRef]
- Figueroa, F.A.; Abdala-Díaz, R.; Hernández, V.; Pedreros, P.; Aranda, M.; Cabrera-Pardo, J.R.; Pérez, C.; Becerra, J.; Urrutia, R. Invasive diatom Didymosphenia geminata as a source of polysaccharides with antioxidant and immunomodulatory effects on macrophage cell lines. J. Appl. Phycol. 2020, 32, 93–102. [Google Scholar] [CrossRef]
- Lai, H.L.; Yang, L.C.; Lin, P.T.; Lai, S.Y.; Wang, M.Y. Phagocytosis activity of three sulfated polysaccharides purified from a marine diatom cultured in a semi-continuous system. Int. J. Biol. Macromol. 2020, 155, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Bergé, J.P.; Bourgougnon, N.; Alban, S.; Pojer, F.; Billaudel, S.; Chermann, J.C.; Robert, J.M.; Franz, G. Antiviral and anticoagulant activities of a water-soluble fraction of the marine diatom Haslea ostrearia. Planta Med. 1999, 65, 604–609. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Hayashi, K.; Hirata, M.; Kuroda, E.; Suzuki, E.; Kubo, Y.; Hayashi, T. Antiviral sulfated polysaccharide from Navicula directa, a diatom collected from deep-sea water in Toyama Bay. Biol. Pharm. Bull. 2006, 29, 2135–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fimbres-Olivarria, D.; Carvajal-Millan, E.; Lopez-Elias, J.A.; Martinez-Robinson, K.G.; Miranda-Baeza, A.; Martinez-Cordova, L.R.; Enriquez-Ocaña, F.; Valdez-Holguin, J.E. Chemical characterization and antioxidant activity of sulfated polysaccharides from Navicula sp. Food Hydrocoll. 2018, 75, 229–236. [Google Scholar] [CrossRef]
- Xia, S.; Gao, B.; Li, A.; Xiong, J.; Ao, Z.; Zhang, C. Preliminary characterization, antioxidant properties and production of chrysolaminarin from marine diatom Odontella Aurita. Mar. Drugs 2014, 12, 4883–4897. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, S.; Gato, A.; Lamela, M.; Freire-Garabal, M.; Calleja, J.M. Anti-inflammatory and immunomodulatory activities of polysaccharide from Chlorella stigmatophora and Phaeodactylum tricornutum. Phyther. Res. 2003, 17, 665–670. [Google Scholar] [CrossRef]
- Carballo, C.; Chronopoulou, E.G.; Letsiou, S.; Maya, C.; Labrou, N.E.; Infante, C.; Power, D.M.; Manchado, M. Antioxidant capacity and immunomodulatory effects of a chrysolaminarin-enriched extract in Senegalese sole. Fish Shellfish. Immunol. 2018, 82, 1–8. [Google Scholar] [CrossRef]
- Stiefvatter, L.; Frick, K.; Lehnert, K.; Vetter, W.; Montoya-Arroyo, A.; Frank, J.; Schmid-Staiger, U.; Bischoff, S.C. Potentially beneficial effects on healthy aging by supplementation of the EPA-rich microalgae Phaeodactylum tricornutum or its Supernatant-A Randomized controlled pilot trial in elderly individuals. Mar. Drugs 2022, 20, 716. [Google Scholar] [CrossRef]
- Kusaikin, M.I.; Ermakova, S.P.; Shevchenko, N.M.; Isakov, V.V.; Gorshkov, A.G.; Vereshchagin, A.L.; Grachev, M.A.; Zvyagintseva, T.N. Structural characteristics and antitumor activity of a new chrysolaminaran from the diatom alga Synedra acus. Chem. Nat. Compd. 2010, 46, 1–4. [Google Scholar] [CrossRef]
- Li, H.B.; Cheng, K.W.; Wong, C.C.; Fan, K.W.; Chen, F.; Jiang, Y. Evaluation of antioxidant capacity and total phenolic content of different fractions of selected microalgae. Food Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]
- Rico, M.; López, A.; Santana-Casiano, J.M.; González, A.G.; González-Dávila, M. Variability of the phenolic profile in the diatom Phaeodactylum tricornutum growing under copper and iron stress. Limnol. Oceanogr. 2013, 58, 144–152. [Google Scholar] [CrossRef]
