Influence of Prolonged Whole Egg Supplementation on Insulin-like Growth Factor 1 and Short-Chain Fatty Acids Product: Implications for Human Health and Gut Microbiota

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Participants and Intervention
2.3. Primary Outcome
2.3.1. Blood Test
2.3.2. Changes in IGF-1
2.3.3. Gut Microbiota Analysis
2.3.4. Short-Chain Fatty Acid Measurement
2.4. Dietary Assessment
2.5. Statistical Analysis
3. Results
3.1. Baseline Characteristics
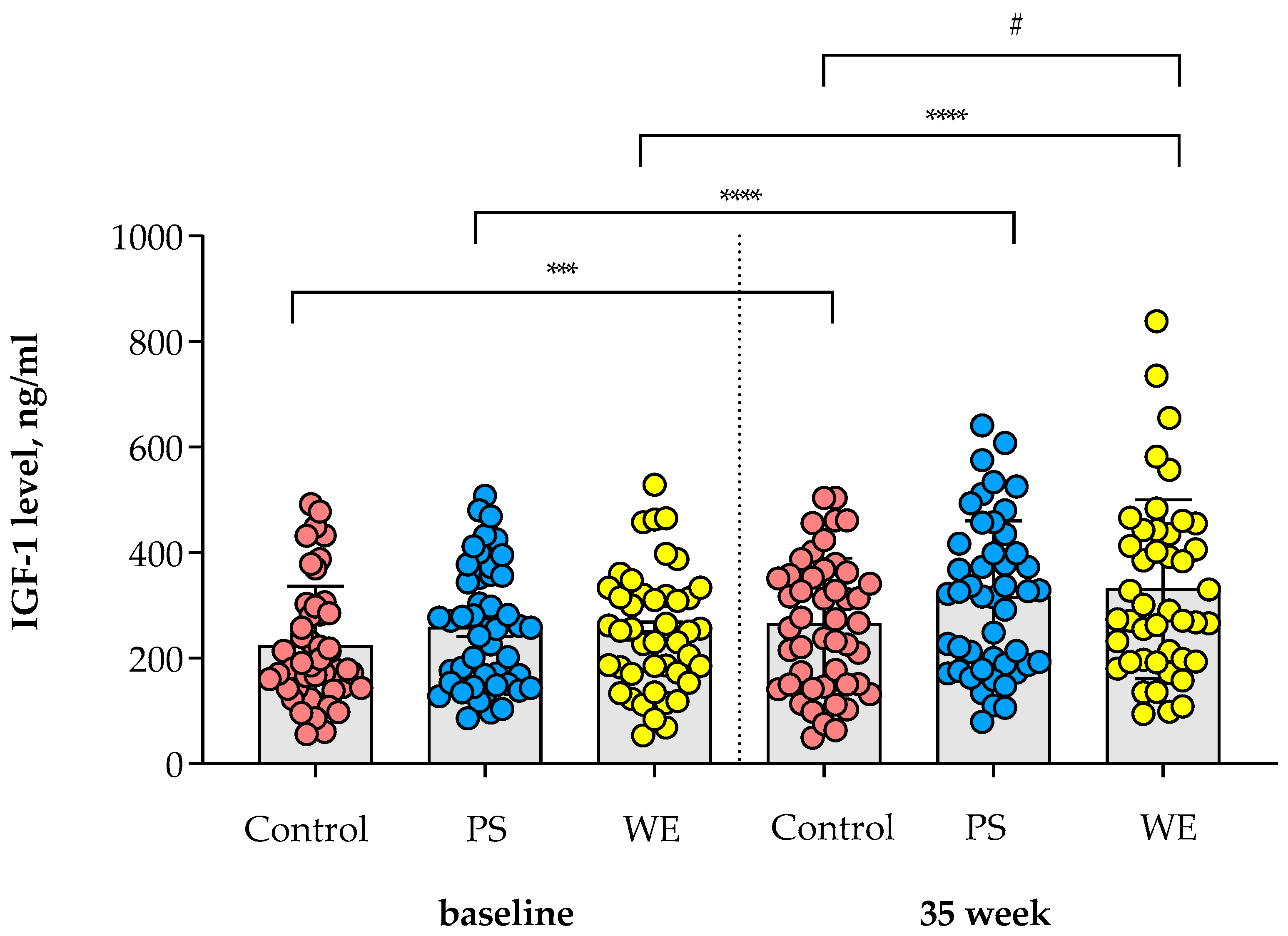
3.2. Change in IGF-1 Level
3.3. Change in SCFAs from Baseline to Endpoint
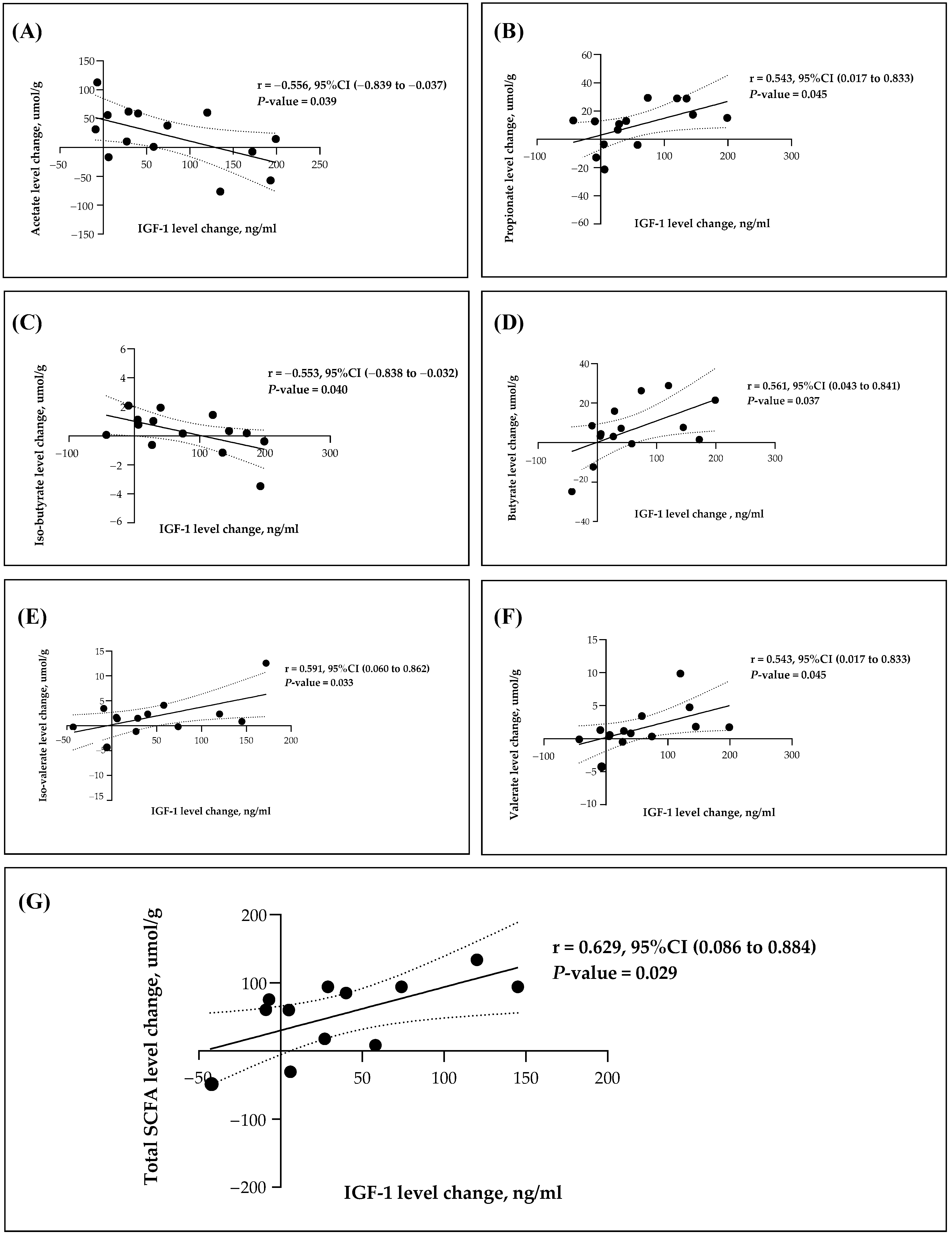
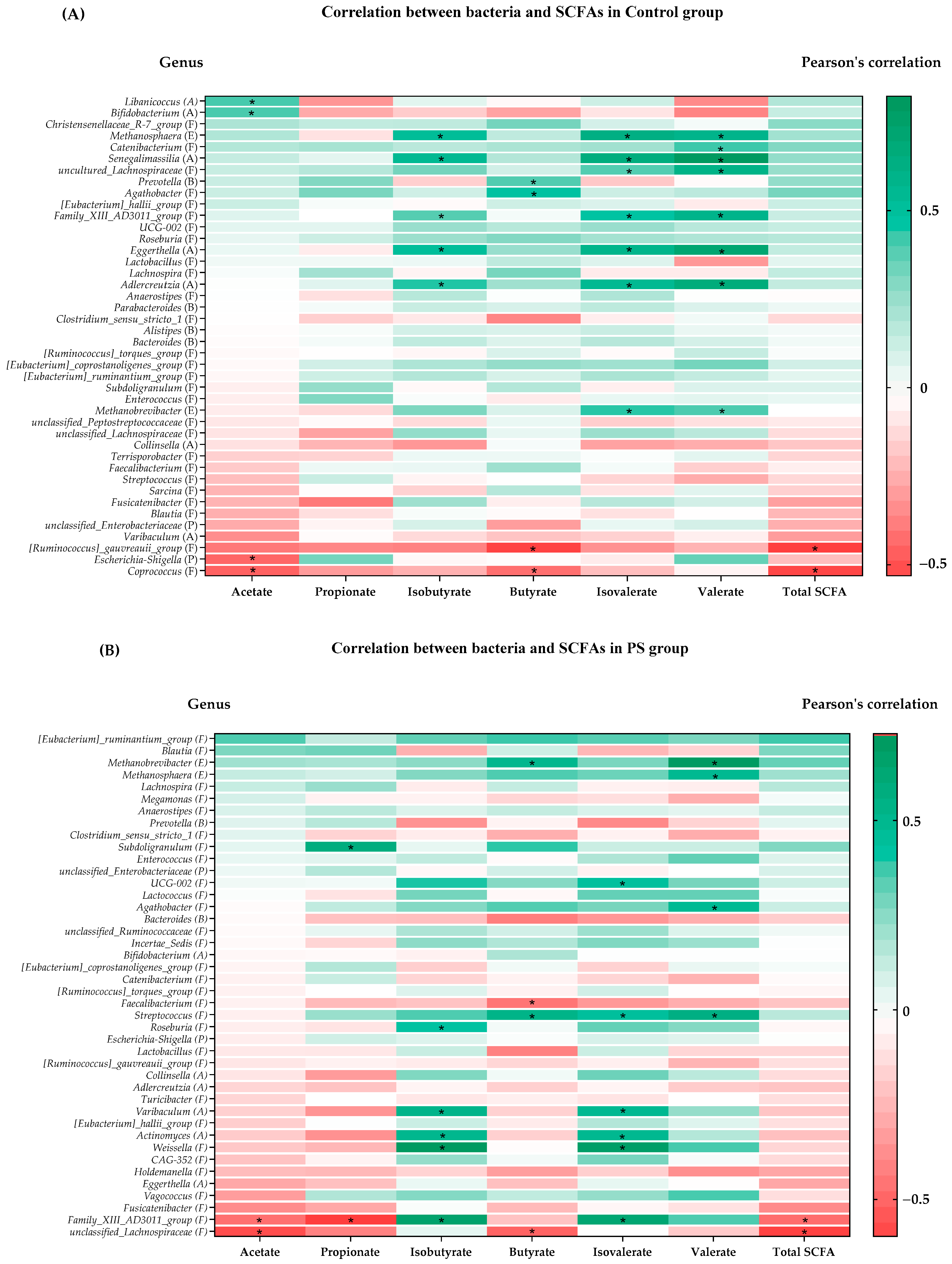
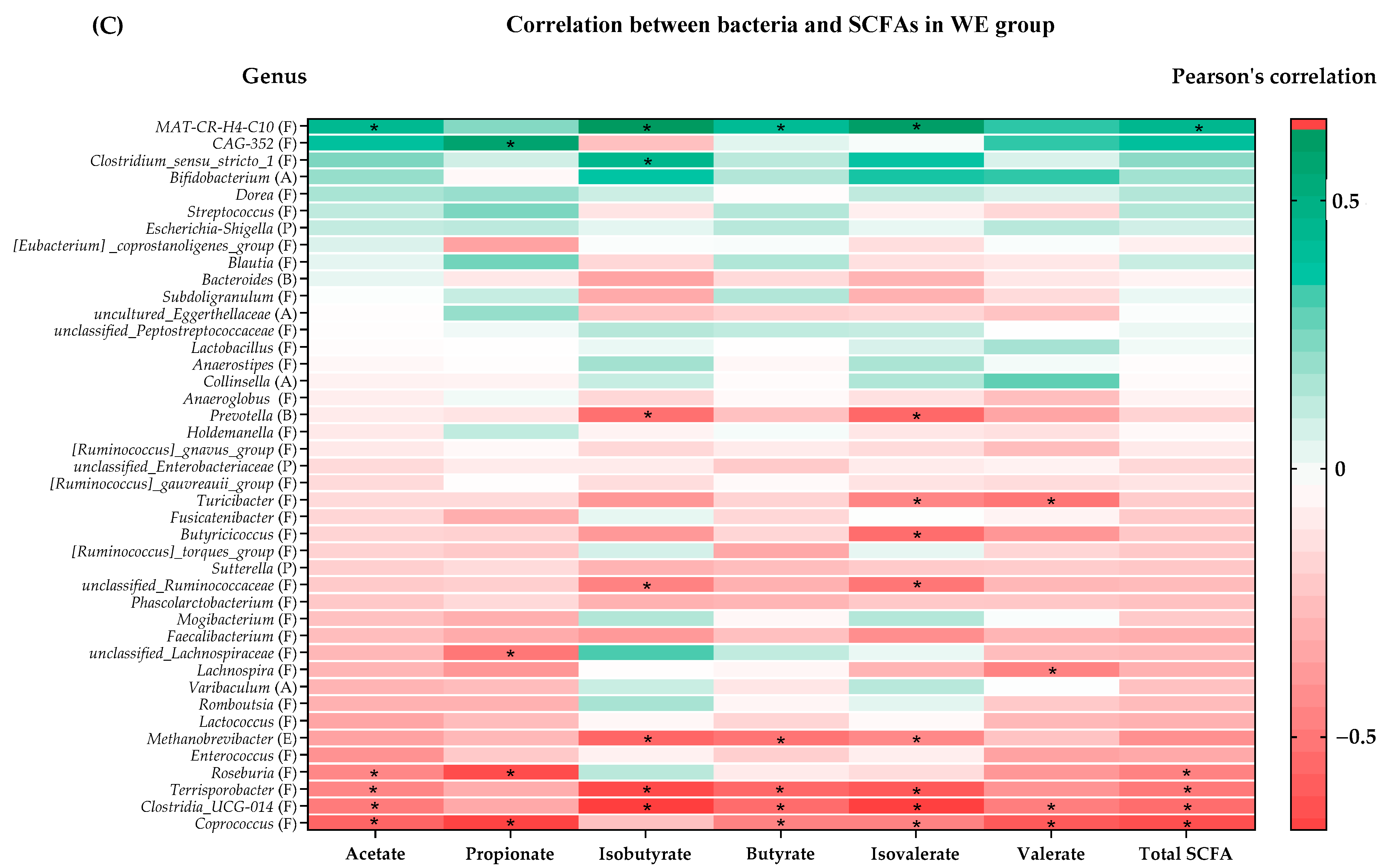
3.4. Association between SCFAs and Gut Microbiota
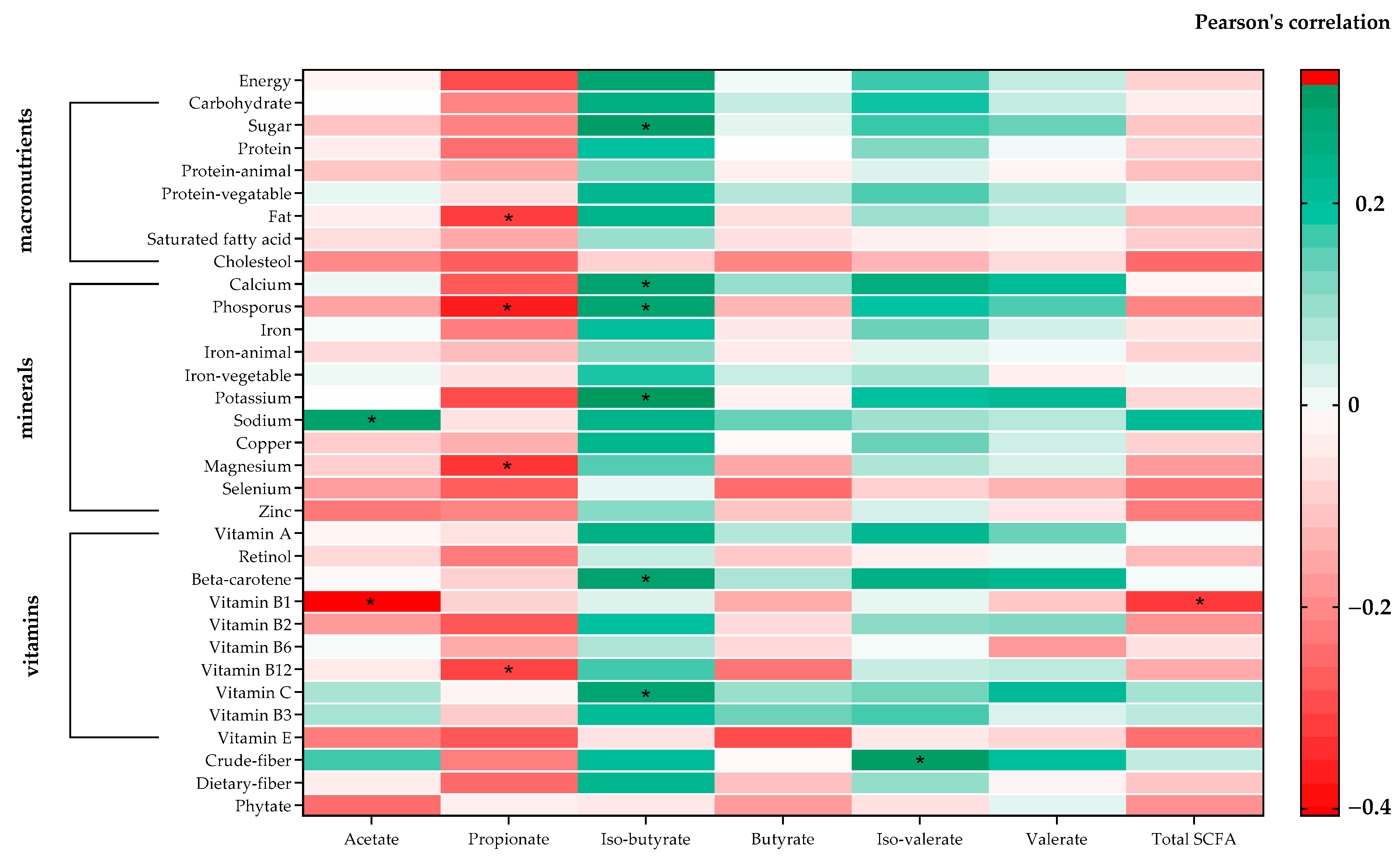
3.5. Association between SCFAs and Nutrition Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Levels and Trends in Child Malnutrition 2023. Available online: https://www.who.int/publications/i/item/9789240073791 (accessed on 21 September 2023).
- Réhault-Godbert, S.; Guyot, N.; Nys, Y. The Golden Egg: Nutritional Value, Bioactivities, and Emerging Benefits for Human Health. Nutrients 2019, 11, 684. [Google Scholar] [CrossRef] [PubMed]
- Suta, S.; Surawit, A.; Mongkolsucharitkul, P.; Pinsawas, B.; Manosan, T.; Ophakas, S.; Pongkunakorn, T.; Pumeiam, S.; Sranacharoenpong, K.; Sutheeworapong, S.; et al. Prolonged Egg Supplement Advances Growing Child’s Growth and Gut Microbiota. Nutrients 2023, 15, 1143. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Takakura, A.; Zandi-Nejad, K.; Charles, J.F. Mechanisms of gut microbiota-mediated bone remodeling. Gut Microbes 2018, 9, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, C.; Rovenna Udam, T.; Lauritzen, L.; Mølgaard, C.; Juul, A.; Fleischer Michaelsen, K. Animal protein intake, serum insulin-like growth factor I, and growth in healthy 2.5-y-old Danish children. Am. J. Clin. Nutr. 2004, 80, 447–452. [Google Scholar] [CrossRef]
- Socha, P.; Grote, V.; Gruszfeld, D.; Janas, R.; Demmelmair, H.; Closa-Monasterolo, R.; Subías, J.E.; Scaglioni, S.; Verduci, E.; Dain, E.; et al. Milk protein intake, the metabolic-endocrine response, and growth in infancy: Data from a randomized clinical trial. Am. J. Clin. Nutr. 2011, 94, S1776–S1784. [Google Scholar] [CrossRef]
