Antitumor Activity of Carboxymethyl Pachymaran with Different Molecular Weights Based on Immunomodulatory and Gut Microbiota


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Degradation Treatments of CMP
2.3. Molecular Weight Determination
2.4. Apparent Viscosity Analysis
2.5. Particle Size Distribution (PSD)
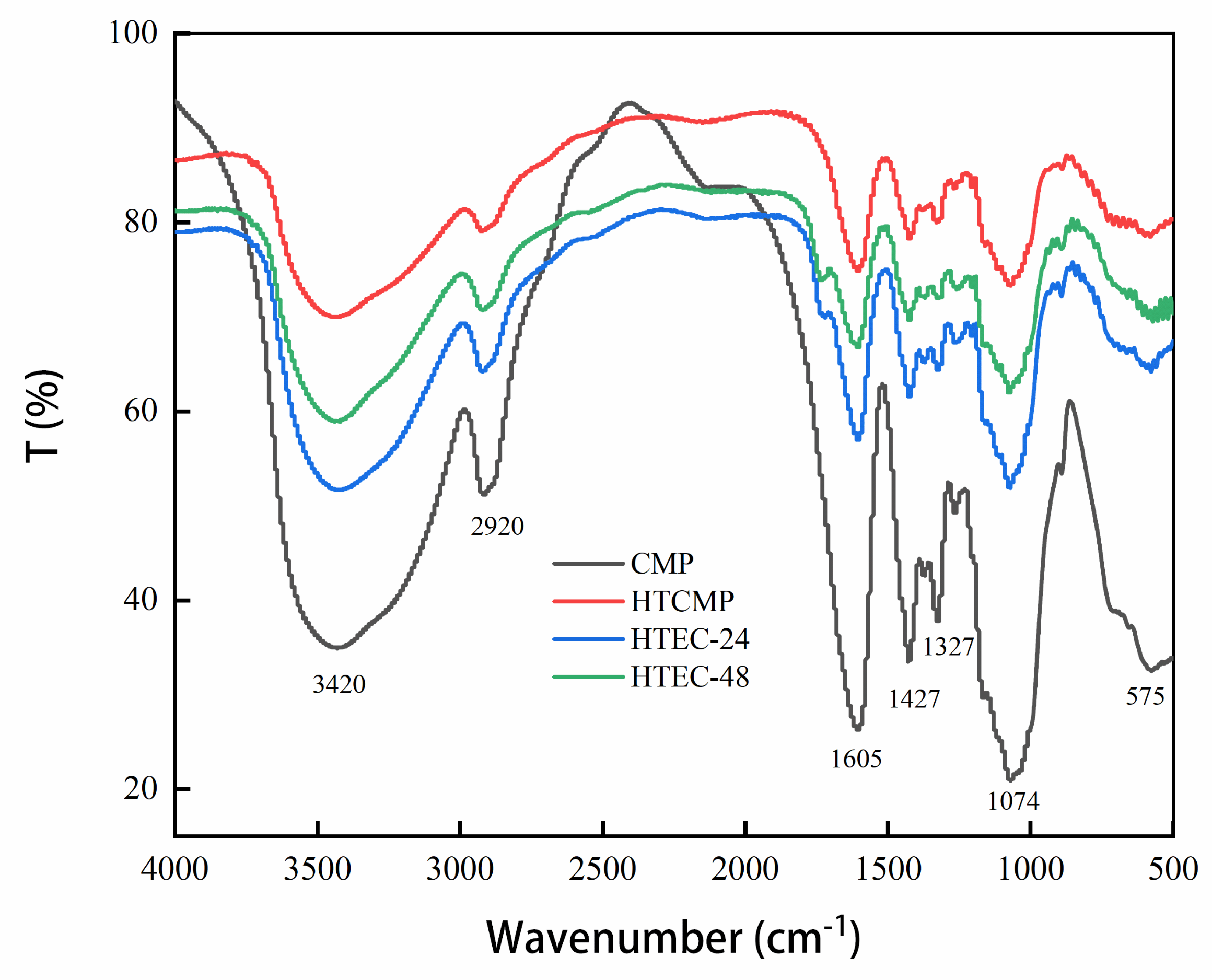
2.6. Fourier Transform Infrared Spectra (FT-IR) Assay
2.7. Antitumor Activity In Vivo
2.7.1. H22 Cell Culture
2.7.2. Animals and Experimental Design
2.7.3. The Inhibition Rate
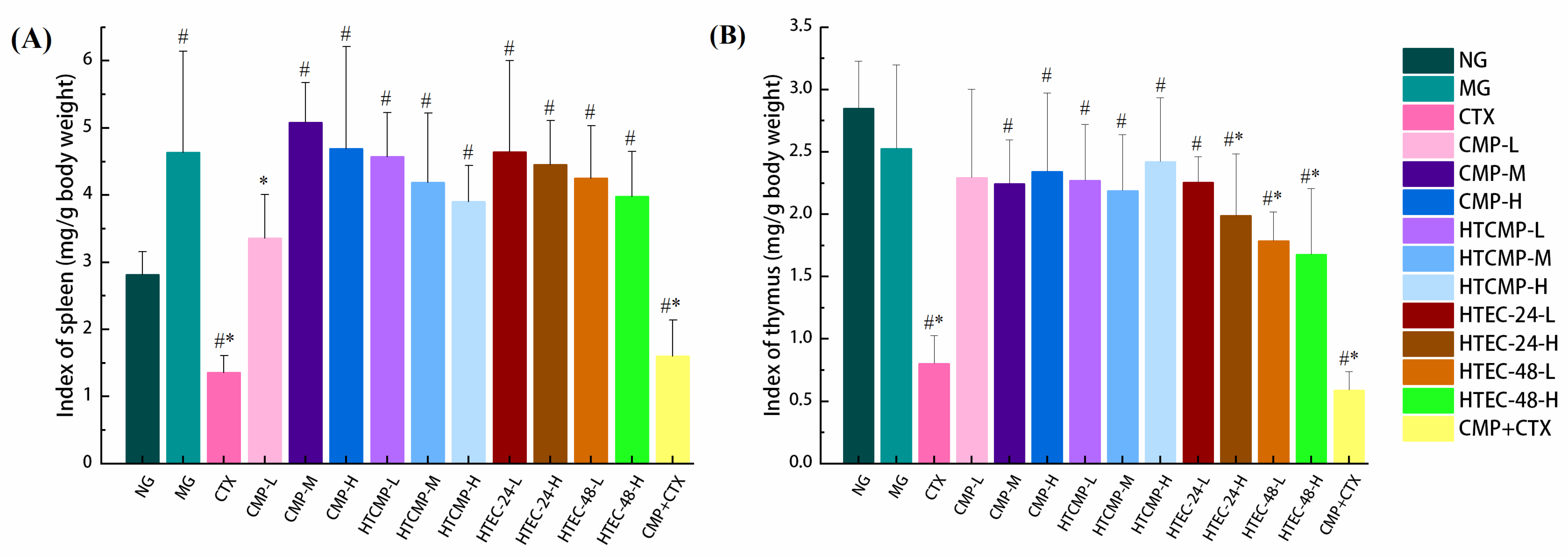
2.7.4. Determination of Organ Index
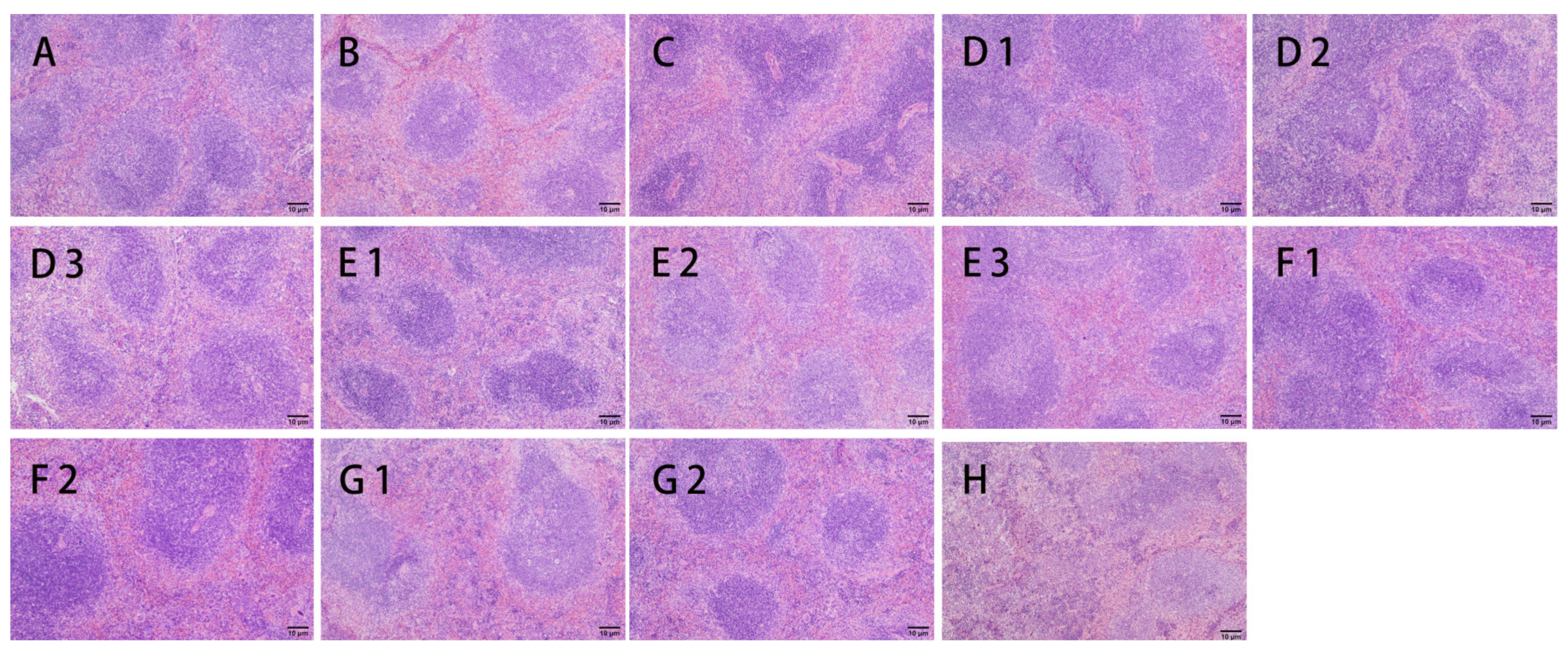
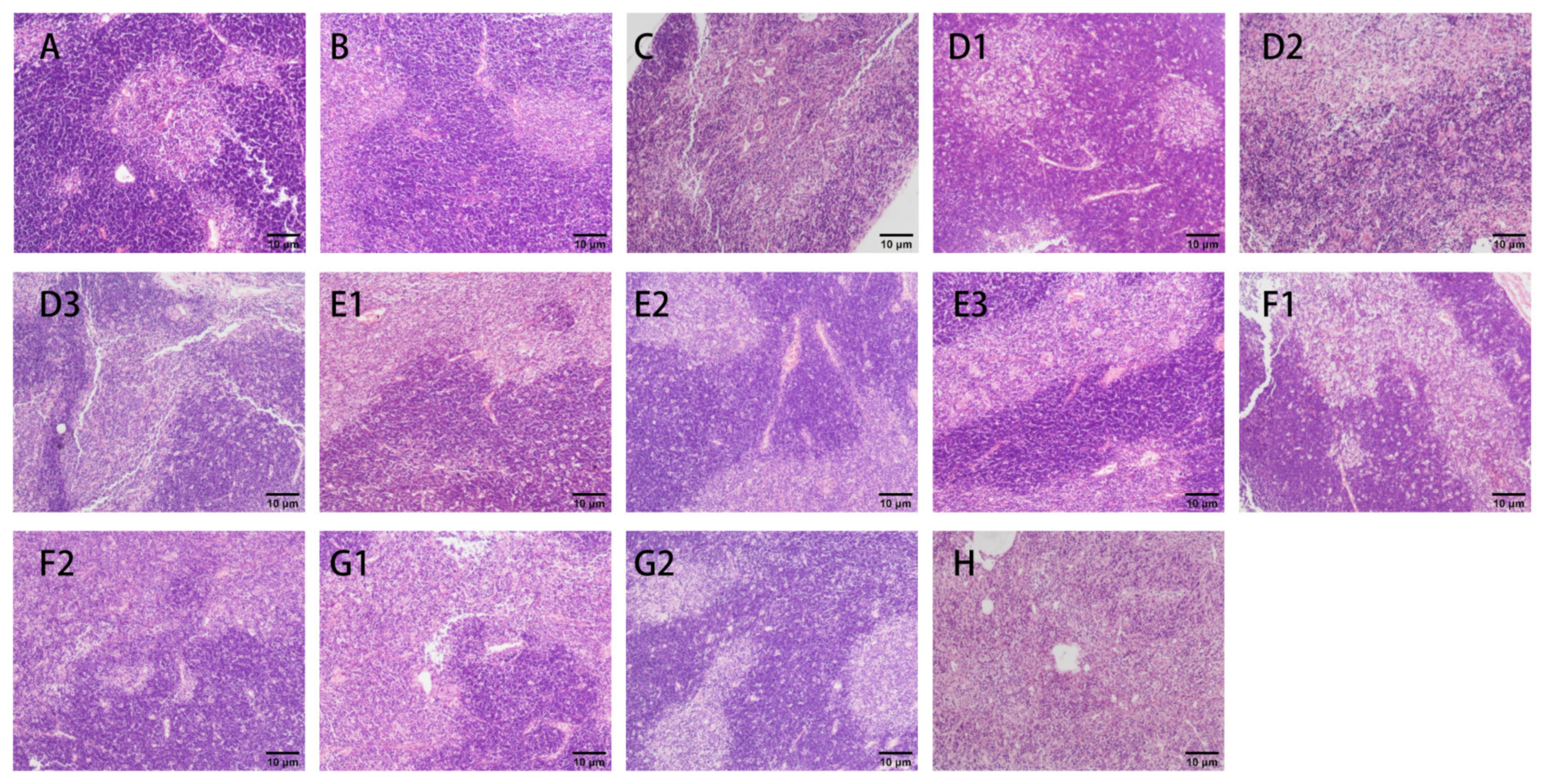
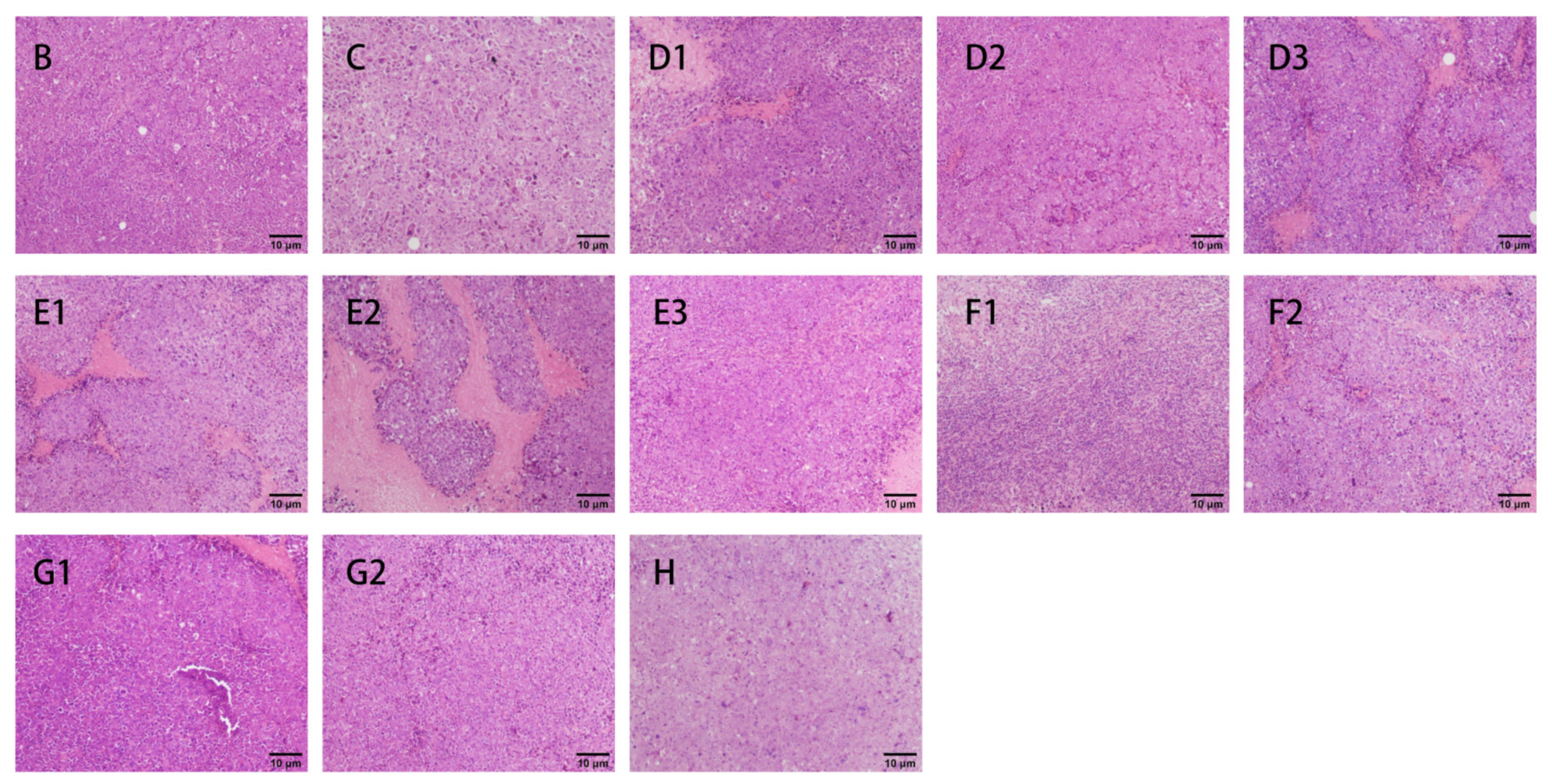
2.7.5. H&E Assays
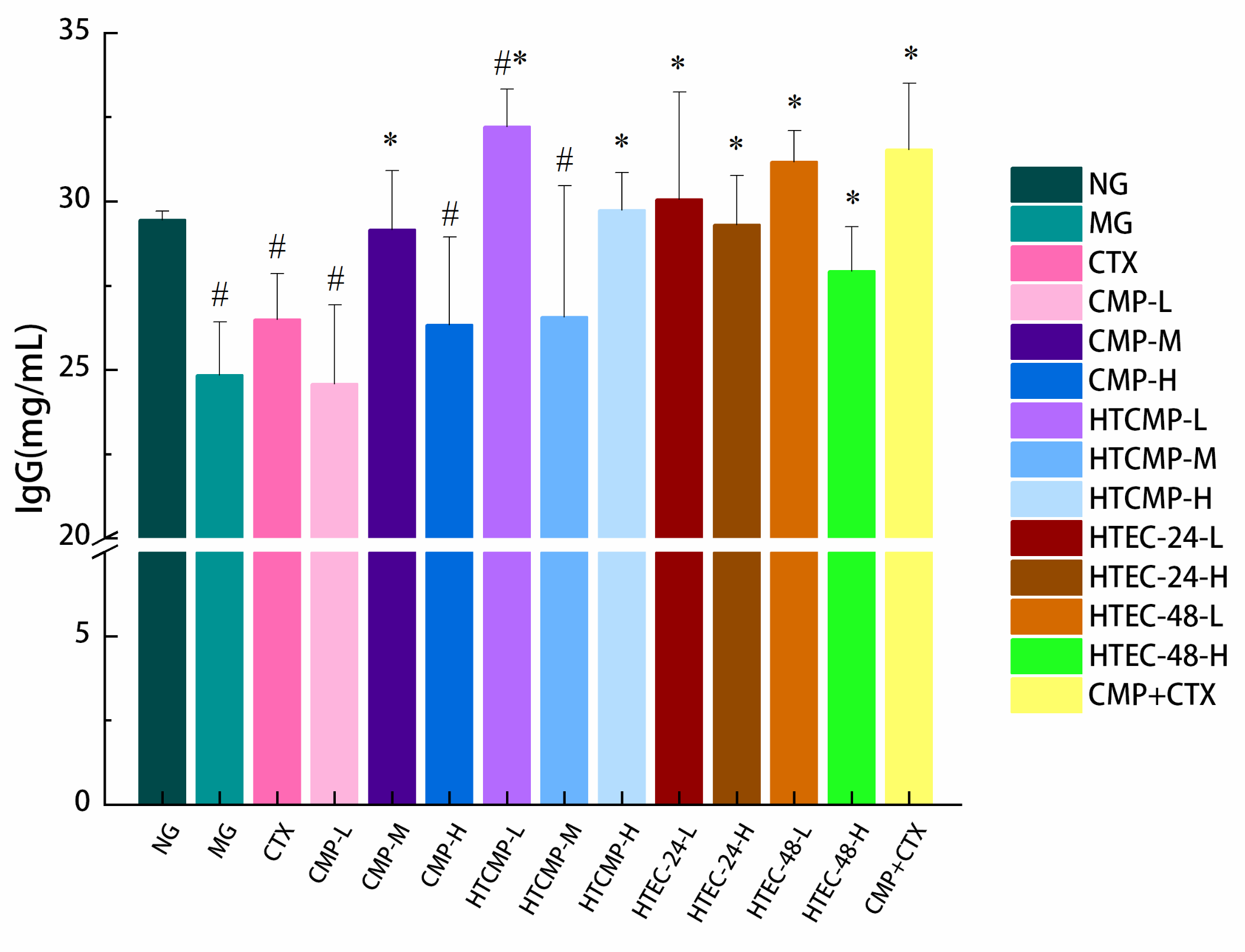
2.7.6. Measurement of Serum Immunoglobulin and Cytokines
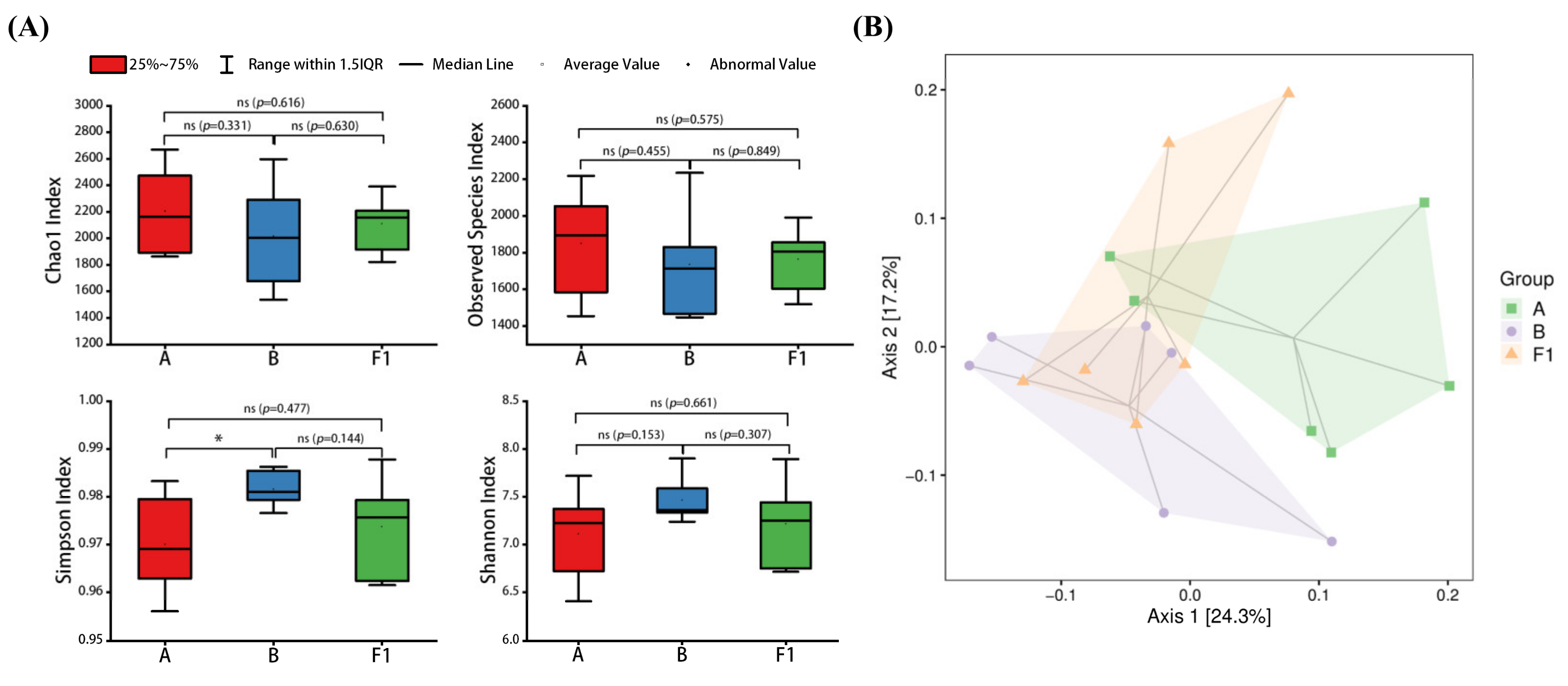
2.7.7. Gut Microbiota Analysis
2.7.8. Statistical Analysis
3. Results and Discussion
3.1. Structural Characterization of CMPs
3.1.1. Molecular Weights of CMPs
3.1.2. The Apparent Viscosity of CMPs
3.1.3. The Particle Size Distribution
3.1.4. FT-IR Spectra of CMPs
3.2. Effects of CMPs in H22 Tumor-Bearing Mice
3.2.1. Effect of CMPs on Tumor Inhibition Rate in Tumor-Bearing Mice
3.2.2. Effect of CMPs on Organ Indexes in Tumor-Bearing Mice
3.2.3. H&E Staining
3.2.4. Analysis of Serum Indicators and Cytokine Assays
3.2.5. Effect of CMPs on Immunoglobulin Content in Serum of Tumor-Bearing Mice
3.3. Effect of HTEC-24 on Gut Microbiota of Tumor-Bearing Mice
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, G.; Chen, Y.H.; Bao, Q.Y.; Jiang, Z.D.; Zhu, Y.B.; Ni, H.; Li, Q.B.; Oda, T. A low-molecular-weight ascophyllan prepared from Ascophyllum nodosum: Optimization, analysis and biological activities. Int. J. Biol. Macromol. 2020, 153, 107–117. [Google Scholar] [CrossRef]
- Son, S.U.; Kim, H.W.; Shin, K.S. Structural identification of active moiety in anti-tumor metastatic polysaccharide purified from fermented barley by sequential enzymatic hydrolysis. Food Biosci. 2022, 50, 101999. [Google Scholar] [CrossRef]