- Smerilli, A.; Balzano, S.; Maselli, M.; Blasio, M.; Orefice, I.; Galasso, C.; Sansone, C.; Brunet, C. Antioxidant and photoprotection networking in the coastal diatom Skeletonema marinoi. Antioxidants 2019, 8, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, P.; Cleary, D.F.R.; Pires, A.C.C.; Rodrigues, A.M.; Quintino, V.; Calado, R.; Gomes, N.C.M. Molecular analysis of bacterial communities and detection of potential pathogens in a recirculating aquaculture system for Scophthalmus maximus and Solea senegalensis. PLoS ONE 2013, 8, e80847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Fulgoni, V.L.; Kris-Etherton, P.M.; Mitmesser, S.H. Dietary intakes of EPA and DHA omega-3 fatty acids among US childbearing-age and pregnant women: An analysis of NHANES 2001–2014. Nutrients 2018, 10, 416. [Google Scholar] [CrossRef] [Green Version]
- SanGiovanni, J.P.; Berkey, C.S.; Dwyer, J.T.; Colditz, G.A. Dietary essential fatty acids, long-chain polyunsaturated fatty acids, and visual resolution acuity in healthy fullterm infants: A systematic review. Early Hum. Dev. 2000, 57, 165–188. [Google Scholar] [CrossRef] [PubMed]
- Yurko-Mauro, K.; McCarthy, D.; Rom, D.; Nelson, E.B.; Ryan, A.S.; Blackwell, A.; Salem, N.; Stedman, M. Beneficial effects of docosahexaenoic acid on cognition in age-related cognitive decline. Alzheimer’s Dement. 2010, 6, 456–464. [Google Scholar] [CrossRef]
- Brett, M.T.; Müller-Navarra, D.C. The role of highly unsaturated fatty acids in aquatic foodweb processes. Freshw. Biol. 1997, 38, 483–499. [Google Scholar] [CrossRef]
- Tocher, D.R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Nevigato, T.; Masci, M.; Caproni, R. Quality of fish-oil-based dietary supplements available on the italian market: A preliminary study. Molecules 2021, 26, 5015. [Google Scholar] [CrossRef]
- Lebeau, T.; Robert, J.M. Diatom cultivation and biotechnologically relevant products. Part II: Current and putative products. Appl. Microbiol. Biotechnol. 2003, 60, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Belarbi, E.H.; Molina, E.; Chisti, Y. RETRACTED: A process for high yield and scaleable recovery of high purity eicosapentaenoic acid esters from microalgae and fish oil. Process Biochem. 2000, 35, 951–969. [Google Scholar] [CrossRef]
- Piepho, M.; Arts, M.T.; Wacker, A. Species-specific variation in fatty acid concentrations of four phytoplankton species: Does phosphorus supply influence the effect of light intensity or temperature? J. Phycol. 2012, 48, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A. Marine natural products. Annu. Rep. Prog. Chem.—Sect. B 2011, 107, 138–156. [Google Scholar] [CrossRef]
- Renaud, S.M.; Zhou, H.C.; Parry, D.L.; Van Thinh, L.; Woo, K.C. Effect of temperature on the growth, total lipid content and fatty acid composition of recently isolated tropical microalgae Isochrysis sp., Nitzschia closterium, Nitzschia paleacea, and commercial species Isochrysis sp. (clone T.ISO). J. Appl. Phycol. 1995, 7, 595–602. [Google Scholar] [CrossRef]
- Kates, M.; Volcani, R.E. Lipid components of diatoms. Biochim. Biophys. Acta 1965, 116, 264–278. [Google Scholar] [CrossRef]
- Li, H.Y.; Lu, Y.; Zheng, J.W.; Yang, W.D.; Liu, J.S. Biochemical and genetic engineering of diatoms for polyunsaturated fatty acid biosynthesis. Mar. Drugs 2014, 12, 153–166. [Google Scholar] [CrossRef]