- Michaelsen, K.F. Effect of Protein Intake from 6 to 24 Months on Insulin-Like Growth Factor 1 (IGF-1) Levels, Body Composition, Linear Growth Velocity, and Linear Growth Acceleration: What are the Implications for Stunting and Wasting? Food Nutr. Bull. 2013, 34, 268–271. [Google Scholar] [CrossRef]
- Romo Ventura, E.; Konigorski, S.; Rohrmann, S.; Schneider, H.; Stalla, G.K.; Pischon, T.; Linseisen, J.; Nimptsch, K. Association of dietary intake of milk and dairy products with blood concentrations of insulin-like growth factor 1 (IGF-1) in Bavarian adults. Eur. J. Nutr. 2020, 59, 1413–1420. [Google Scholar] [CrossRef]
- Lovell, A.L.; Milne, T.; Matsuyama, M.; Hill, R.J.; Davies, P.S.; Grant, C.C.; Wall, C.R. Protein Intake, IGF-1 Concentrations, and Growth in the Second Year of Life in Children Receiving Growing Up Milk–Lite (GUMLi) or Cow’s Milk (CM) Intervention. Front. Nutr. 2021, 8, 666228. [Google Scholar] [CrossRef]
- Van Coeverden, S.C.; Netelenbos, J.C.; De Ridder, C.M.; Roos, J.C.; Popp-Snijders, C.; Delemarre-van de Waal, H.A. Bone metabolism markers and bone mass in healthy pubertal boys and girls. Clin. Endocrinol. 2002, 57, 107–116. [Google Scholar] [CrossRef]
- Hua, C.; Yu, D. Correlation Study between Levels of Gastrin, Serum IGF-1, and GHBP and Growth and Development in Children with Short Stature Based on Big Data Analysis. Dis. Markers 2022, 2022, 4614099. [Google Scholar] [CrossRef]
- Wolters, M.; Ahrens, J.; Romaní-Pérez, M.; Watkins, C.; Sanz, Y.; Benítez-Páez, A.; Stanton, C.; Günther, K. Dietary fat, the gut microbiota, and metabolic health–A systematic review conducted within the MyNewGut project. Clin. Nutr. 2019, 38, 2504–2520. [Google Scholar] [CrossRef] [PubMed]
- Graf, D.; Di Cagno, R.; Fåk, F.; Flint, H.J.; Nyman, M.; Saarela, M.; Watzl, B. Contribution of diet to the composition of the human gut microbiota. Microb. Ecol. Health Dis. 2015, 26, 26164. [Google Scholar] [CrossRef]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Sellin, C. Short chain fatty acids in health and disease. Aliment. Pharmacol. Ther. 1998, 12, 499–507. [Google Scholar] [CrossRef]
- Hijova, E.; Chmelarova, A. Short chain fatty acids and colonic health. Bratisl. Lek. Listy 2007, 108, 354. [Google Scholar] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Rios-Covián, D.; Nogacka, A.M.; Saturio, S.; Gómez-Martín, M.; González, S.; de los Reyes-Gavilán, C.G.; Gueimonde, M.; Arboleya, S.; Salazar, N. Short-chain fatty acids and human nutrition. In Fats and Associated Compounds; Royal Society of Chemistry: London, UK, 2021; Chapter 11; pp. 254–285. [Google Scholar] [CrossRef]