- Wang, Z.C.; Zhou, X.Y.; Shu, Z.H.; Zheng, Y.; Hu, X.L.; Zhang, P.Y.; Huang, H.T.; Sheng, L.L.; Zhang, P.S.; Wang, Q.; et al. Regulation strategy, bioactivity, and physical property of plant and microbial polysaccharides based on molecular weight. Int. J. Biol. Macromol. 2023, 244, 125360. [Google Scholar] [CrossRef] [PubMed]
- Perez, A.A.; Sánchez, C.C.; Rodríguez Patino, J.M.; Rubiolo, A.C.; Santiago, L.G. Effect of enzymatic hydrolysis and polysaccharide addition on the β-lactoglobulin adsorption at the air–water interface. J. Food Eng. 2012, 109, 712–720. [Google Scholar] [CrossRef]
- Zeng, S.H.; Long, J.W.; Sun, J.H.; Wang, G.; Zhou, L. A review on peach gum polysaccharide: Hydrolysis, structure, properties and applications. Carbohydr. Polym. 2022, 279, 119015. [Google Scholar] [CrossRef]
- Tian, W.L.; Song, X.L.; Wang, F.S.; Jiang, W.J. Study on the preparation and biological activities of low molecular weight squid ink polysaccharide from Sepiella maindroni. Int. J. Biol. Macromol. 2023, 237, 124040. [Google Scholar] [CrossRef]
- Xu, T.R.; Zhang, H.M.; Wang, S.G.; Xiang, Z.D.; Kong, H.W.; Xue, Q.; He, M.Y.; Yu, X.J.; Li, Y.N.; Sun, D.J.; et al. A review on the advances in the extraction methods and structure elucidation of Poria cocos polysaccharide and its pharmacological activities and drug carrier applications. Int. J. Biol. Macromol. 2022, 217, 536–551. [Google Scholar] [CrossRef]
- Zhu, L.X.; Xu, J.; Zhang, S.J.; Wang, R.J.; Huang, Q.; Chen, H.B.; Dong, X.P.; Zhao, Z.Z. Qualitatively and quantitatively comparing secondary metabolites in three medicinal parts derived from Poria cocos (Schw.) Wolf using UHPLC-QTOF-MS/MS-based chemical profiling. J. Pharm. Biomed. Anal. 2018, 150, 278–286. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Zhang, J.; Zhao, Y.L.; Li, T.; Shen, T.; Li, J.Q.; Li, W.Y.; Liu, H.G. Mycology, cultivation, traditional uses, phytochemistry and pharmacology of Wolfiporia cocos (Schwein.) Ryvarden et Gilb.: A review. J. Ethnopharmacol. 2013, 147, 265–276. [Google Scholar] [CrossRef]
- Liu, F.; Liu, Y.; Feng, X.; Ibrahim, S.A.; Huang, W. Structure characterization and in vitro immunomodulatory activities of carboxymethyl pachymaran. Int. J. Biol. Macromol. 2021, 178, 94–103. [Google Scholar] [CrossRef]