- Zupo, V.; Jüttner, F.; Maibam, C.; Butera, E.; Blom, J.F. Apoptogenic metabolites in fractions of the benthic diatom Cocconeis scutellum parva. Mar. Drugs 2014, 12, 547–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zupo, V. Strategies of sexual inversion in Hippolyte inermis Leach (Crustacea, Decapoda) from a Mediterranean seagrass meadow. J. Exp. Mar. Bio. Ecol. 1994, 178, 131–145. [Google Scholar] [CrossRef]
- Clifton, P. Dietary fatty acids and inflammation. Nutr. Diet. 2009, 66, 7–11. [Google Scholar] [CrossRef]
- Chang, E.; Kim, Y. Vitamin D decreases adipocyte lipid storage and increases NAD-SIRT1 pathway in 3T3-L1 adipocytes. Nutrition 2016, 32, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Darnet, S.; Blary, A.; Chevalier, Q.; Schaller, H. Phytosterol profiles, genomes and enzymes—An overview. Front. Plant Sci. 2021, 12, 878. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.; Park, S.Y.; Sun, Z.W.; Shin, H.S.; Lee, D.G.; Yi, T.H. The protective effects of fucosterol against skin damage in UVB-irradiated human dermal fibroblasts. Mar. Biotechnol. 2014, 16, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Khanavi, M.; Gheidarloo, R.; Sadati, N.; Shams Ardekani, M.R.; Bagher Nabavi, S.M.; Tavajohi, S.; Ostad, S.N. Cytotoxicity of fucosterol containing fraction of marine algae against breast and colon carcinoma cell line. Pharmacogn. Mag. 2012, 8, 60–64. [Google Scholar] [CrossRef] [Green Version]
- Galasso, C.; Gentile, A.; Orefice, I.; Ianora, A.; Bruno, A.; Noonan, D.M.; Sansone, C.; Albini, A.; Brunet, C. Microalgal derivatives as potential nutraceutical and food supplements for human health: A focus on cancer prevention and interception. Nutrients 2019, 11, 1226. [Google Scholar] [CrossRef] [Green Version]
- Rampen, S.W.; Abbas, B.A.; Schouten, S.; Damsté, J.S.S. A comprehensive study of sterols in marine diatoms (Bacillariophyta): Implications for their use as tracers for diatom productivity. Limnol. Oceanogr. 2010, 55, 91–105. [Google Scholar] [CrossRef]
- Jaramillo-Madrid, A.C.; Abbriano, R.; Ashworth, J.; Fabris, M.; Pernice, M.; Ralph, P.J. Overexpression of key sterol pathway enzymes in two model marine diatoms alters sterol profiles in Phaeodactylum tricornutum. Pharmaceuticals 2020, 13, 481. [Google Scholar] [CrossRef]
- Gallo, C.; Landi, S.; d’Ippolito, G.; Nuzzo, G.; Manzo, E.; Sardo, A.; Fontana, A. Diatoms synthesize sterols by inclusion of animal and fungal genes in the plant pathway. Sci. Rep. 2020, 10, 4204. [Google Scholar] [CrossRef] [Green Version]
- Nuzzo, G.; Gallo, C.; D’Ippolito, G.; Manzo, E.; Ruocco, N.; Russo, E.; Carotenuto, Y.; Costantini, M.; Zupo, V.; Sardo, A.; et al. UPLC–MS/MS identification of sterol sulfates in marine diatoms. Mar. Drugs 2019, 17, 10. [Google Scholar] [CrossRef] [Green Version]
- Gügi, B.; Le Costaouec, T.; Burel, C.; Lerouge, P.; Helbert, W.; Bardor, M. Diatom-specific oligosaccharide and polysaccharide structures help to unravel biosynthetic capabilities in diatoms. Mar. Drugs 2015, 13, 5993–6018. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Weng, D.; Du, C.; Wang, J.; Cao, S. Contribution of frustules and mucilage trails to the mobility of diatom Navicula sp. Sci. Rep. 2019, 9, 7342. [Google Scholar] [CrossRef] [PubMed]
- Tounsi, L.; Hentati, F.; Ben Hlima, H.; Barkallah, M.; Smaoui, S.; Fendri, I.; Michaud, P.; Abdelkafi, S. Microalgae as feedstock for bioactive polysaccharides. Int. J. Biol. Macromol. 2022, 221, 1238–1250. [Google Scholar] [CrossRef] [PubMed]
