
Bifidobacterium: Host–Microbiome Interaction and Mechanism of Action in Preventing Common Gut-Microbiota-Associated Complications in Preterm Infants: A Narrative Review


Abstract
:1. Introduction
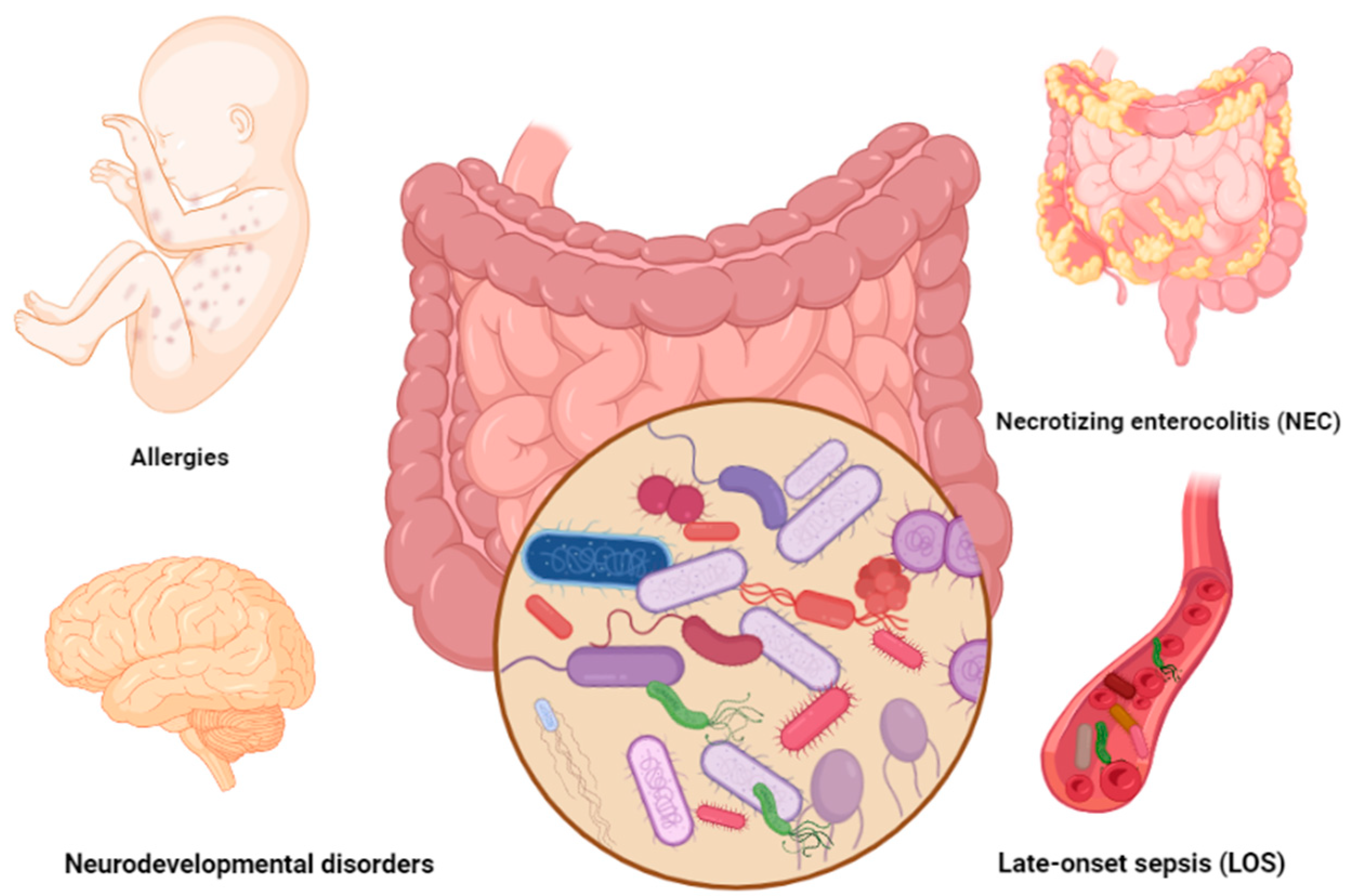
2. Common Gut-Microbiota-Associated Complications in Preterm Infants
2.1. Gastrointestinal Disorders
2.1.1. Necrotizing Enterocolitis (NEC)
2.1.2. Late-Onset Sepsis (LOS)
2.2. Allergies
2.3. Neurodevelopmental Diseases
2.3.1. Attention Deficit Hyperactivity Disorder
2.3.2. Schizophrenia Spectrum Disorder
2.3.3. Autism Spectrum Disorder
3. General Characteristics of Bifidobacterium
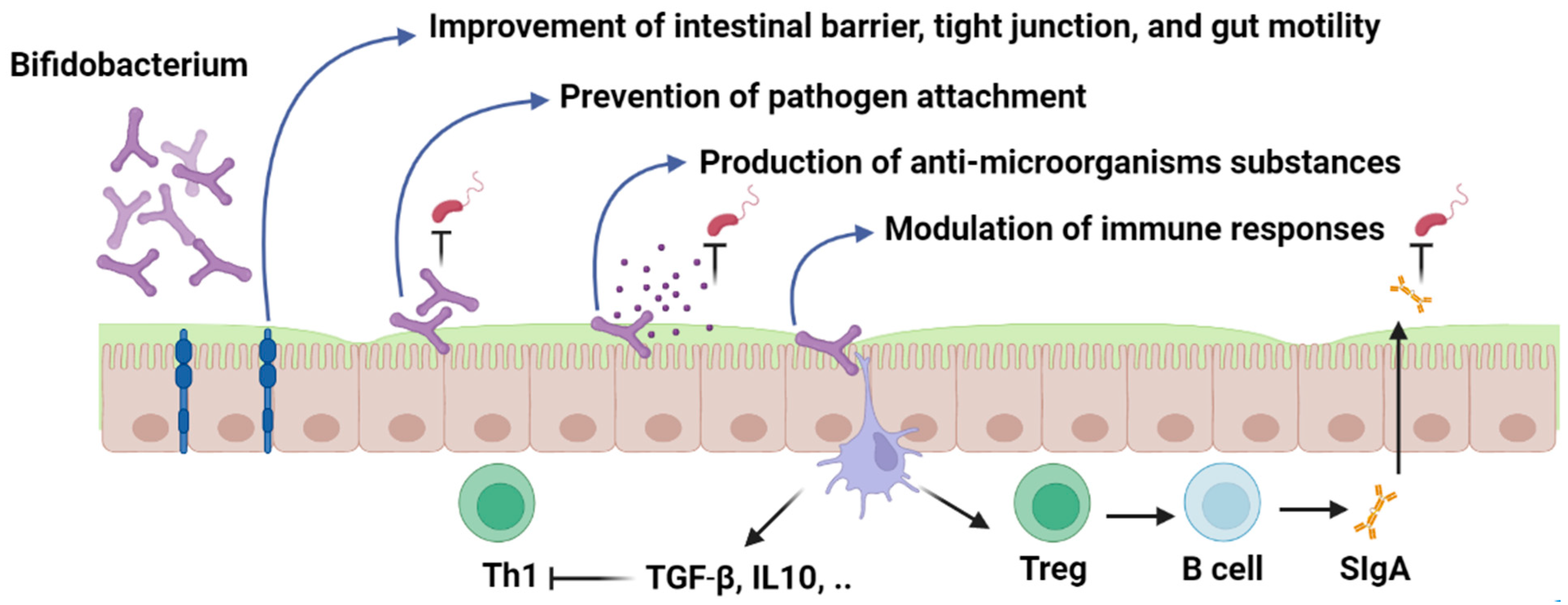
4. Immunomodulatory Effects of Bifidobacterium
5. Bifidobacterium as Probiotic
5.1. Bifidobacterium and Prevention of NEC and LOS
5.2. Bifidobacterium and Prevention of Neurodevelopmental Diseases
6. Bifidobacterium: Mechanism of Action
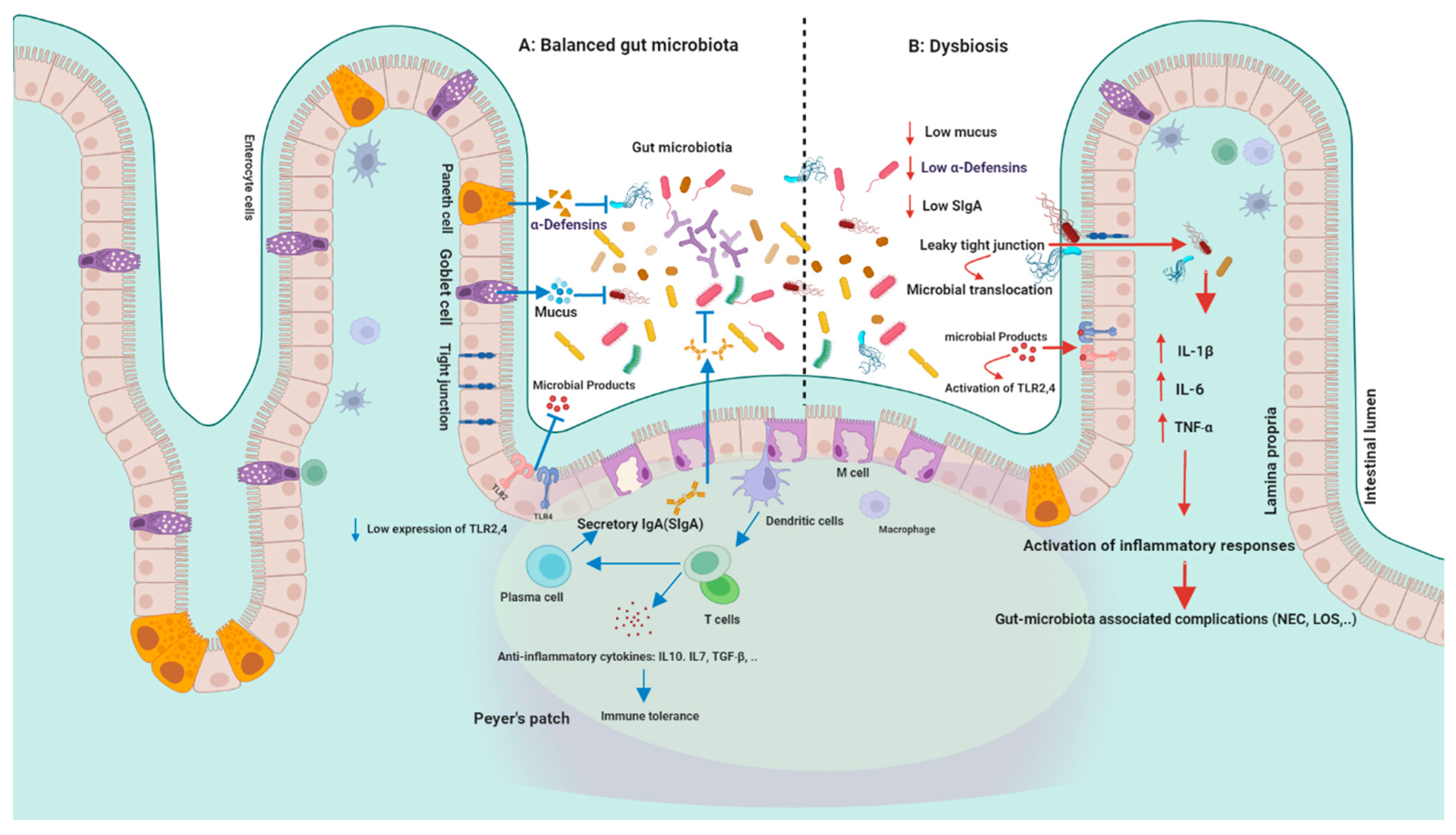
6.1. Modulation of the Immune System
6.2. Intracellular Immune Receptors (TLRs, NLRs) and Anti-Inflammatory Mediators
6.3. Regulation of Intestinal Epithelium Function
6.4. Competitive Exclusion and Adhesion Properties
6.5. Synthesis of Antimicrobial Compounds
7. Safety of Bifidobacterium Probiotic
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TLR | toll-like receptor |
| FOS | Fructooligosaccharides |
| GOS | galactooligosaccharides |
| NEC | Necrotizing enterocolitis |
| NOD2 | nucleotide-binding oligomerization domain-containing 2 |
| LOS | Late-onset sepsis |
| AD | atopic dermatitis |
| ADHD | Attention deficit hyperactivity disorder |
| ASD | autism spectrum disorders |
| HMOs | human milk oligosaccharides |
| GALT | gut-associated lymphoid tissue |
| DC | dendritic cells |
| sIgA | Secretory immunoglobulin A |
| IECs | intestinal epithelial cells |
| PRRs | pattern-recognition receptors |
| CLRs | C-type lectin receptors |
| RLR | RIG-I-like receptor |
| AIM2 | Absent in melanoma 2 like receptors |
| OAS | oligoadenylate synthase receptor |
| AMPs | antimicrobial peptides |
| TGF β | transforming growth factor-β |
| IL | interleukin |
| IFN-γ | interferon-γ |
| Ig | immunoglobulin |
| TNF | tumor necrosis factor |
| QPS | Qualified Presumption of Safety |
| EFSA | European Food Safety Authority |
| FDA | Food and Drug Administration |
| PAMPs | pathogen-associated molecular patterns |
| TJs | tight junctions |
| LMWs | low molecular weight compounds |
References
- Kumbhare, S.V.; Patangia, D.V.; Patil, R.H.; Shouche, Y.S.; Patil, N.P. Factors influencing the gut microbiome in children: From infancy to childhood. J. Biosci. 2019, 44, 49. [Google Scholar] [CrossRef] [PubMed]
- Kapourchali, F.R.; Cresci, G.A. Early-Life Gut Microbiome—The Importance of Maternal and Infant Factors in Its Establishment. Nutr. Clin. Pract. 2020, 35, 386–405. [Google Scholar] [CrossRef] [PubMed]
- Walker, W. Dysbiosis. In The Microbiota in Gastrointestinal Pathophysiology; Elsevier: Amsterdam, The Netherlands, 2017; pp. 227–232. [Google Scholar]
- WHO. Available online: https://www.who.int/news-room/fact-sheets/detail/preterm-birth (accessed on 15 August 2022).
- Schmutz, N.; Henry, E.; Jopling, J.; Christensen, R. Expected ranges for blood neutrophil concentrations of neonates: The Manroe and Mouzinho charts revisited. J. Perinatol. 2008, 28, 275–281. [Google Scholar] [CrossRef]
- Li, J.; Li, H.; Mao, H.; Yu, M.; Yang, F.; Feng, T.; Fan, Y.; Lu, Q.; Shen, C.; Yin, Z. Impaired NK cell antiviral cytokine response against influenza virus in small-for-gestational-age neonates. Cell. Mol. Immunol. 2013, 10, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Pammi, M.; Cope, J.; Tarr, P.I.; Warner, B.B.; Morrow, A.L.; Mai, V.; Gregory, K.E.; Kroll, J.S.; McMurtry, V.; Ferris, M.J. Intestinal dysbiosis in preterm infants preceding necrotizing enterocolitis: A systematic review and meta-analysis. Microbiome 2017, 5, 31. [Google Scholar] [CrossRef] [Green Version]
- Pietzak, M. Bacterial colonization of the neonatal gut. J. Craniofacial Surg. 2004, 38, 389–391. [Google Scholar] [CrossRef]
- Carbone, F.; Montecucco, F.; Sahebkar, A. Current and emerging treatments for neonatal sepsis. Expert Opin. Pharmacother. 2020, 21, 549–556. [Google Scholar] [CrossRef]
- Harmsen, H.J.; Wildeboer–Veloo, A.C.; Raangs, G.C.; Wagendorp, A.A.; Klijn, N.; Bindels, J.G.; Welling, G.W. Analysis of intestinal flora development in breast-fed and formula-fed infants by using molecular identification and detection methods. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 61–67. [Google Scholar] [CrossRef]
- Tannock, G.W. Analysis of bifidobacterial populations in bowel ecology studies. Bifidobacteria. Genom. Mol. Asp. 2010, 1–15. [Google Scholar]
- Avershina, E.; Storrø, O.; Øien, T.; Johnsen, R.; Wilson, R.; Egeland, T.; Rudi, K. Bifidobacterial succession and correlation networks in a large unselected cohort of mothers and their children. Appl. Environ. Microbiol. 2013, 79, 497–507. [Google Scholar] [CrossRef] [Green Version]
- Masi, A.C.; Embleton, N.D.; Lamb, C.A.; Young, G.; Granger, C.L.; Najera, J.; Smith, D.P.; Hoffman, K.L.; Petrosino, J.F.; Bode, L. Human milk oligosaccharide DSLNT and gut microbiome in preterm infants predicts necrotising enterocolitis. Gut 2021, 70, 2273–2282. [Google Scholar] [CrossRef] [PubMed]
- Kliegman, R.; Fanaroff, A. Necrotizing enterocolitis. New Engl. J. Med. 1984, 310, 1093–1103. [Google Scholar] [CrossRef] [PubMed]
- Alsaied, A.; Islam, N.; Thalib, L. Global incidence of necrotizing enterocolitis: A systematic review and meta-analysis. BMC Pediatr. 2020, 20, 344. [Google Scholar] [CrossRef] [PubMed]
- Hackam, D.J. Necrotizing Enterocolitis: Pathogenesis, Diagnosis and Treatment; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Brown, C.T.; Xiong, W.; Olm, M.R.; Thomas, B.C.; Baker, R.; Firek, B.; Morowitz, M.J.; Hettich, R.L.; Banfield, J.F. Hospitalized premature infants are colonized by related bacterial strains with distinct proteomic profiles. MBio 2018, 9, e00441-18. [Google Scholar] [CrossRef] [Green Version]
- Kaelin, E.A.; Rodriguez, C.; Hall-Moore, C.; Hoffmann, J.A.; Linneman, L.A.; Ndao, I.M.; Warner, B.B.; Tarr, P.I.; Holtz, L.R.; Lim, E.S. Longitudinal gut virome analysis identifies specific viral signatures that precede necrotizing enterocolitis onset in preterm infants. Nat. Microbiol. 2022, 7, 653–662. [Google Scholar] [CrossRef]