- Ke, R.D.; Lin, S.F.; Chen, Y.; Ji, C.R.; Shu, Q.G. Analysis of chemical composition of polysaccharides from Poria cocos Wolf and its anti-tumor activity by NMR spectroscopy. Carbohydr. Polym. 2010, 80, 31–34. [Google Scholar]
- Tang, J.; Nie, J.; Li, D.P.; Zhu, W.J.; Zhang, S.O.; Ma, F.; Sun, Q.; Song, J.; Zheng, Y.L.; Chen, P. Characterization and antioxidant activities of degraded polysaccharides from Poria cocos sclerotium. Carbohydr. Polym. 2014, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.H.; Guan, Z.Y.; Tang, N.; Cheng, Y.Q. The differences between the water- and alkaline-soluble Poria cocos polysaccharide: A review. Int. J. Biol. Macromol. 2023, 235, 123925. [Google Scholar] [CrossRef]
- Huang, S.Y.; Chen, F.; Cheng, H.; Huang, G.L. Modification and application of polysaccharide from traditional Chinese medicine such as Dendrobium officinale. Int. J. Biol. Macromol. 2020, 157, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Madruga, L.Y.C.; Sabino, R.M.; Santos, E.C.G.; Popat, K.C.; Balaban, R.C.; Kipper, M.J. Carboxymethyl-kappa-carrageenan: A study of biocompatibility, antioxidant and antibacterial activities. Int. J. Biol. Macromol. 2020, 152, 483–491. [Google Scholar] [CrossRef]
- Kou, F.; Ge, Y.F.; Wang, W.H.; Mei, Y.X.; Cao, L.K.; Wei, X.T.; Xiao, H.; Wu, X. A review of Ganoderma lucidum polysaccharides: Health benefit, structure-activity relationship, modification, and nanoparticle encapsulation. Int. J. Biol. Macromol. 2023, 243, 125199. [Google Scholar] [CrossRef]
- Xie, L.M.; Shen, M.Y.; Wang, Z.J.; Xie, J.H. Structure, function and food applications of carboxymethylated polysaccharides: A comprehensive review. Trends Food Sci. Technol. 2021, 118, 539–557. [Google Scholar] [CrossRef]
- Song, B.; Li, X.L.; Wu, Y.Z.; Shen, Y.; Deng, W.Y. Study on Preparation and Anti-tumor Activity of Carboxymethylpachymaran. Chin. J. Mod. Appl. Pharm. 2019, 36, 1328–1332. [Google Scholar]
- Liu, X.F.; Wang, X.Q.; Xu, X.F.; Zhang, X.W. Purification, antitumor and anti-inflammation activities of an alkali-soluble and carboxymethyl polysaccharide CMP33 from Poria cocos. Int. J. Biol. Macromol. 2019, 127, 39–47. [Google Scholar] [CrossRef]