- Blachier, F.; Beaumont, M.; Portune, K.J.; Steuer, N.; Lan, A.; Audebert, M.; Khodorova, N.; Andriamihaja, M.; Airinei, G.; Benamouzig, R.; et al. High-protein diets for weight management: Interactions with the intestinal microbiota and consequences for gut health. A position paper by the my new gut study group. Clin. Nutr. 2019, 38, 1012–1022. [Google Scholar] [CrossRef]
- Mu, C.; Yang, Y.; Luo, Z.; Guan, L.; Zhu, W. The colonic microbiome and epithelial transcriptome are altered in rats fed a high-protein diet compared with a normal-protein diet. J. Nutr. 2016, 146, 474–483. [Google Scholar] [CrossRef]
- Mu, C.; Yang, Y.; Luo, Z.; Zhu, W. Temporal microbiota changes of high-protein diet intake in a rat model. Anaerobe 2017, 47, 218–225. [Google Scholar] [CrossRef]
- Petchoo, J.; Kaewchutima, N.; Tangsuphoom, N. Nutritional quality of lunch meals and plate waste in school lunch programme in Southern Thailand. J. Nutr. Sci. 2022, 11, e35. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.S. Principles of Nutritional Assessment, 2nd ed.; Oxford University Press: Oxford, UK, 2005; pp. 41–46. [Google Scholar]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid. Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.; Ecklu-Mensah, G.; Maseng, M.G.; Donato, S.; Coo-Kang, C.; Dugas, L.; Bovet, P.; Bedu-Addo, K.; Plange-Rhule, J.; Forrester, T.; et al. Gut microbiota and fecal short chain fatty acids differ with adiposity and country of origin: The METS-Microbiome Study. Nat. Commun. 2023, 14, 5160. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and butyrate-producing colon bacteria: Importance and strategies for their stimulation in the human gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.D.; Pollak, M.N.; Willett, W.C.; Hankinson, S.E. Dietary correlates of plasma insulin-like growth factor I and insulin-like growth factor binding protein 3 concentrations. Cancer Epidemiol. Biomark. Prev. 2002, 11, 852–861. [Google Scholar]
- Rogers, I.S.; Gunnell, D.; Emmett, P.M.; Glynn, L.R.; Dunger, D.B.; Holly, J.M.; ALSPAC Study Team. Cross-sectional associations of diet and insulin-like growth factor levels in 7-to 8-year-old children. Cancer Epidemiol. Biomark. Prev. 2005, 14, 204–212. [Google Scholar] [CrossRef]