- Fundora, J.B.; Guha, P.; Shores, D.R.; Pammi, M.; Maheshwari, A. Intestinal dysbiosis and necrotizing enterocolitis: Assessment for causality using Bradford Hill criteria. Pediatr. Res. 2020, 87, 235–248. [Google Scholar] [CrossRef]
- Jilling, T.; Simon, D.; Lu, J.; Meng, F.J.; Li, D.; Schy, R.; Thomson, R.B.; Soliman, A.; Arditi, M.; Caplan, M.S. The roles of bacteria and TLR4 in rat and murine models of necrotizing enterocolitis. J. Immunol. 2006, 177, 3273–3282. [Google Scholar] [CrossRef] [Green Version]
- Leaphart, C.L.; Cavallo, J.; Gribar, S.C.; Cetin, S.; Li, J.; Branca, M.F.; Dubowski, T.D.; Sodhi, C.P.; Hackam, D.J. A critical role for TLR4 in the pathogenesis of necrotizing enterocolitis by modulating intestinal injury and repair. J. Immunol. 2007, 179, 4808–4820. [Google Scholar] [CrossRef] [Green Version]
- Hackam, D.J.; Sodhi, C.P. Bench to bedside—New insights into the pathogenesis of necrotizing enterocolitis. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 468–479. [Google Scholar] [CrossRef]
- Richardson, W.M.; Sodhi, C.P.; Russo, A.; Siggers, R.H.; Afrazi, A.; Gribar, S.C.; Neal, M.D.; Dai, S.; Prindle, T., Jr.; Branca, M. Nucleotide-binding oligomerization domain-2 inhibits toll-like receptor-4 signaling in the intestinal epithelium. Gastroenterology 2010, 139, 904–917.e906. [Google Scholar] [CrossRef] [Green Version]
- Egan, C.E.; Sodhi, C.P.; Good, M.; Lin, J.; Jia, H.; Yamaguchi, Y.; Lu, P.; Ma, C.; Branca, M.F.; Weyandt, S. Toll-like receptor 4–mediated lymphocyte influx induces neonatal necrotizing enterocolitis. J. Clin. Investig. 2016, 126, 495–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, A.G.; Sim, K.; Rose, G.; Wooldridge, D.J.; Li, M.-S.; Misra, R.V.; Gharbia, S.; Kroll, J.S. Premature neonatal gut microbial community patterns supporting an epithelial TLR-mediated pathway for necrotizing enterocolitis. BMC Microbiol. 2021, 21, 225. [Google Scholar] [CrossRef]
- Greenwood, C.; Morrow, A.L.; Lagomarcino, A.J.; Altaye, M.; Taft, D.H.; Yu, Z.; Newburg, D.S.; Ward, D.V.; Schibler, K.R. Early empiric antibiotic use in preterm infants is associated with lower bacterial diversity and higher relative abundance of Enterobacter. J. Pediatr. 2014, 165, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Fleischmann-Struzek, C.; Goldfarb, D.M.; Schlattmann, P.; Schlapbach, L.J.; Reinhart, K.; Kissoon, N. The global burden of paediatric and neonatal sepsis: A systematic review. Lancet Respir. Med. 2018, 6, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.; Buys, N.; Li, C.; Sun, J.; Yin, C. Effects of prebiotics on sepsis, necrotizing enterocolitis, mortality, feeding intolerance, time to full enteral feeding, length of hospital stay, and stool frequency in preterm infants: A meta-analysis. Eur. J. Clin. Nutr. 2019, 73, 657–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castagnola, E.; Dufour, C. Role of G-CSF GM-CSF in the management of infections in preterm newborns: An update. Early Hum. Dev. 2014, 90, S15–S17. [Google Scholar] [CrossRef] [PubMed]
- Shaw, A.G.; Sim, K.; Randell, P.; Cox, M.J.; McClure, Z.E.; Li, M.-S.; Donaldson, H.; Langford, P.R.; Cookson, W.O.; Moffatt, M.F. Late-onset bloodstream infection and perturbed maturation of the gastrointestinal microbiota in premature infants. PloS One 2015, 10, e0132923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mai, V.; Torrazza, R.M.; Ukhanova, M.; Wang, X.; Sun, Y.; Li, N.; Shuster, J.; Sharma, R.; Hudak, M.L.; Neu, J. Distortions in development of intestinal microbiota associated with late onset sepsis in preterm infants. PLoS ONE 2013, 8, e52876. [Google Scholar] [CrossRef]
- Okada, H.; Kuhn, C.; Feillet, H.; Bach, J.-F. The ‘hygiene hypothesis’ for autoimmune and allergic diseases: An update. Clin. Exp. Immunol. 2010, 160, 1–9. [Google Scholar] [CrossRef]
- Wold, A.E. The hygiene hypotheslis revised: Is the rising frequency of allergy due to changes in the intestinal flora? Allergy 1998, 53, 20–25. [Google Scholar] [CrossRef]
- O’Mahony, C.; Scully, P.; O’Mahony, D.; Murphy, S.; O’Brien, F.; Lyons, A.; Sherlock, G.; MacSharry, J.; Kiely, B.; Shanahan, F. Commensal-induced regulatory T cells mediate protection against pathogen-stimulated NF-κB activation. PLoS Pathog. 2008, 4, e1000112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cukrowska, B. Microbial and nutritional programming—The importance of the microbiome and early exposure to potential food allergens in the development of allergies. Nutrients 2018, 10, 1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garland, S.M.; Tobin, J.M.; Pirotta, M.; Tabrizi, S.N.; Opie, G.; Donath, S.; Tang, M.L.; Morley, C.J.; Hickey, L.; Ung, L. The ProPrems trial: Investigating the effects of probiotics on late onset sepsis in very preterm infants. BMC Infect. Dis. 2011, 11, 210. [Google Scholar] [CrossRef] [Green Version]
- Low, J.; Soh, S.-E.; Lee, Y.; Kwek, K.; Holbrook, J.; Van der Beek, E.; Shek, L.; Goh, A.; Teoh, O.; Godfrey, K. Ratio of Klebsiella/Bifidobacterium in early life correlates with later development of paediatric allergy. Benef. Microbes 2017, 8, 681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Jin, S.; Wang, J.; Zhang, L.; Mu, Y.; Huang, K.; Zhao, B.; Zhang, K.; Cui, Y.; Li, S. Variations in early gut microbiome are associated with childhood eczema. FEMS Microbiol. Lett. 2019, 366, fnz020. [Google Scholar] [CrossRef] [PubMed]
- Simonyté Sjödin, K.; Hammarström, M.L.; Rydén, P.; Sjödin, A.; Hernell, O.; Engstrand, L.; West, C.E. Temporal and long-term gut microbiota variation in allergic disease: A prospective study from infancy to school age. Allergy 2019, 74, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Melli, L.C.F.L.; do Carmo-Rodrigues, M.S.; Araújo-Filho, H.B.; Mello, C.S.; Tahan, S.; Pignatari, A.C.C.; Solé, D.; de Morais, M.B. Gut microbiota of children with atopic dermatitis: Controlled study in the metropolitan region of São Paulo, Brazil. Allergol. Et Immunopathol. 2020, 48, 107–115. [Google Scholar] [CrossRef]
- Mah, K.; Björkstén, B.; Lee, B.; Van Bever, H.; Shek, L.; Tan, T.; Lee, Y.; Chua, K. Distinct pattern of commensal gut microbiota in toddlers with eczema. Int. Arch. Allergy Immunol. 2006, 140, 157–163. [Google Scholar] [CrossRef]