- Wang, C.H.; Yang, S.X.; Gao, L.; Wang, L.L.; Cao, L. Carboxymethyl pachyman (CMP) reduces intestinal mucositis and regulates the intestinal microflora in 5-fluorouracil-treated CT26 tumour-bearing mice. Food Funct. 2018, 9, 2695–2704. [Google Scholar] [CrossRef]
- Wang, C.H.; Huo, X.W.; He, X.S.; Liu, D.Y.; Li, L.Y.; Cao, L. Effect of carboxymethyl Pachymaran on life extension and attenuation of cyclophosphamide-induced toxicity in CT26 tumor-bearing mice. Food Sci. 2016, 37, 229–233. [Google Scholar]
- Ishimoto, Y.; Ishibashi, K.I.; Yamanaka, D.; Adachi, Y.; Kanzaki, K.; Okita, K.; Iwakura, Y.; Ohno, N. Modulation of an innate immune response by soluble yeast beta-glucan prepared by a heat degradation method. Int. J. Biol. Macromol. 2017, 104, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.Q.; Liu, X.J.; Xiao, C.L. Study on the extraction technology of polysaccharide from purple corn by enzyme method. Sci. Technol. Food Ind. 2012, 33, 188–191. [Google Scholar]
- Morris, G.A.; Adams, G.G.; Harding, S.E. On hydrodynamic methods for the analysis of the sizes and shapes of polysaccharides in dilute solution: A short review. Food Hydrocoll. 2014, 42, 318–334. [Google Scholar] [CrossRef]
- Rajendar, B.; Reddy, M.J.; Madhavi, A.; Shravankumar, G.; Rao, G.S.; Matur, R.V. Multi angle light scattering as a process analytical technology tool for real-time monitoring of molar mass of protein-polysaccharide conjugate fractions. J. Chromatogr. 2022, 2, 100045. [Google Scholar] [CrossRef]
- Alvarez-Sabatel, S.; Marañón, I.M.d.; Arboleya, J.-C. Impact of high pressure homogenisation (HPH) on inulin gelling properties, stability and development during storage. Food Hydrocoll. 2015, 44, 333–344. [Google Scholar] [CrossRef]
- Li, Y.M.; Guo, X.X.; Zhong, R.F.; Ye, C.M.; Chen, J. Structure characterization and biological activities evaluation of two hetero-polysaccharides from Lepista nuda: Cell antioxidant, anticancer and immune-modulatory activities. Int. J. Biol. Macromol. 2023, 244, 125204. [Google Scholar] [CrossRef]
- Liu, M.D.; Dai, Y.D.; Song, C.M.; Wang, J.; Liu, Y.; Wang, Q. Structural Characterization of a Pleurotus sajor-caju Polysaccharide and Its Neuroprotection Related to the Inhibition of Oxidative Stress. Nutrients 2022, 14, 4047. [Google Scholar] [CrossRef]