- Switkowski, K.M.; Jacques, P.F.; Must, A.; Fleisch, A.; Oken, E. Associations of protein intake in early childhood with body composition, height, and insulin-like growth factor I in mid-childhood and early adolescence. Am. J. Clin. 2019, 109, 1154–1163. [Google Scholar] [CrossRef]
- Yan, J.; Herzog, J.W.; Tsang, K.; Brennan, C.A.; Bower, M.A.; Garrett, W.S.; Sartor, B.R.; Aliprantis, A.O.; Charles, J.F. Gut microbiota induce IGF-1 and promote bone formation and growth. Proc. Nat. Acad. Sci. USA 2016, 113, e7554–e7563. [Google Scholar] [CrossRef]
- Li, L.; Chen, L.; Yang, Y.; Wang, J.; Guo, L.; An, J.; Ma, X.; Lu, W.; Xiao, Y.; Wang, X.; et al. Characteristics of Gut Microbiome and Its Metabolites, Short-Chain Fatty Acids, in Children with Idiopathic Short Stature. Front. Endocrinol. 2022, 13, 890200. [Google Scholar] [CrossRef]
- Wong, J.M.; De Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Di Gioia, D.; Aloisio, I.; Mazzola, G.; Biavati, B. Bifidobacteria: Their impact on gut microbiota composition and their applications as probiotics in infants. Appl. Microbiol. Biotechnol. 2014, 98, 563–577. [Google Scholar] [CrossRef] [PubMed]
- Grimm, V.; Westermann, C.; Riedel, C.U. Bifidobacteria-host interactions—An update on colonisation factors. BioMed Res. Int. 2014, 2014, 960826. [Google Scholar] [CrossRef] [PubMed]
- Laureys, D.; De Vuyst, L. Microbial species diversity, community dynamics, and metabolite kinetics of water kefir fermentation. Appl. Environ. Microbiol. 2014, 80, 2564–2572. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, M.; Savard, P.; Rivière, A.; LaPointe, G.; Roy, D. Bioaccessible antioxidants in milk fermented by Bifidobacterium longum subsp. Longum Strains. BioMed Res. Int. 2015, 2015, 169381. [Google Scholar] [CrossRef]
- Pascale, A.; Marchesi, N.; Marelli, C.; Coppola, A.; Luzi, L.; Govoni, S.; Giustina, A.; Gazzaruso, C. Microbiota and metabolic diseases. Endocrine 2018, 61, 357–371. [Google Scholar] [CrossRef]
- Dicks, L.M.; Geldenhuys, J.; Mikkelsen, L.S.; Brandsborg, E.; Marcotte, H. Our gut microbiota: A long walk to homeostasis. Benef. Microbes 2018, 9, 3–20. [Google Scholar] [CrossRef]
- Ryan, P.M.; Ross, R.P.; Fitzgerald, G.F.; Caplice, N.M.; Stanton, C. Sugar-coated: Exopolysaccharide producing lactic acid bacteria for food and human health applications. Food Funct. 2015, 6, 679–693. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef]
- Holen, T.; Norheim, F.; Gundersen, T.E.; Mitry, P.; Linseisen, J.; Iversen, P.O.; Drevon, C.A. Biomarkers for nutrient intake with focus on alternative sampling techniques. Genes Nutr. 2016, 11, 12. [Google Scholar] [CrossRef]