- Severo, I.A.; Dias, R.R.; do Nascimento, T.C.; Deprá, M.C.; Maroneze, M.M.; Zepka, L.Q.; Jacob-Lopes, E. Microalgae-derived polysaccharides: Potential building blocks for biomedical applications. World J. Microbiol. Biotechnol. 2022, 38, 150. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Wei, D.; Xie, J. Diatoms as cell factories for high-value products: Chrysolaminarin, eicosapentaenoic acid, and fucoxanthin. Crit. Rev. Biotechnol. 2020, 40, 993–1009. [Google Scholar] [CrossRef]
- Butler, T.; Kapoore, R.V.; Vaidyanathan, S. Phaeodactylum tricornutum: A diatom cell factory. Trends Biotechnol. 2020, 38, 606–622. [Google Scholar] [CrossRef]
- De Jesus Raposo, M.F.; De Morais, R.M.S.C.; De Morais, A.M.M.B. Bioactivity and applications of sulphated polysaccharides from marine microalgae. Mar. Drugs 2013, 11, 233–252. [Google Scholar] [CrossRef] [Green Version]
- Hao, C.; Yu, G.; He, Y.; Xu, C.; Zhang, L.; Wang, W. Marine glycan–based antiviral agents in clinical or preclinical trials. Rev. Med. Virol. 2019, 29, e2043. [Google Scholar] [CrossRef]
- Riccio, G.; Lauritano, C. Microalgae with immunomodulatory activities. Mar. Drugs 2020, 18, 2. [Google Scholar] [CrossRef] [Green Version]
- Panggabean, J.A.; Adiguna, S.P.; Rahmawati, S.I.; Ahmadi, P.; Zainuddin, E.N.; Bayu, A.; Putra, M.Y. Antiviral activities of algal-based sulfated polysaccharides. Molecules 2022, 27, 1178. [Google Scholar] [CrossRef]
- Guzmán, S.; Gato, A.; Calleja, J.M. Antiinflammatory, analgesic and free radical scavenging activities of the marine microalgae Chlorella stigmatophora and Phaeodactylum tricornutum. Phyther. Res. 2001, 15, 224–230. [Google Scholar] [CrossRef]
- Kuczynska, P.; Jemiola-Rzeminska, M.; Strzalka, K. Photosynthetic pigments in diatoms. Mar. Drugs 2015, 13, 5847–5881. [Google Scholar] [CrossRef] [PubMed]
- Bayu, A.; Rachman, A.; Noerdjito, D.R.; Putra, M.Y.; Widayatno, W.B. High-value chemicals from marine diatoms: A biorefinery approach. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Raipur, India, 25–26 February 2022; IOP Publishing: Bristol, UK, 2020. [Google Scholar]
- Seth, K.; Kumar, A.; Rastogi, R.P.; Meena, M.; Vinayak, V. Harish Bioprospecting of fucoxanthin from diatoms—Challenges and perspectives. Algal Res. 2021, 60, 102475. [Google Scholar] [CrossRef]
- Aronoff, S. The absorption spectra of chlorophyll and related compounds. Chem. Rev. 1950, 47, 175–195. [Google Scholar] [CrossRef] [PubMed]
- Ambati, R.R.; Gogisetty, D.; Aswathanarayana, R.G.; Ravi, S.; Bikkina, P.N.; Bo, L.; Yuepeng, S. Industrial potential of carotenoid pigments from microalgae: Current trends and future prospects. Crit. Rev. Food Sci. Nutr. 2019, 59, 1880–1902. [Google Scholar] [CrossRef] [PubMed]
- Mumu, M.; Das, A.; Bin Emran, T.; Mitra, S.; Islam, F.; Roy, A.; Karim, M.M.; Das, R.; Park, M.N.; Chandran, D.; et al. Fucoxanthin: A promising phytochemical on diverse pharmacological targets. Front. Pharmacol. 2022, 13, 929442. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; Ozaki, Y.; Taminato, M.; Das, S.K.; Mizuno, M.; Yoshimura, K.; Maoka, T.; Kanazawa, K. The distribution and accumulation of fucoxanthin and its metabolites after oral administration in mice. Br. J. Nutr. 2009, 102, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, J.; Yuan, J.P.; Wu, C.F.; Wang, J.H. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Funayama, K.; Miyashita, K. Effect of medium-chain triacyglycerols on anto-obesity effect of fucoxanthin. J. Oleo Sci. 2007, 621, 615–621. [Google Scholar] [CrossRef] [Green Version]