- Chen, Z.X.; Zhao, Y.L.; Feng, X.; Zhang, L.J.; Ibrahim, S.A.; Huang, W.; Liu, Y. Effects of degradation on the physicochemical and antioxidant properties of carboxymethyl pachymaran. Int. J. Biol. Macromol. 2023, 245, 125560. [Google Scholar] [CrossRef]
- Yan, Q.J.; Han, L.J.; Jiang, Z.Q. Extraction of Astragalus polysaccharides by cellulase. Chin. Tradit. Herb. Drugs 2005, 36, 1804–1807. [Google Scholar]
- Hu, T.G.; Zou, Y.X.; Li, E.N.; Liao, S.T.; Wu, H.; Wen, P. Effects of enzymatic hydrolysis on the structural, rheological, and functional properties of mulberry leaf polysaccharide. Food Chem. 2021, 355, 129608. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.Y.; Zou, Y.; Guo, L.Q.; Lin, J.F.; Jiang, Z.; Zheng, Q.W. Rheological and microstructural properties of polysaccharide obtained from the gelatinous Tremella fuciformis fungus. Int. J. Biol. Macromol. 2023, 228, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.Y.; Zou, X.Y.; Zhu, B.Y.; You, L.J.; Zhao, Z.G.; Hileuskaya, K. The characteristics of polysaccharide from Gracilaria chouae and its application in food packaging with carboxymethyl cellulose and lysozyme. Food Hydrocoll. 2023, 135, 108109. [Google Scholar] [CrossRef]
- Xie, F.; Zhang, H.; Xia, Y.J.; Ai, L.Z. Effects of tamarind seed polysaccharide on gelatinization, rheological, and structural properties of corn starch with different amylose/amylopectin ratios. Food Hydrocoll. 2020, 105, 105854. [Google Scholar] [CrossRef]
- Vargas, M.; Perdones, Á.; Chiralt, A.; Cháfer, M.; González-Martínez, C. Effect of homogenization conditions on physicochemical properties of chitosan-based film-forming dispersions and films. Food Hydrocoll. 2011, 25, 1158–1164. [Google Scholar] [CrossRef]
- Ma, Y.Q.; Xiu, W.Y.; Wang, X.; Yu, S.Y.; Luo, Y.; Gu, X.Y. Structural characterization and in vitro antioxidant and hypoglycemic activities of degraded polysaccharides from sweet corncob. J. Cereal Sci. 2022, 108, 103579. [Google Scholar] [CrossRef]
- Peng, Y.; Song, X.L.; Shen, M.H. Inhibitory Effect of Hericium erinaceus Polysaccharide on Hepatoma-22(H22) Tumor Bearing Mice. Food Sci. 2012, 33, 244–246. [Google Scholar]