- Fujimura, K.E.; Sitarik, A.R.; Havstad, S.; Lin, D.L.; Levan, S.; Fadrosh, D.; Panzer, A.R.; LaMere, B.; Rackaityte, E.; Lukacs, N.W. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 2016, 22, 1187–1191. [Google Scholar] [CrossRef] [Green Version]
- Blümer, N.; Herz, U.; Renz, H. Das pränatale/frühkindliche Immunsystem und Allergie–Ergebnisse humaner und tierexperimenteller Studien/Prenatal and early postnatal immune system and allergy-outcomes of human and animal studies. LaboratoriumsMedizin 2004, 28, 273–278. [Google Scholar] [CrossRef]
- Abrahamsson, T.; Jakobsson, H.; Andersson, A.F.; Björkstén, B.; Engstrand, L.; Jenmalm, M. Low gut microbiota diversity in early infancy precedes asthma at school age. Clin. Exp. Allergy 2014, 44, 842–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kogevinas, M.; Sunyer Deu, J.; Vrijheid, M.; Duijts, L. Preterm birth, infant weight gain, and childhood asthma risk: A meta-analysis of 147,000 European children. J. Allergy Clin. Immunol. 2014, 133, 1317–1329. [Google Scholar]
- Woodward, L.J.; Edgin, J.O.; Thompson, D.; Inder, T.E. Object working memory deficits predicted by early brain injury and development in the preterm infant. Brain 2005, 128, 2578–2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhutta, A.T.; Cleves, M.A.; Casey, P.H.; Cradock, M.M.; Anand, K.J. Cognitive and behavioral outcomes of school-aged children who were born preterm: A meta-analysis. Jama 2002, 288, 728–737. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Ginsberg, Y.; Khatib, N.; Weiner, Z.; Beloosesky, R. Maternal inflammation, fetal brain implications and suggested neuroprotection: A summary of 10 years of research in animal models. Rambam Maimonides Med. J. 2017, 8, e0028. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Yu, Y.; Guo, Y.; Wang, Y.; Chang, E.B.; Claud, E.C. Transcriptional modulation of intestinal innate defense/inflammation genes by preterm infant microbiota in a humanized gnotobiotic mouse model. PloS One 2015, 10, e0124504. [Google Scholar] [CrossRef] [Green Version]
- Aarts, E.; Ederveen, T.H.; Naaijen, J.; Zwiers, M.P.; Boekhorst, J.; Timmerman, H.M.; Smeekens, S.P.; Netea, M.G.; Buitelaar, J.K.; Franke, B. Gut microbiome in ADHD and its relation to neural reward anticipation. PloS One 2017, 12, e0183509. [Google Scholar] [CrossRef] [Green Version]
- Vanes, L.D.; Murray, R.M.; Nosarti, C. Adult outcome of preterm birth: Implications for neurodevelopmental theories of psychosis. Schizophr. Res. 2021, 247, 41–54. [Google Scholar] [CrossRef]
- Nosarti, C.; Reichenberg, A.; Murray, R.M.; Cnattingius, S.; Lambe, M.P.; Yin, L.; MacCabe, J.; Rifkin, L.; Hultman, C.M. Preterm birth and psychiatric disorders in young adult life. Arch. Gen. Psychiatry 2012, 69, 610–617. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, E.; Maukonen, J.; Hyytiäinen, T.; Kieseppä, T.; Orešič, M.; Sabunciyan, S.; Mantere, O.; Saarela, M.; Yolken, R.; Suvisaari, J. Analysis of microbiota in first episode psychosis identifies preliminary associations with symptom severity and treatment response. Schizophr. Res. 2018, 192, 398–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Nallar, E.; Bendall, M.L.; Pérez-Losada, M.; Sabuncyan, S.; Severance, E.G.; Dickerson, F.B.; Schroeder, J.R.; Yolken, R.H.; Crandall, K.A. Composition, taxonomy and functional diversity of the oropharynx microbiome in individuals with schizophrenia and controls. PeerJ 2015, 3, e1140. [Google Scholar] [CrossRef]
- Laverty, C.; Surtees, A.; O’Sullivan, R.; Sutherland, D.; Jones, C.; Richards, C. The prevalence and profile of autism in individuals born preterm: A systematic review and meta-analysis. J. Neurodev. Disord. 2021, 13, 41. [Google Scholar] [CrossRef] [PubMed]
- Yap, I.K.; Angley, M.; Veselkov, K.A.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Urinary metabolic phenotyping differentiates children with autism from their unaffected siblings and age-matched controls. J. Proteome Res. 2010, 9, 2996–3004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milani, C.; Mangifesta, M.; Mancabelli, L.; Lugli, G.A.; James, K.; Duranti, S.; Turroni, F.; Ferrario, C.; Ossiprandi, M.C.; van Sinderen, D. Unveiling bifidobacterial biogeography across the mammalian branch of the tree of life. ISME J. 2017, 11, 2834–2847. [Google Scholar] [CrossRef] [Green Version]
- Tissier, H. Le bacterium coli et la reaction chromophile d’escherich. Crit. Rev. Soc. Biol. 1899, 51, 943–945. [Google Scholar]
- Alessandri, G.; van Sinderen, D.; Ventura, M. The genus Bifidobacterium: From genomics to functionality of an important component of the mammalian gut microbiota. Comput. Struct. Biotechnol. J. 2021, 19, 1472–1487. [Google Scholar] [CrossRef] [PubMed]
- Lugli, G.A.; Milani, C.; Turroni, F.; Duranti, S.; Mancabelli, L.; Mangifesta, M.; Ferrario, C.; Modesto, M.; Mattarelli, P.; van Sinderen, D. Comparative genomic and phylogenomic analyses of the Bifidobacteriaceae family. BMC Genom. 2017, 18, 568. [Google Scholar] [CrossRef] [Green Version]
- Turroni, F.; Peano, C.; Pass, D.A.; Foroni, E.; Severgnini, M.; Claesson, M.J.; Kerr, C.; Hourihane, J.; Murray, D.; Fuligni, F. Diversity of bifidobacteria within the infant gut microbiota. PLoS ONE 2012, 7, e36957. [Google Scholar] [CrossRef] [Green Version]
- Turroni, F.; Foroni, E.; Pizzetti, P.; Giubellini, V.; Ribbera, A.; Merusi, P.; Cagnasso, P.; Bizzarri, B.; de’Angelis, G.L.; Shanahan, F. Exploring the diversity of the bifidobacterial population in the human intestinal tract. Appl. Environ. Microbiol. 2009, 75, 1534–1545. [Google Scholar] [CrossRef] [Green Version]
- Sela, D.A.; Price, N.P.; Mills, D.A. Metabolism of bifidobacteria. Bifidobact. Genom. Mol. Asp. 2010, 45–70. [Google Scholar]
- Stewart, C.J. Breastfeeding promotes bifidobacterial immunomodulatory metabolites. Nat. Microbiol. 2021, 6, 1335–1336. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Ding, Y.; Saedi, N.; Choi, M.; Sridharan, G.V.; Sherr, D.H.; Yarmush, M.L.; Alaniz, R.C.; Jayaraman, A.; Lee, K. Gut microbiota-derived tryptophan metabolites modulate inflammatory response in hepatocytes and macrophages. Cell Rep. 2018, 23, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Bron, P.A.; Van Baarlen, P.; Kleerebezem, M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat. Rev. Microbiol. 2012, 10, 66–78. [Google Scholar] [CrossRef]