- La Fata, G.; Rastall, R.A.; Lacroix, C.; Harmsen, H.J.; Mohajeri, M.H.; Weber, P.; Steinert, R.E. Recent development of prebiotic research—Statement from an expert workshop. Nutrients 2017, 9, 1376. [Google Scholar] [CrossRef]
- Gratton, J.; Phetcharaburanin, J.; Mullish, B.H.; Williams, H.R.; Thursz, M.; Nicholson, J.K.; Holmes, E.; Marchesi, J.R.; Li, J.V. Optimized sample handling strategy for metabolic profiling of human feces. Anal. Chem. 2016, 88, 4661–4668. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Control [n = 26] | PS [n = 25] | WE [n = 24] | p-Value | |
|---|---|---|---|---|---|
| n (%) | n (%) | n (%) | |||
| Age, mean (SD), year | 9.7 (1.15) | 9.8 (0.93) | 9.7 (0.96) | 0.948 | |
| Sex | 0.841 | ||||
| Male | 12 (46.15) | 13 (52.00) | 13 (54.17) | ||
| Female | 14 (53.85) | 12 (48.00) | 11 (45.83) | ||
| Weight, mean (SD), kg | 37.1 (16.23) | 32.3 (10.56) | 32.5 (8.03) | 0.298 | |
| Height, mean (SD), cm | 139.3 (9.21) | 138.5 (11.20) | 138.7 (7.92) | 0.958 | |
| Obesity status | 0.829 | ||||
| Underweight | 3 (11.5) | 3 (12.0) | 3 (12.5) | ||
| Normal | 15 (57.6) | 18 (72.0) | 17 (70.8) | ||
| Overweight | 3 (11.5) | 2 (8.0) | 1 (4.2) | ||
| Obese | 5 (19.2) | 2 (8.0) | 3 (12.5) | ||
| Height status | 0.386 | ||||
| Stunted | 3 (11.5) | 4 (16.0) | 2 (8.3) | ||
| Normal | 20 (76.9) | 18 (72.0) | 21 (87.5) | ||
| Tall stature | 3 (11.5) | 3 (12.0) | 1 (4.2) | ||
| Blood pressure, mean (SD), mm Hg | |||||
| Systolic | 102.3 (10.93) | 104.0 (11.42) | 98.7 (7.35) | 0.178 | |
| Diastolic | 69.1 (6.14) | 71.2 (5.80) | 69.7 (6.16) | 0.447 | |
| Fasting blood sugar, mean (SD), mmol/L | 88.6 (10.34) | 86.4 (7.58) | 87.9 (8.95) | 0.674 | |
| Transferrin, mean (SD), g/L | 256.6 (20.50) | 264.1 (29.98) | 262.8 (33.29) | 0.601 | |
| Prealbumin, mean (SD), μmol/L | 0.2 (0.03) | 0.2 (0.04) | 0.2 (0.04) | 0.314 | |
| Albumin, mean (SD), g/L | 4.4 (0.24) | 4.4 (0.23) | 4.4 (0.25) | 0.579 | |
| Blood lipid level, mean (SD), mmol/L | |||||
| TC | 176.2 (30.24) | 176.2 (31.97) | 185 (31.00) | 0.521 | |
| TG | 84.9 (23.24) | 80.2 (35.86) | 82.9 (30.10) | 0.855 | |
| HDL-C | 55.5 (11.61) | 57.8 (9.84) | 57.5 (10.05) | 0.710 | |
| LDL-C | 103.6 (25.60) | 102.3 (27.53) | 110.8 (25.61) | 0.477 | |
| Nutrition intake, mean (SD) | |||||
| Total energy intake, kcal/day | 951.1 (233.95) | 863.9 (263.33) | 749.7 (268.36) | 0.110 | |