- Liu, X.H.; Guo, X.N.; Zhan, J.P. The effects of Polysaccharide from Gastrodia Elata B1 on cell cycle and caspase proteins activity in H22 tumor bearing mice. Chin. J. Gerontol. 2015, 35, 5681–5682. [Google Scholar]
- Cheng, Y.; Zhou, J.L.; Li, Q.; Liu, Y.; Wang, K.P.; Zhang, Y. The effects of polysaccharides from the root of Angelica sinensis on tumor growth and iron metabolism in H22-bearing mice. Food Funct. 2016, 7, 1033–1039. [Google Scholar] [CrossRef]
- Han, H.P.; Xie, H.C. A Study on the extraction and purification process of Lily Polysaccharide and its anti-tumor effect. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 485–489. [Google Scholar] [CrossRef]
- Feng, Y.R.; Liu, W.; Yang, J.G. Preparation of carboxymethylated pachyman with different molecular weight and study on its antioxidative activity. China Food Addit. 2019, 30, 67–74. [Google Scholar]
- Sheng, J.W.; Sun, Y.L. Antioxidant properties of different molecular weight polysaccharides from Athyrium multidentatum (Doll.) Ching. Carbohydr. Polym. 2014, 108, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.S.; Wang, X.M.; Liu, C.B.; Li, J.F. The degradation, antioxidant and antimutagenic activity of the mucilage polysaccharide from Dioscorea opposita. Carbohydr. Polym. 2016, 150, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.Q.; Wang, L.; Zhou, Y. Immunomodulation and antitumor activities of different-molecular-weight polysaccharides from Porphyridium cruentum. Carbohydr. Polym. 2012, 87, 1206–1210. [Google Scholar] [CrossRef]
- Seo, J.Y.; Lee, C.W.; Choi, D.J.; Lee, J.; Lee, J.Y.; Park, Y.I. Ginseng marc-derived low-molecular weight oligosaccharide inhibits the growth of skin melanoma cells via activation of RAW264.7 cells. Int. Immunopharmacol. 2015, 29, 344–353. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Yu, J.J.; Huang, J.; Liu, D.M.; Liang, M.H. Structural characterization of a novel Lactarius volemus Fr. polysaccharide and its immunity activity in BALB/c mice. RSC Adv. 2020, 10, 30254–30264. [Google Scholar] [CrossRef]
- Liu, C.; Dai, K.Y.; Ji, H.Y.; Jia, X.Y.; Liu, A.J. Structural characterization of a low molecular weight Bletilla striata polysaccharide and antitumor activity on H22 tumor-bearing mice. Int. J. Biol. Macromol. 2022, 205, 553–562. [Google Scholar] [CrossRef]