- Roberts, D.J. Molecular mechanisms of development of the gastrointestinal tract. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2000, 219, 109–120. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Levy, M.; Itav, S.; Elinav, E. Integration of innate immune signaling. Trends Immunol. 2016, 37, 84–101. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Slack, E.; Geuking, M.B.; McCoy, K.D. The mucosal firewalls against commensal intestinal microbes. Semin. Immunopathol. 2009, 31, 145–149. [Google Scholar] [CrossRef]
- Owaga, E.; Hsieh, R.-H.; Mugendi, B.; Masuku, S.; Shih, C.-K.; Chang, J.-S. Th17 cells as potential probiotic therapeutic targets in inflammatory bowel diseases. Int. J. Mol. Sci. 2015, 16, 20841–20858. [Google Scholar] [CrossRef] [Green Version]
- Inturri, R.; Stivala, A.; Furneri, P.; Blandino, G. Growth and adhesion to HT-29 cells inhibition of Gram-negatives by Bifidobacterium longum BB536 e Lactobacillus rhamnosus HN001 alone and in combination. Eur. Rev. Med. Pharmacol. Sci 2016, 20, 4943–4949. [Google Scholar]
- Abdelhamid, A.G.; Esaam, A.; Hazaa, M.M. Cell free preparations of probiotics exerted antibacterial and antibiofilm activities against multidrug resistant E. coli. Saudi Pharm. J. 2018, 26, 603–607. [Google Scholar] [CrossRef]
- Yun, J.-H.; Kim, Y.-A.; Song, M.-S.; Kang, B.-Y.; Ha, N.-J. Lactic acid bacteria isolated from healthy Korean having antimicrobial activity against VISA and VRE. Yakhak Hoeji 2006, 50, 78–83. [Google Scholar]
- Lkhagvadorj, E.; Nagata, S.; Wada, M.; Bian, L.; Wang, C.; Chiba, Y.; Yamashiro, Y.; Shimizu, T.; Asahara, T.; Nomoto, K. Anti-infectious activity of synbiotics in a novel mouse model of methicillin-resistant Staphylococcus aureus infection. Microbiol. Immunol. 2010, 54, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, M.; Vimont, A.; Darveau, A.; Fliss, I.; Jean, J. Study of the ability of bifidobacteria of human origin to prevent and treat rotavirus infection using colonic cell and mouse models. PLoS ONE 2016, 11, e0164512. [Google Scholar] [CrossRef] [Green Version]
- Ishizuka, T.; Kanmani, P.; Kobayashi, H.; Miyazaki, A.; Soma, J.; Suda, Y.; Aso, H.; Nochi, T.; Iwabuchi, N.; Xiao, J.-z. Immunobiotic bifidobacteria strains modulate rotavirus immune response in porcine intestinal epitheliocytes via pattern recognition receptor signaling. PLoS ONE 2016, 11, e0152416. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Chattha, K.S.; Kandasamy, S.; Liu, Z.; Esseili, M.; Shao, L.; Rajashekara, G.; Saif, L.J. Lactobacilli and bifidobacteria promote immune homeostasis by modulating innate immune responses to human rotavirus in neonatal gnotobiotic pigs. PLoS ONE 2013, 8, e76962. [Google Scholar] [CrossRef] [PubMed]
- Moreno Muñoz, J.A.; Chenoll, E.; Casinos, B.; Bataller, E.; Ramón, D.; Genovés, S.; Montava, R.; Ribes, J.M.; Buesa, J.; Fàbrega, J. Novel probiotic Bifidobacterium longum subsp. infantis CECT 7210 strain active against rotavirus infections. Appl. Environ. Microbiol. 2011, 77, 8775–8783. [Google Scholar] [CrossRef] [Green Version]
- Holscher, H.D.; Czerkies, L.A.; Cekola, P.; Litov, R.; Benbow, M.; Santema, S.; Alexander, D.D.; Perez, V.; Sun, S.; Saavedra, J.M. Bifidobacterium lactis Bb12 enhances intestinal antibody response in formula-fed infants: A randomized, double-blind, controlled trial. J. Parenter. Enter. Nutr. 2012, 36, 106S–117S. [Google Scholar] [CrossRef]
- Lee, A.; Lee, Y.J.; Yoo, H.J.; Kim, M.; Chang, Y.; Lee, D.S.; Lee, J.H. Consumption of dairy yogurt containing Lactobacillus paracasei ssp. paracasei, Bifidobacterium animalis ssp. lactis and heat-treated Lactobacillus plantarum improves immune function including natural killer cell activity. Nutrients 2017, 9, 558. [Google Scholar] [CrossRef] [Green Version]
- HAN, S.; CHO, K.; LEE, C.-K.; SONG, Y.; Park, S.H.; HA, N.-J.; KIM, K. Enhancement of antigen presentation capability of dendritic cells and activation of macrophages by the components of Bifidobacterium pseudocatenulatum SPM 1204. Biomol. Ther. 2005, 13, 174–180. [Google Scholar]
- Kitajima, H.; Sumida, Y.; Tanaka, R.; Yuki, N.; Takayama, H.; Fujimura, M. Early administration of Bifidobacterium breve to preterm infants: Randomised controlled trial. Arch. Dis. Child. Fetal Neonatal Ed. 1997, 76, F101–F107. [Google Scholar] [CrossRef]
- Li, Y.; Shimizu, T.; Hosaka, A.; Kaneko, N.; Ohtsuka, Y.; Yamashiro, Y. Effects of Bifidobacterium breve supplementation on intestinal flora of low birth weight infants. Pediatr. Int. 2004, 46, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Shoji, H.; Sato, H.; Nagata, S.; Ohtsuka, Y.; Shimizu, T.; Yamashiro, Y. Effects of oral administration of Bifidobacterium breve on fecal lactic acid and short-chain fatty acids in low birth weight infants. J. Pediatr. Gastroenterol. Nutr. 2007, 44, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Mohan, R.; Koebnick, C.; Schildt, J.; Schmidt, S.; Mueller, M.; Possner, M.; Radke, M.; Blaut, M. Effects of Bifidobacterium lactis Bb12 supplementation on intestinal microbiota of preterm infants: A double-blind, placebo-controlled, randomized study. J. Clin. Microbiol. 2006, 44, 4025–4031. [Google Scholar] [CrossRef] [Green Version]
- Stratiki, Z.; Costalos, C.; Sevastiadou, S.; Kastanidou, O.; Skouroliakou, M.; Giakoumatou, A.; Petrohilou, V. The effect of a bifidobacter supplemented bovine milk on intestinal permeability of preterm infants. Early Hum. Dev. 2007, 83, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Ohtsuka, Y.; Lee, T.; Kudo, T.; Shoji, H.; Sato, H.; Nagata, S.; Shimizu, T.; Yamashiro, Y. Bifidobacterium breve enhances transforming growth factor β1 signaling by regulating smad7 expression in preterm infants. J. Pediatr. Gastroenterol. Nutr. 2006, 43, 83–88. [Google Scholar] [CrossRef]
- Butel, M.-J.; Suau, A.; Campeotto, F.; Magne, F.; Aires, J.; Ferraris, L.; Kalach, N.; Leroux, B.; Dupont, C. Conditions of bifidobacterial colonization in preterm infants: A prospective analysis. J. Pediatr. Gastroenterol. Nutr. 2007, 44, 577–582. [Google Scholar] [CrossRef]