| Protein intake, g/day | 37.8 (9.08) | 39.9 (12.98) | 35.0 (9.50) | 0.455 | |
| Carbohydrate, g/day | 99.2 (22.58) | 92.9 (41.17) | 86.1 (42.34) | 0.287 | |
| Fat, g/day | 34.6 (11.34) | 36.9 (15.76) | 29.5 (9.75) | 0.262 | |
| Saturated fatty acid, g/day | 7.7 (3.45) | 8.3 (2.69) | 7.1 (3.44) | 0.127 | |
| Cholesterol, mg/day | 182.3 (90.72) b | 209.8 (116.96) b | 302.6 (115.90) a | 0.011 | |
| Dietary fiber, g/day | 3.6 (1.87) | 3.3 (1.97) | 2.8 (1.43) | 0.454 | |
| Parameters | Control [n = 26] | PS [n = 25] | WE [n = 24] | p-Value | |
|---|---|---|---|---|---|
| Mean ± SEM | Mean ± SEM | Mean ± SEM | |||
| Acetate, μmol/g | |||||
| baseline | 75.16 ± 8.63 | 84.03 ± 8.63 | 72.98 ± 8.63 | 0.633 | |
| week 35 | 88.21 ± 10.08 | 97.67 ± 10.08 | 75.41 ± 10.08 | 0.299 | |
| p-value | 0.135 | 0.266 | 0.771 | ||
| Propionate, μmol/g | |||||
| baseline | 27.83 ± 3.57 | 31.11 ± 3.64 | 28.01 ± 3.57 | 0.732 | |
| week 35 | 29.73 ± 3.93 ab | 39.21 ± 4.01 a | 24.39 ± 3.93 b | 0.033 | |
| p-value | 0.644 | 0.102 | 0.385 | ||
| Isobutyrate, μmol/g | |||||
| baseline | 2.58 ± 0.46 | 2.62 ± 0.44 | 2.74 ± 0.46 | 0.931 | |
| week 35 | 2.17 ± 0.35 | 2.54 ± 0.33 | 2.68 ± 0.35 | 0.482 | |
| p-value | 0.297 | 0.845 | 0.891 | ||
| Butyrate, μmol/g | |||||
| baseline | 21.86 ± 2.17 | 21.96 ± 2.17 | 20.30 ± 2.17 | 0.832 | |
| week 35 | 21.13 ± 2.29 | 22.04 ± 2.29 | 18.62 ± 2.29 | 0.551 | |
| p-value | 0.752 | 0.976 | 0.602 | ||
| Isovalerate, μmol/g | |||||
| baseline | 3.92 ± 0.68 | 4.36 ± 0.70 | 4.01 ± 0.70 | 0.894 | |
| week 35 | 3.12 ± 0.55 | 3.50 ± 0.56 | 4.80 ± 0.56 | 0.082 | |
| p-value | 0.198 | 0.250 | 0.388 | ||
| Valerate, μmol/g | |||||
| baseline | 3.09 ± 0.50 | 2.62 ± 0.52 | 2.54 ± 0.51 | 0.704 | |
| week 35 | 2.49 ± 0.38 | 2.97 ± 0.39 | 2.73 ± 0.39 | 0.674 | |
| p-value | 0.384 | 0.556 | 0.525 | ||
| Hexanoic acid, μmol/g | |||||
| baseline | 0.46 ± 0.17 | 0.28 ± 0.18 | 0.65 ± 0.15 | 0.948 | |
| week 35 | 0.36 ± 1.11 | 0.30 ± 1.20 | 2.15 ± 0.98 | 0.426 | |
| p-value | 0.449 | 0.900 | 0.375 | ||
| Phenol, μmol/g | |||||
| baseline | 0.22 ± 0.36 | 0.82 ± 0.36 | 0.19 ± 0.36 | 0.757 | |
| week 35 | 0.19 ± 2.20 | 0.57 ± 2.20 | 3.92 ± 2.20 | 0.681 | |
| p-value | 0.192 | 0.649 | 0.499 | ||
| p-cresol, μmol/g | |||||
| baseline | 0.66 ± 0.15 ab | 0.97 ± 0.14 a | 0.44 ± 0.15 b | 0.022 | |
| week 35 | 0.86 ± 0.16 | 0.91 ± 0.16 | 0.73 ± 0.17 | 0.752 | |
| p-value | 0.108 | 0.564 | 0.017 | ||
| Total SCFA, μmol/g | |||||