- Abidov, M.; Ramazanov, Z.; Seifulla, R.; Grachev, S. The effects of XanthigenTM in the weight management of obese premenopausal women with non-alcoholic fatty liver disease and normal liver fat. Diabetes Obes. Metab. 2010, 12, 72–81. [Google Scholar] [CrossRef]
- Asai, A.; Yonekura, L.; Nagao, A. Low bioavailability of dietary epoxyxanthophylls in humans. Br. J. Nutr. 2008, 100, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Beppu, F.; Niwano, Y.; Tsukui, T.; Hosokawa, M.; Miyashita, K. Single and repeated oral dose toxicity study of fucoxanthin (FX), a marine carotenoid, in mice. J. Toxicol. Sci. 2009, 34, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Kumiko, I.; Yumika, O.; Masaharu, I. Bacterial reverse mutation test and micronucleus test of fucoxanthin oil from microalgae. Shokuhin Eiseigaku Zasshi 2011, 52, 190–193. [Google Scholar] [CrossRef] [Green Version]
- Beppu, F.; Niwano, Y.; Sato, E.; Kohno, M.; Tsukui, T.; Hosokawa, M.; Miyashita, K. In vitro and in vivo evaluation of mutagenicity of fucoxanthin (FX) and its metabolite fucoxanthinol (FXOH). J. Toxicol. Sci. 2009, 34, 693–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiefvatter, L.; Lehnert, K.; Frick, K.; Montoya-Arroyo, A.; Frank, J.; Vetter, W.; Schmid-Staiger, U.; Bischoff, S.C. Oral bioavailability of omega-3 fatty acids and carotenoids from the microalgae Phaeodactylum tricornutum in healthy young adults. Mar. Drugs 2021, 19, 700. [Google Scholar] [CrossRef] [PubMed]
- Lykov, A.; Rachkovsky, E.; Gevorgiz, R.; Zheleznova, S.; Kotlyarova, A. Toxicity of fucoxanthin on Balb/c mice splenocytes and thymocytes. In 2020 Cognitive Sciences, Genomics and Bioinformatics (CSGB); IEEE: Piscataway, NJ, USA, 2020; pp. 277–280. [Google Scholar] [CrossRef]
- Guan, B.; Chen, K.; Tong, Z.; Chen, L.; Chen, Q.; Su, J. Advances in fucoxanthin research for the prevention and treatment of inflammation-related diseases. Nutrients 2022, 14, 4768. [Google Scholar] [CrossRef]
- Mohibbullah, M.; Haque, M.N.; Sohag, A.A.M.; Hossain, M.T.; Zahan, M.S.; Uddin, M.J.; Hannan, M.A.; Moon, I.S.; Choi, J.S. A systematic review on marine algae-derived fucoxanthin: An update of pharmacological insights. Mar. Drugs 2022, 20, 279. [Google Scholar] [CrossRef]
- Oliyaei, N.; Moosavi-nasab, M.; Tanideh, N.; Iraji, A. Multiple roles of fucoxanthin and astaxanthin against Alzheimer ’ s disease: Their pharmacological potential and therapeutic insights. Brain Res. Bull. 2022, 193, 11–21. [Google Scholar] [CrossRef]
- Yuan, Y.; Ma, M.; Zhang, S. Recent advances in delivery systems of fucoxanthin. Food Chem. 2022, 404, 134685. [Google Scholar] [CrossRef]
- Li, C.Z.E.; Deng, Z.W. The Embedded modules solution of household internet of things system and the future development. Procedia Comput. Sci. 2020, 166, 350–356. [Google Scholar] [CrossRef]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chemie—Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; De Brabanter, J.; De Cooman, L. Antioxidant potential of microalgae in relation to their phenolic and carotenoid content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Agati, G.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Pollastri, S.; Tattini, M. Functional roles of flavonoids in photoprotection: New evidence, lessons from the past. Plant Physiol. Biochem. 