- Gao, Z.Z.; Zhang, C.; Jing, L.R.; Feng, M.; Li, R.; Yang, Y. The structural characterization and immune modulation activitives comparison of Codonopsis pilosula polysaccharide (CPPS) and selenizing CPPS (sCPPS) on mouse in vitro and vivo. Int. J. Biol. Macromol. 2020, 160, 814–822. [Google Scholar] [CrossRef]
- Wang, K.P.; Zhang, Q.L.; Liu, Y.; Wang, J.; Cheng, Y.; Zhang, Y. Structure and inducing tumor cell apoptosis activity of polysaccharides isolated from Lentinus edodes. J. Agric. Food Chem. 2013, 61, 9849–9858. [Google Scholar] [CrossRef]
- Su, S.Y.; Ding, X.; Hou, Y.L.; Liu, B.B.; Du, Z.H.; Liu, J.F. Structure elucidation, immunomodulatory activity, antitumor activity and its molecular mechanism of a novel polysaccharide from Boletus reticulatus Schaeff. Food Sci. Hum. Wellness 2023, 12, 647–661. [Google Scholar] [CrossRef]
- Dahlgren, D.; Lennernas, H. Review on the effect of chemotherapy on the intestinal barrier: Epithelial permeability, mucus and bacterial translocation. Biomed. Pharmacother. 2023, 162, 114644. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.T.; Cai, B.N.; Chen, H.; Wan, P.; Chen, D.K.; Ye, Z.Q.; Duan, A.L.; Chen, X.; Sun, H.L.; Pan, J.Y. Tuna trimmings (Thunnas albacares) hydrolysate alleviates immune stress and intestinal mucosal injury during chemotherapy on mice and identification of potentially active peptides. Curr. Res. Food Sci. 2023, 7, 100547. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.N.; Pan, J.Y.; Chen, H.; Chen, X.; Ye, Z.Q.; Yuan, H.B.; Sun, H.L.; Wan, P. Oyster polysaccharides ameliorate intestinal mucositis and improve metabolism in 5-fluorouracil-treated S180 tumour-bearing mice. Carbohydr. Polym. 2021, 256, 117545. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.W.; Zhang, M.; Yao, Y.; Zhou, X.F.; Liu, K.H. Interaction of polysaccharides, gut microbiota and immunity. Sci. Technol. Food Ind. 2017, 38, 315. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Mw (kDa) | Mn (kDa) | Mw/Mn | Rz (nm) |
|---|---|---|---|---|
| CMP | 706.4 ± 38.4 | 285.0 ± 28.4 | 2.479 ± 0.281 | 137.5 ± 2.2 |
| HTCMP | 429.8 ± 14.9 | 196.7 ± 10.2 | 2.186 ± 0.136 | 72.7 ± 2.1 |
| HTEC-24 | 129.9 ± 5.1 | 75.86 ± 3.61 | 1.713 ± 0.106 | 49.8 ± 3.9 |