- Mohan, R.; Koebnick, C.; Schildt, J.; Mueller, M.; Radke, M.; Blaut, M. Effects of Bifidobacterium lactis Bb12 supplementation on body weight, fecal pH, acetate, lactate, calprotectin, and IgA in preterm infants. Pediatr. Res. 2008, 64, 418–422. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.-C.; Hsu, C.-H.; Chen, H.-L.; Chung, M.-Y.; Hsu, J.-F.; Lien, R.-i.; Tsao, L.-Y.; Chen, C.-H.; Su, B.-H. Oral probiotics prevent necrotizing enterocolitis in very low birth weight preterm infants: A multicenter, randomized, controlled trial. Pediatrics 2008, 122, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Mihatsch, W.A.; Vossbeck, S.; Eikmanns, B.; Högel, J.; Pohlandt, F. Effect of Bifidobacterium lactis on the incidence of nosocomial infections in very-low-birth-weight infants: A randomized controlled trial. Neonatology 2010, 98, 156–163. [Google Scholar] [CrossRef]
- Braga, T.D.; da Silva, G.A.P.; de Lira, P.I.C.; de Carvalho Lima, M. Efficacy of Bifidobacterium breve and Lactobacillus casei oral supplementation on necrotizing enterocolitis in very-low-birth-weight preterm infants: A double-blind, randomized, controlled trial. Am. J. Clin. Nutr. 2011, 93, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, C.; Totsu, S.; Uchiyama, A.; Nakanishi, H.; Masumoto, K.; Washio, Y.; Shuri, K.; Ishida, S.; Imai, K.; Kusuda, S. Effect of Bifidobacterium administration on very-low-birthweight infants. Pediatr. Int. 2012, 54, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Ishizeki, S.; Sugita, M.; Takata, M.; Yaeshima, T. Effect of administration of bifidobacteria on intestinal microbiota in low-birth-weight infants and transition of administered bifidobacteria: A comparison between one-species and three-species administration. Anaerobe 2013, 23, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, S.E.; Tobin, J.M.; Opie, G.F.; Donath, S.; Tabrizi, S.N.; Pirotta, M.; Morley, C.J.; Garland, S.M. Probiotic effects on late-onset sepsis in very preterm infants: A randomized controlled trial. Pediatrics 2013, 132, 1055–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patole, S.; Keil, A.D.; Chang, A.; Nathan, E.; Doherty, D.; Simmer, K.; Esvaran, M.; Conway, P. Effect of Bifidobacterium breve M-16V supplementation on fecal bifidobacteria in preterm neonates-a randomised double blind placebo controlled trial. PLoS ONE 2014, 9, e89511. [Google Scholar] [CrossRef]
- Andriantsoanirina, V.; Teolis, A.-C.; Xin, L.X.; Butel, M.J.; Aires, J. Bifidobacterium longum and Bifidobacterium breve isolates from preterm and full term neonates: Comparison of cell surface properties. Anaerobe 2014, 28, 212–215. [Google Scholar] [CrossRef]
- Costeloe, K.; Hardy, P.; Juszczak, E.; Wilks, M.; Millar, M.R. Bifidobacterium breve BBG-001 in very preterm infants: A randomised controlled phase 3 trial. Lancet 2016, 387, 649–660. [Google Scholar] [CrossRef]
- Grześkowiak, Ł.; Teixeira, T.F.S.; Bigonha, S.M.; Lobo, G.; Salminen, S.; Ferreira, C.L.d.L.F. Gut Bifidobacterium microbiota in one-month-old Brazilian newborns. Anaerobe 2015, 35, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Dilli, D.; Aydin, B.; Fettah, N.D.; Özyazıcı, E.; Beken, S.; Zenciroğlu, A.; Okumuş, N.; Özyurt, B.M.; İpek, M.Ş.; Akdağ, A. The propre-save study: Effects of probiotics and prebiotics alone or combined on necrotizing enterocolitis in very low birth weight infants. J. Pediatr. 2015, 166, 545–551.e541. [Google Scholar] [CrossRef]
- Patole, S.K.; Rao, S.C.; Keil, A.D.; Nathan, E.A.; Doherty, D.A.; Simmer, K.N. Benefits of Bifidobacterium breve M-16V supplementation in preterm neonates-a retrospective cohort study. PloS One 2016, 11, e0150775. [Google Scholar] [CrossRef] [Green Version]
- Patole, S.K.; Keil, A.D.; Nathan, E.; Doherty, D.; Esvaran, M.; Simmer, K.N.; Conway, P. Effect of Bifidobacterium breve M-16V supplementation on faecal bifidobacteria in growth restricted very preterm infants–analysis from a randomised trial. J. Matern. Fetal Neonatal Med. 2016, 29, 3751–3755. [Google Scholar] [CrossRef]
- Zhang, S.-F.; Tang, Z.-S.; Tong, L.; Tao, X.-X.; Suo, Q.-F.; Xu, X.-M. Effects of clostridium butyricum and bifidobacterium on BTLA expression on CD4+ T cells and lymphocyte differentiation in late preterm infants. Microb. Pathog. 2016, 100, 112–118. [Google Scholar] [CrossRef]
- Härtel, C.; Pagel, J.; Spiegler, J.; Buma, J.; Henneke, P.; Zemlin, M.; Viemann, D.; Gille, C.; Gehring, S.; Frommhold, D. Lactobacillus acidophilus/Bifidobacterium infantis probiotics are associated with increased growth of VLBWI among those exposed to antibiotics. Sci. Rep. 2017, 7, 5633. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, S.E.; Hickey, L.; Donath, S.; Opie, G.F.; Anderson, P.J.; Garland, S.M.; Cheong, J.L.; Groups, P. Probiotics, prematurity and neurodevelopment: Follow-up of a randomised trial. BMJ Paediatr. Open 2017, 1, e000176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plummer, E.L.; Bulach, D.M.; Murray, G.L.; Jacobs, S.E.; Tabrizi, S.N.; Garland, S.M. Gut microbiota of preterm infants supplemented with probiotics: Sub-study of the ProPrems trial. BMC Microbiol. 2018, 18, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, C.; Savva, G.M.; Clapuci, R.; Jones, J.; Maimouni, H.; Brown, E.; Minocha, A.; Hall, L.J.; Clarke, P. Incidence of necrotising enterocolitis before and after introducing routine prophylactic Lactobacillus and Bifidobacterium probiotics. Arch. Dis. Child. Fetal Neonatal Ed. 2020, 105, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Nakamura, Y.; Terahara, M.; Yanagi, T.; Nakahara, S.; Furukawa, O.; Tsutsui, H.; Inoue, R.; Tsukahara, T.; Koshida, S. Poor Bifidobacterial colonization is associated with late provision of colostrum and improved with probiotic supplementation in low birth weight infants. Nutrients 2019, 11, 839. [Google Scholar] [CrossRef] [Green Version]
- Alcon-Giner, C.; Dalby, M.J.; Caim, S.; Ketskemety, J.; Shaw, A.; Sim, K.; Lawson, M.A.; Kiu, R.; Leclaire, C.; Chalklen, L. Microbiota supplementation with Bifidobacterium and Lactobacillus modifies the preterm infant gut microbiota and metabolome: An observational study. Cell Rep. Med. 2020, 1, 100077. [Google Scholar] [CrossRef]
- Agrawal, S.; Pestell, C.; Granich, J.; Rao, S.; Nathan, E.; Wray, J.; Whitehouse, A.; Patole, S. Difficulties in developmental follow-up of preterm neonates in a randomised-controlled trial of Bifidobacterium breve M16-V—Experience from Western Australia. Early Hum. Dev. 2020, 151, 105165. [Google Scholar] [CrossRef]
- Arboleya, S.; Saturio, S.; Suárez, M.; Fernández, N.; Mancabelli, L.; de Los Reyes-Gavilán, C.G.; Ventura, M.; Solís, G.; Gueimonde, M. Donated human milk as a determinant factor for the gut bifidobacterial ecology in premature babies. Microorganisms 2020, 8, 760. [Google Scholar] [CrossRef] [PubMed]