| baseline | 135.11 ± 13.18 | 148.03 ± 13.18 | 130.67 ± 13.18 | 0.628 | |
| week 35 | 147.79 ± 14.88 | 167.37 ± 14.88 | 129.88 ± 14.88 | 0.212 | |
| p-value | 0.289 | 0.275 | 0.958 | ||
| Groups | SCFA Levels Change | Bacterial Genera (Phylum) | Bacterial Genera Producing SCFAs [% Change in Relative Abundance from Baseline] | Significant Correlation of SCFAs (p < 0.05) |
|---|---|---|---|---|
| WE | ↑ acetate ↓ propionate | Firmicutes | ↓ MAT-CR-H4-C10 | (+) acetate, isobutyrate, butyrate, iso-valerate, total SCFAs |
| ↓ butyrate | ↓ CAG-352 | (+) propionate | ||
| Stability of total SCFAs | ↓ Clostridium_sensu_stricto_1 | (+) isobutyrate | ||
| ↓ Butyricicoccus | (−) isovalerate | |||
| ↓ unclassified_Ruminococcaceae | (−) isobutyrate, isovalerate | |||
| ↓ unclassified_Lachnospiraceae | (−) propionate | |||
| ↓ Lachnospira | (−) valerate | |||
| ↑ Turicibacter | (−) isovalerate, valerate | |||
| ↑ Roseburia | (−) acetate, propionate, total SCFAs | |||
| ↑ Terrisporobacter | (−) acetate, isobutyrate, butyrate, isovalerate, total SCFAs | |||
| ↑ Clostridia_UCG_014 | (−) acetate, isobutyrate, butyrate, isovalerate, valerate, total SCFAs | |||
| ↑ Coprococcus | (−) acetate, propionate, butyrate, isovalerate, valerate, total SCFAs | |||
| Bacteroidota | ↓ Prevotella | (−) isobutyrate, isovalerate | ||
| Euryarchaeota | ↑ Methanobrevibacter | (−) isobutyrate, butyrate, isovalerate | ||
| PS | ↑ acetate | Firmicutes | ↓ Streptococcus | (+) butyrate, isovalerate, valerate |
| ↑ propionate | ↓ Weissella | (+) isobutyrate, isovalerate | ||
| ↑ butyrate ↑ total SCFA | ↓ Family_XIII_AD3011_group | (+) isobutyrate, isovalerate (−) acetate, propionate, total SCFAs | ||
| ↑ Subdoligranulum | (+) propionate | |||
| ↑ UCG-002 | (+) isovalerate | |||
| ↑ Agathobacter | (+) valerate | |||
| ↑ Faecalibacterium | (−) butyrate | |||
| ↑ Roseburia | (+) isobutyrate | |||
| ↑ unclassified_Lachnospiraceae | (−) acetate, butyrate, total SCFAs | |||
| Actinobacteriota | ↓ Varibaculum | (+) isobutyrate, isovalerate | ||
| ↓ Actinomyces | (+) isobutyrate, isovalerate | |||
| Euryarchaeota | ↑ Methanobrevibacter | (+) butyrate, valerate | ||
| ↑ Methanosphaera | (+) valerate | |||
| Control | ↑ acetate | Firmicutes | ↓ Catenibacterium | (+) valerate |
| ↑propionate | ↓ uncultured_Lachnospiraceae | (+) isovalerate, valerate | ||
| Stability of butyrate | ↓ Family_XIII_AD3011_group | (+) isobutyrate, isovalerate, valerate | ||