| HTEC-48 | 68.62 ± 3.15 | 45.53 ± 3.82 | 1.507 ± 0.144 | 29.9 ± 5.9 |
| Sample | d (0.5) | D (3,2) (μm) | D (4,3) (μm) |
|---|---|---|---|
| CMP | 251.479 ± 3.459 a | 180.884 ± 0.918 a | 300.084 ± 9.227 a |
| HTCMP | 116.367 ± 4.444 b | 83.976 ± 3.007 b | 131.413 ± 5.317 b |
| HTEC-24 | 109.186 ± 7.333 b | 76.433 ± 3.470 c | 141.536 ± 13.866 b |
| HTEC-48 | 60.215 ± 2.159 c | 47.384 ± 0.805 d | 75.746 ± 7.878 c |
| Initial Weight | Final Weight | Weight Increment | |
|---|---|---|---|
| A | 31.52 ± 2.72 | 37.48 ± 3.67 | 5.96 ± 1.83 |
| B | 31.45 ± 4.01 | 38.85 ± 4.66 | 7.40 ± 1.63 |
| C | 31.59 ± 2.53 | 34.65 ± 2.27 * | 3.06 ± 1.89 #* |
| D1 | 31.24 ± 2.65 | 37.96 ± 2.87 | 6.71 ± 1.66 |
| D2 | 31.86 ± 1.90 | 38.72 ± 2.36 | 6.86 ± 1.88 |
| D3 | 31.86 ± 2.45 | 38.52 ± 2.57 | 6.66 ± 0.89 |
| E1 | 31.84 ± 1.93 | 38.19 ± 2.55 | 6.35 ± 2.03 |
| E2 | 31.70 ± 1.94 | 37.89 ± 3.01 | 6.19 ± 1.76 |
| E3 | 31.69 ± 2.16 | 39.43 ± 3.19 | 7.74 ± 1.98 |
| F1 | 31.54 ± 1.78 | 38.30 ± 3.01 | 6.76 ± 2.40 |
| F2 | 31.64 ± 1.51 | 38.48 ± 2.18 | 6.84 ± 2.32 |
| G1 | 31.55 ± 1.21 | 37.86 ± 1.68 | 6.31 ± 1.64 |
| G2 | 31.59 ± 1.74 | 39.51 ± 3.23 | 7.92 ± 1.75 |
| H | 30.92 ± 1.76 | 33.50 ± 2.47 #* | 2.58 ± 1.51 #* |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Feng, X.; Zhang, L.; Huang, W.; Liu, Y. Antitumor Activity of Carboxymethyl Pachymaran with Different Molecular Weights Based on Immunomodulatory and Gut Microbiota. Nutrients 2023, 15, 4527. https://doi.org/10.3390/nu15214527
Zhao Y, Feng X, Zhang L, Huang W, Liu Y. Antitumor Activity of Carboxymethyl Pachymaran with Different Molecular Weights Based on Immunomodulatory and Gut Microbiota. Nutrients. 2023; 15(21):4527. https://doi.org/10.3390/nu15214527
Chicago/Turabian StyleZhao, Yalin, Xi Feng, Lijia Zhang, Wen Huang, and Ying Liu. 2023. "Antitumor Activity of Carboxymethyl Pachymaran with Different Molecular Weights Based on Immunomodulatory and Gut Microbiota" Nutrients 15, no. 21: 4527. https://doi.org/10.3390/nu15214527
APA StyleZhao, Y., Feng, X., Zhang, L., Huang, W., & Liu, Y. (2023). Antitumor Activity of Carboxymethyl Pachymaran with Different Molecular Weights Based on Immunomodulatory and Gut Microbiota. Nutrients, 15(21), 4527. https://doi.org/10.3390/nu15214527