- Fortmann, I.; Marißen, J.; Siller, B.; Spiegler, J.; Humberg, A.; Hanke, K.; Faust, K.; Pagel, J.; Eyvazzadeh, L.; Brenner, K. Lactobacillus acidophilus/bifidobacterium infantis probiotics are beneficial to extremely low gestational age infants fed human milk. Nutrients 2020, 12, 850. [Google Scholar] [CrossRef] [Green Version]
- Athalye-Jape, G.; Minaee, N.; Nathan, E.; Simmer, K.; Patole, S. Outcomes in preterm small versus appropriate for gestation infants after Bifidobacterium breve M-16 V supplementation. J. Matern. Fetal Neonatal Med. 2020, 33, 2209–2215. [Google Scholar] [CrossRef] [PubMed]
- Juber, B.A.; Boly, T.J.; Pitcher, G.J.; McElroy, S.J. Routine Administration of a Multispecies Probiotic Containing Bifidobacterium and Lactobacillus to Very Low Birth Weight Infants Had No Significant Impact on the Incidence of Necrotizing Enterocolitis. Front. Pediatr. 2021, 1237. [Google Scholar] [CrossRef]
- Tobias, J.; Olyaei, A.; Laraway, B.; Jordan, B.K.; Dickinson, S.; Arroyo, L.G.; Fialkowski, E.; Owora, A.; Scottoline, B. Feeding Activated Bifidobacterium infantis EVC001 to Very Low Birth Weight Infants is Associated with Significant Reduction in Rates of Necrotizing Enterocolitis. MedRxiv 2021. [Google Scholar]
- Wydau-Dematteis, S.; Delannoy, J.; Téolis, A.-C.; Giuseppi, A.; Campeotto, F.; Lapillonne, A.; Butel, M.-J.; Aires, J. Isolation and Characterization of Commensal Bifidobacteria Strains in Gut Microbiota of Neonates Born Preterm: A Prospective Longitudinal Study. Microorganisms 2022, 10, 654. [Google Scholar] [CrossRef] [PubMed]
- Hotel, A.C.P.; Cordoba, A. Health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria. Prevention 2001, 5, 1–10. [Google Scholar]
- Barker, D.J. The origins of the developmental origins theory. J. Intern. Med. 2007, 261, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Authority, E.F.S. Opinion of the Scientific Panel on biological hazards (BIOHAZ) on microbiological criteria and targets based on risk analysis. EFSA J. 2007, 5, 462. [Google Scholar]
- Henrick, B.M.; Rodriguez, L.; Lakshmikanth, T.; Pou, C.; Henckel, E.; Arzoomand, A.; Olin, A.; Wang, J.; Mikes, J.; Tan, Z. Bifidobacteria-mediated immune system imprinting early in life. Cell 2021, 184, 3884–3898.e3811. [Google Scholar] [CrossRef]
- Kavanaugh, D.; O’Callaghan, J.; Butto, L.; Slattery, H.; Lane, J.; Clyne, M.; Kane, M.; Joshi, L.; Hickey, R. Exposure of subsp. To milk oligosaccharides increases adhesion to epithelial cells and induces a substantial transcriptional response. PLoS ONE 2013, 8, e67224. [Google Scholar] [CrossRef] [Green Version]
- Chichlowski, M.; De Lartigue, G.; German, J.B.; Raybould, H.E.; Mills, D.A. Bifidobacteria isolated from infants and cultured on human milk oligosaccharides affect intestinal epithelial function. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 321. [Google Scholar] [CrossRef] [Green Version]
- Underwood, M.A.; Gaerlan, S.; De Leoz, M.L.A.; Dimapasoc, L.; Kalanetra, K.M.; Lemay, D.G.; German, J.B.; Mills, D.A.; Lebrilla, C.B. Human milk oligosaccharides in premature infants: Absorption, excretion, and influence on the intestinal microbiota. Pediatr. Res. 2015, 78, 670–677. [Google Scholar] [CrossRef] [Green Version]
- Dermyshi, E.; Wang, Y.; Yan, C.; Hong, W.; Qiu, G.; Gong, X.; Zhang, T. The “golden age” of probiotics: A systematic review and meta-analysis of randomized and observational studies in preterm infants. Neonatology 2017, 112, 9–23. [Google Scholar] [CrossRef] [PubMed]
- AlFaleh, K.; Anabrees, J. Probiotics for prevention of necrotizing enterocolitis in preterm infants. Evid. Based Child Health Cochrane Rev. J. 2014, 9, 584–671. [Google Scholar] [CrossRef] [PubMed]
- Dani, C.; Biadaioli, R.; Bertini, G.; Martelli, E.; Rubaltelli, F.F. Probiotics feeding in prevention of urinary tract infection, bacterial sepsis and necrotizing enterocolitis in preterm infants. Neonatology 2002, 82, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, P.; Mostert, M.; Leonessa, M.; Priolo, C.; Farina, D.; Monetti, C.; Latino, M.; Gomirato, G. Oral supplementation with Lactobacillus casei subspecies rhamnosus prevents enteric colonization by Candida species in preterm neonates: A randomized study. Clin. Infect. Dis. 2006, 42, 1735–1742. [Google Scholar] [CrossRef] [PubMed]
- Oncel, M.Y.; Sari, F.N.; Arayici, S.; Guzoglu, N.; Erdeve, O.; Uras, N.; Oguz, S.S.; Dilmen, U. Lactobacillus reuteri for the prevention of necrotising enterocolitis in very low birthweight infants: A randomised controlled trial. Arch. Dis. Child. Fetal Neonatal Ed. 2014, 99, F110–F115. [Google Scholar] [CrossRef]
- Rojas, M.A.; Lozano, J.M.; Rojas, M.X.; Rodriguez, V.A.; Rondon, M.A.; Bastidas, J.A.; Perez, L.A.; Rojas, C.; Ovalle, O.; Garcia-Harker, J.E. Prophylactic probiotics to prevent death and nosocomial infection in preterm infants. Pediatrics 2012, 130, e1113–e1120. [Google Scholar] [CrossRef] [Green Version]
- Serce, O.; Benzer, D.; Gursoy, T.; Karatekin, G.; Ovali, F. Efficacy of Saccharomyces boulardii on necrotizing enterocolitis or sepsis in very low birth weight infants: A randomised controlled trial. Early Hum. Dev. 2013, 89, 1033–1036. [Google Scholar] [CrossRef]
- Demirel, G.; Erdeve, O.; Celik, I.H.; Dilmen, U. Saccharomyces boulardii for prevention of necrotizing enterocolitis in preterm infants: A randomized, controlled study. Acta Paediatr. 2013, 102, e560–e565. [Google Scholar] [CrossRef]
- Costalos, C.; Skouteri, V.; Gounaris, A.; Sevastiadou, S.; Triandafilidou, A.; Ekonomidou, C.; Kontaxaki, F.; Petrochilou, V. Enteral feeding of premature infants with Saccharomyces boulardii. Early Hum. Dev. 2003, 74, 89–96. [Google Scholar] [CrossRef]
- Sari, F.; Dizdar, E.; Oguz, S.; Erdeve, O.; Uras, N.; Dilmen, U. Oral probiotics: Lactobacillus sporogenes for prevention of necrotizing enterocolitis in very low-birth weight infants: A randomized, controlled trial. Eur. J. Clin. Nutr. 2011, 65, 434–439. [Google Scholar] [CrossRef] [Green Version]
- Totsu, S.; Yamasaki, C.; Terahara, M.; Uchiyama, A.; Kusuda, S.; Japan, P.S.G.i. bifidobacterium and enteral feeding in preterm infants: Cluster-randomized trial. Pediatr. Int. 2014, 56, 714–719. [Google Scholar] [CrossRef] [Green Version]