2013, 72, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Mutalipassi, M.; Mazzella, V.; Romano, G.; Ruocco, N.; Costantini, M.; Glaviano, F.; Zupo, V. Growth and toxicity of Halomicronema metazoicum (Cyanoprokaryota, Cyanophyta) at different conditions of light, salinity and temperature. Biol. Open 2019, 8, bio043604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thangaraj, S.; Sun, J. The biotechnological potential of the marine diatom Skeletonema dohrnii to the elevated temperature and pCO2. Mar. Drugs 2020, 18, 259. [Google Scholar] [CrossRef] [PubMed]
- Nur, M.M.A.; Muizelaar, W.; Boelen, P.; Buma, A.G.J. Environmental and nutrient conditions influence fucoxanthin productivity of the marine diatom Phaeodactylum tricornutum grown on palm oil mill effluent. J. Appl. Phycol. 2019, 31, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Mal, N.; Srivastava, K.; Sharma, Y.; Singh, M.; Rao, K.M.; Enamala, M.K.; Chandrasekhar, K.; Chavali, M. Facets of diatom biology and their potential applications. Biomass Convers. Biorefinery 2022, 12, 1959–1975. [Google Scholar] [CrossRef]
- Matsumoto, M.; Nojima, D.; Nonoyama, T.; Ikeda, K.; Maeda, Y.; Yoshino, T.; Tanaka, T. Outdoor cultivation of marine diatoms for year-round production of biofuels. Mar. Drugs 2017, 15, 8–10. [Google Scholar] [CrossRef]
- Chiriboga, O.; Rorrer, G.L. Effects of nitrogen delivery on chitin nanofiber production during batch cultivation of the diatom Cyclotella sp. in a bubble column photobioreactor. J. Appl. Phycol. 2018, 30, 1575–1581. [Google Scholar] [CrossRef]
- Sasso, S.; Pohnert, G.; Lohr, M.; Mittag, M.; Hertweck, C. Microalgae in the postgenomic era: A blooming reservoir for new natural products. FEMS Microbiol. Rev. 2012, 36, 761–785. [Google Scholar] [CrossRef]
- Hagerthey, S.E.; William Louda, J.; Mongkronsri, P. Evaluation of pigment extraction methods and a recommended protocol for periphyton chlorophyll a determination and chemotaxonomic assessment. J. Phycol. 2006, 42, 1125–1136. [Google Scholar] [CrossRef]
- Cutignano, A.; Lamari, N.; D’ippolito, G.; Manzo, E.; Cimino, G.; Fontana, A. Lipoxygenase products in marine diatoms: A concise analytical method to explore the functional potential of oxylipins. J. Phycol. 2011, 47, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Nappo, M.; Berkov, S.; Codina, C.; Avila, C.; Messina, P.; Zupo, V.; Bastida, J. Metabolite profiling of the benthic diatom Cocconeis scutellum by GC-MS. J. Appl. Phycol. 2009, 21, 295–306. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nieri, P.; Carpi, S.; Esposito, R.; Costantini, M.; Zupo, V. Bioactive Molecules from Marine Diatoms and Their Value for the Nutraceutical Industry. Nutrients 2023, 15, 464. https://doi.org/10.3390/nu15020464
Nieri P, Carpi S, Esposito R, Costantini M, Zupo V. Bioactive Molecules from Marine Diatoms and Their Value for the Nutraceutical Industry. Nutrients. 2023; 15(2):464. https://doi.org/10.3390/nu15020464
Chicago/Turabian StyleNieri, Paola, Sara Carpi, Roberta Esposito, Maria Costantini, and Valerio Zupo. 2023. "Bioactive Molecules from Marine Diatoms and Their Value for the Nutraceutical Industry" Nutrients 15, no. 2: 464. https://doi.org/10.3390/nu15020464
APA StyleNieri, P., Carpi, S., Esposito, R., Costantini, M., & Zupo, V. (2023). Bioactive Molecules from Marine Diatoms and Their Value for the Nutraceutical Industry. Nutrients, 15(2), 464. https://doi.org/10.3390/nu15020464