| ↑ total SCFA | ↓ [Ruminococcus]_gauvreauii_group | (−) butyrate, total SCFAs | ||
| ↑ Agathobacter | (+) butyrate | |||
| ↑ Coprococcus | (+) acetate, butyrate, total SCFAs | |||
| Control | Actinobacteriota | ↓ Libanicoccus | (+) acetate | |
| ↓ Bifidobacterium | (+) acetate | |||
| ↓ Senegalimassilia | (+) isobutyrate, isovalerate, valerate | |||
| ↓ Eggerthella | (+) isobutyrate, isovalerate, valerate | |||
| ↓ Adlercreutzia | (+) isobutyrate, isovalerate, valerate | |||
| Euryarchaeota | ↓ Methanosphaera | (+) isobutyrate, isovalerate, valerate | ||
| ↓ Methanobrevibacter | (+) isovalerate, valerate | |||
| Bacteroidota | ↑ Prevotella | (+) butyrate | ||
| Proteobacteria | ↑ Escherichia-Shigella | (−) acetate |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suta, S.; Ophakas, S.; Manosan, T.; Honwichit, O.; Charoensiddhi, S.; Surawit, A.; Pongkunakorn, T.; Pumeiam, S.; Mongkolsucharitkul, P.; Pinsawas, B.; et al. Influence of Prolonged Whole Egg Supplementation on Insulin-like Growth Factor 1 and Short-Chain Fatty Acids Product: Implications for Human Health and Gut Microbiota. Nutrients 2023, 15, 4804. https://doi.org/10.3390/nu15224804
Suta S, Ophakas S, Manosan T, Honwichit O, Charoensiddhi S, Surawit A, Pongkunakorn T, Pumeiam S, Mongkolsucharitkul P, Pinsawas B, et al. Influence of Prolonged Whole Egg Supplementation on Insulin-like Growth Factor 1 and Short-Chain Fatty Acids Product: Implications for Human Health and Gut Microbiota. Nutrients. 2023; 15(22):4804. https://doi.org/10.3390/nu15224804
Chicago/Turabian StyleSuta, Sophida, Suphawan Ophakas, Thamonwan Manosan, Orranich Honwichit, Suvimol Charoensiddhi, Apinya Surawit, Tanyaporn Pongkunakorn, Sureeporn Pumeiam, Pichanun Mongkolsucharitkul, Bonggochpass Pinsawas, and et al. 2023. "Influence of Prolonged Whole Egg Supplementation on Insulin-like Growth Factor 1 and Short-Chain Fatty Acids Product: Implications for Human Health and Gut Microbiota" Nutrients 15, no. 22: 4804. https://doi.org/10.3390/nu15224804
APA StyleSuta, S., Ophakas, S., Manosan, T., Honwichit, O., Charoensiddhi, S., Surawit, A., Pongkunakorn, T., Pumeiam, S., Mongkolsucharitkul, P., Pinsawas, B., Sutheeworapong, S., Puangsombat, P., Khoomrung, S., & Mayurasakorn, K. (2023). Influence of Prolonged Whole Egg Supplementation on Insulin-like Growth Factor 1 and Short-Chain Fatty Acids Product: Implications for Human Health and Gut Microbiota. Nutrients, 15(22), 4804. https://doi.org/10.3390/nu15224804