- Romond, M.-B.; Colavizza, M.; Mullié, C.; Kalach, N.; Kremp, O.; Mielcarek, C.; Izard, D. Does the intestinal bifidobacterial colonisation affect bacterial translocation? Anaerobe 2008, 14, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Underwood, M.A.; Arriola, J.; Gerber, C.W.; Kaveti, A.; Kalanetra, K.M.; Kananurak, A.; Bevins, C.L.; Mills, D.A.; Dvorak, B. Bifidobacterium longum subsp. infantis in experimental necrotizing enterocolitis: Alterations in inflammation, innate immune response, and the microbiota. Pediatr. Res. 2014, 76, 326–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, K.R.; Liu, S.X.; Tian, R.; Kushnir, A.; Turner, J.R.; Li, H.-L.; Chou, P.M.; Weber, C.R.; De Plaen, I.G. Bifidobacteria stabilize claudins at tight junctions and prevent intestinal barrier dysfunction in mouse necrotizing enterocolitis. Am. J. Pathol. 2013, 182, 1595–1606. [Google Scholar] [CrossRef] [Green Version]
- Plummer, E.L.; Danielewski, J.A.; Garland, S.M.; Su, J.; Jacobs, S.E.; Murray, G.L. The effect of probiotic supplementation on the gut microbiota of preterm infants. J. Med. Microbiol. 2021, 70, 001403. [Google Scholar] [CrossRef] [PubMed]
- Frazer, L.C.; Yakah, W.; Martin, C.R. Decreased Acetic Acid in the Stool of Preterm Infants Is Associated with an Increased Risk of Bronchopulmonary Dysplasia. Nutrients 2022, 14, 2412. [Google Scholar] [CrossRef] [PubMed]
- Cormack, B.E.; Harding, J.E.; Miller, S.P.; Bloomfield, F.H. The influence of early nutrition on brain growth and neurodevelopment in extremely preterm babies: A narrative review. Nutrients 2019, 11, 2029. [Google Scholar] [CrossRef] [Green Version]
- Pärtty, A.; Kalliomäki, M.; Wacklin, P.; Salminen, S.; Isolauri, E. A possible link between early probiotic intervention and the risk of neuropsychiatric disorders later in childhood: A randomized trial. Pediatr. Res. 2015, 77, 823–828. [Google Scholar] [CrossRef] [Green Version]
- McComb, S.; Thiriot, A.; Akache, B.; Krishnan, L.; Stark, F. Introduction to the immune system. In Immunoproteomics; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–24. [Google Scholar]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 461. [Google Scholar] [CrossRef] [Green Version]
- Dąbek, J.; Kułach, A.; Gąsior, Z. Nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB): A new potential therapeutic target in atherosclerosis? Pharmacol. Rep. 2010, 62, 778–783. [Google Scholar] [CrossRef]
- Ganguli, K.; Meng, D.; Rautava, S.; Lu, L.; Walker, W.A.; Nanthakumar, N. Probiotics prevent necrotizing enterocolitis by modulating enterocyte genes that regulate innate immune-mediated inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G132–G141. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Guo, Y.; Ergun, A.; Lu, L.; Walker, W.A.; Ganguli, K. Secreted metabolites of Bifidobacterium infantis and Lactobacillus acidophilus protect immature human enterocytes from IL-1β-induced inflammation: A transcription profiling analysis. PLoS ONE 2015, 10, e0124549. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Nascimento, L.; Massari, P.; Wetzler, L.M. The role of TLR2 in infection and immunity. Front. Immunol. 2012, 3, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Gao, X.-C.; Liu, J.; Ren, H.-Y. Effect of EPEC endotoxin and bifidobacteria on intestinal barrier function through modulation of toll-like receptor 2 and toll-like receptor 4 expression in intestinal epithelial cell-18. World J. Gastroenterol. 2017, 23, 4744. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Wang, L.; Gong, Z.; Wang, Y.; Gao, Y.; Cai, W.; Wu, J. Blockage of NLRP3 inflammasome activation ameliorates acute inflammatory injury and long-term cognitive impairment induced by necrotizing enterocolitis in mice. J. Neuroinflammation 2021, 18, 66. [Google Scholar] [CrossRef]
- Jeon, S.G.; Kayama, H.; Ueda, Y.; Takahashi, T.; Asahara, T.; Tsuji, H.; Tsuji, N.M.; Kiyono, H.; Ma, J.S.; Kusu, T. Probiotic Bifidobacterium breve induces IL-10-producing Tr1 cells in the colon. PLoS Pathog. 2012, 8, e1002714. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Wang, Y.; Zou, J.; Long, F.; Yan, H.; Zeng, L.; Chen, Y. Bifidobacterium adolescentis protects against necrotizing enterocolitis and upregulates TOLLIP and SIGIRR in premature neonatal rats. BMC Pediatr. 2017, 17, 1. [Google Scholar] [CrossRef] [Green Version]
- JIANG, H.; YiQing, Y.; JIANG, W.; RongBin, Z. NLRP3 inflammasome: Activation, regulation, and role in diseases. Sci. Sin. Vitae 2017, 47, 125–131. [Google Scholar]
- Gomez-Lopez, N.; Romero, R.; Garcia-Flores, V.; Leng, Y.; Miller, D.; Hassan, S.S.; Hsu, C.-D.; Panaitescu, B. Inhibition of the NLRP3 inflammasome can prevent sterile intra-amniotic inflammation, preterm labor/birth, and adverse neonatal outcomes. Biol. Reprod. 2019, 100, 1306–1318. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, C.Y.; Osaka, T.; Moriyama, E.; Date, Y.; Kikuchi, J.; Tsuneda, S. Strengthening of the intestinal epithelial tight junction by Bifidobacterium bifidum. Physiol. Rep. 2015, 3, e12327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, X.; Linglong, P.; Weixia, D.; Hong, W. Protective effects of bifidobacterium on intestinal barrier function in LPS-induced enterocyte barrier injury of Caco-2 monolayers and in a rat NEC model. PLoS ONE 2016, 11, e0161635. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, R.; Dharmaprakash, V.; Nighot, P.; Guo, S.; Nighot, M.; Do, T.; Ma, T.Y. Bifidobacterium bifidum Enhances the Intestinal Epithelial Tight Junction Barrier and Protects against Intestinal Inflammation by Targeting the Toll-like Receptor-2 Pathway in an NF-κB-Independent Manner. Int. J. Mol. Sci. 2021, 22, 8070. [Google Scholar] [CrossRef] [PubMed]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface components and metabolites of probiotics for regulation of intestinal epithelial barrier. Microb. Cell Factories 2020, 19, 23. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, D.; Emonet, C.; Foata, F.; Affolter, M.; Delley, M.; Fisseha, M.; Blum-Sperisen, S.; Kochhar, S.; Arigoni, F. A serpin from the gut bacterium Bifidobacterium longum inhibits eukaryotic elastase-like serine proteases. J. Biol. Chem. 2006, 281, 17246–17252. [Google Scholar] [CrossRef] [Green Version]
- Mitsuma, T.; Odajima, H.; Momiyama, Z.; Watanabe, K.; Masuguchi, M.; Sekine, T.; Shidara, S.; Hirano, S. Enhancement of gene expression by a peptide p (CHWPR) produced by Bifidobacterium lactis BB-12. Microbiol. Immunol. 2008, 52, 144–155. [Google Scholar] [CrossRef]
- Collado, M.; Meriluoto, J.; Salminen, S. Role of commercial probiotic strains against human pathogen adhesion to intestinal mucus. Lett. Appl. Microbiol. 2007, 45, 454–460. [Google Scholar] [CrossRef]
- O’Connell Motherway, M.; Houston, A.; O’Callaghan, G.; Reunanen, J.; O’Brien, F.; O’Driscoll, T.; Casey, P.G.; de Vos, W.M.; van Sinderen, D.; Shanahan, F. A Bifidobacterial pilus-associated protein promotes colonic epithelial proliferation. Mol. Microbiol. 2019, 111, 287–301. [Google Scholar] [CrossRef] [Green Version]
- Collado, M.C.; Gueimonde, M.; Sanz, Y.; Salminen, S. Adhesion properties and competitive pathogen exclusion ability of bifidobacteria with acquired acid resistance. J. Food Prot. 2006, 69, 1675–1679. [Google Scholar] [CrossRef] [PubMed]
- Candela, M.; Bergmann, S.; Vici, M.; Vitali, B.; Turroni, S.; Eikmanns, B.J.; Hammerschmidt, S.; Brigidi, P. Binding of human plasminogen to Bifidobacterium. J. Bacteriol. 2007, 189, 5929–5936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trindade, L.; Martins, V.; Rodrigues, N.; Souza, E.; Martins, F.; Costa, G.; Almeida-Leite, C.; Faria, A.; Cardoso, V.; Maioli, T. Oral administration of Simbioflora®(synbiotic) attenuates intestinal damage in a mouse model of 5-fluorouracil-induced mucositis. Benef. Microbes 2018, 9, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Liévin, V.; Peiffer, I.; Hudault, S.; Rochat, F.; Brassart, D.; Neeser, J.; Servin, A. Bifidobacterium strains from resident infant human gastrointestinal microflora exert antimicrobial activity. Gut 2000, 47, 646–652. [Google Scholar] [CrossRef] [Green Version]
- Gibson, G.R.; Wang, X. Regulatory effects of bifidobacteria on the growth of other colonic bacteria. J. Appl. Bacteriol. 1994, 77, 412–420. [Google Scholar] [CrossRef]
- Ohishi, A.; Takahashi, S.; Ito, Y.; Ohishi, Y.; Tsukamoto, K.; Nanba, Y.; Ito, N.; Kakiuchi, S.; Saitoh, A.; Morotomi, M. Bifidobacterium septicemia associated with postoperative probiotic therapy in a neonate with omphalocele. J. Pediatr. 2010, 156, 679–681. [Google Scholar] [CrossRef]
- Brecht, M.; Garg, A.; Longstaff, K.; Cooper, C.; Andersen, C. Lactobacillus sepsis following a laparotomy in a preterm infant: A note of caution. Neonatology 2016, 109, 186–189. [Google Scholar] [CrossRef]
- van den Akker, C.H.; Van Goudoever, J.B.; Szajewska, H.; Embleton, N.D.; Hojsak, I.; Reid, D.; Shamir, R. Probiotics for preterm infants: A strain-specific systematic review and network meta-analysis. J. Pediatr. Gastroenterol. Nutr. 2018, 67, 103–122. [Google Scholar] [CrossRef]
- Aceti, A.; Gori, D.; Barone, G.; Callegari, M.L.; Di Mauro, A.; Fantini, M.P.; Indrio, F.; Maggio, L.; Meneghin, F.; Morelli, L. Probiotics for prevention of necrotizing enterocolitis in preterm infants: Systematic review and meta-analysis. Ital. J. Pediatr. 2015, 41, 89. [Google Scholar] [CrossRef] [Green Version]
- De Simone, C. The unregulated probiotic market. Clin. Gastroenterol. Hepatol. 2019, 17, 809–817. [Google Scholar] [CrossRef] [Green Version]
- Lewis, Z.T.; Shani, G.; Masarweh, C.F.; Popovic, M.; Frese, S.A.; Sela, D.A.; Underwood, M.A.; Mills, D.A. Validating bifidobacterial species and subspecies identity in commercial probiotic products. Pediatr. Res. 2016, 79, 445–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, S.A.; Schoeni, J.L.; Vegge, C.; Pane, M.; Stahl, B.; Bradley, M.; Goldman, V.S.; Burguière, P.; Atwater, J.B.; Sanders, M.E. Improving end-user trust in the quality of commercial probiotic products. Front. Microbiol. 2019, 10, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Reference | Year | Location | Study Design | Aim | Sample Size | Recruiting Center | Inclusion Criteria | Exclusion Criteria | Gestational Age | Weight | Probiotic/Case Group | Probiotic Dose | Placebo/Control Group | Grouping Assignment | Treatment Duration | Disease | Probiotic Safety | Limitation (As Described in the Study) | Conclusion |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [83] | 1997 | Osaka_ Japan | randomized controlled trial | Impact of Bifidobacterium breve YIT4010 (BBG) supplementation on fecal counts and possible adverse effects | 116 (66 probiotic, 50 placebo) | 1 neonatal intensive care unit | birthweight of under 1500 g | major anomalies, severe asphyxia, severe intrauterine growth retardation | ≅28.20 | <1000 g | B breve YIT4010 | 0.5 × 109 | distilled water | randomly allocated | Daily single dose from initiation of milk feeds to 28 days | no side effects | Possible cross-contamination of placebo and probiotic groups | Effective colonization of B. breve, Probiotic association with less abnormal abdominal signs and better weight gain | |
| [84] | 2004 | Tokyo_ Japan | controlled trial | Evaluation of Bifidobacterium breve impact on intestinal flora and fecal Bifidobacterium abundance | 30 (20: probiotic (10: received probiotic several hours after birth (group A), 10: received probiotic 24 h after birth (group)), 10: placebo) | 1 neonatal intensive care unit | admitted to the Neonatal Intensive care unit of Juntendo University Hospital between 2000 and August 2002 | deformities, chromosomal abnormalities, or intrauterine intrauterine infection | ≅32.8 weeks | from 780 to 2250 | Bifidobacterium breve | 1.6 × 108 cells | fed normally without supplement | subjects were randomly divided into three groups | twice a day at the time of normal feeding continued until discharge | Respiratory Distress Syndrome, NEC, sepsis | No side effect | ND | Infants with early Bifidobacterium administration had significantly earlier detectable Bifidobacterium count |
| [85] | 2007 | Tokyo_ Japan | ND | Bifidobacterium breve M-16V supplementation impact on fecal lactic acid and short-chain fatty acids (acetate, propionate, and butyrate acids) | 66 | 1 neonatal intensive care unit, 1 hospital | ND | malformations, chromosomal abnormalities, or intrauterine infections | <36 weeks | <2500 | B breve M-16V (Morinaga Milk Industry, Kanagawa, Japan) | 1.6 × 108 cells | no supplement | randomly divided into probiotic an placebo groups | first day of life irrespective of the use of enteral feeding twice daily until discharge | ND | ND | ND | A lower concentration of fecal acetic acid and butyric acid was detected after probiotic administration which may have protective roles against digestive diseases |
| [86] | 2006 | Potsdam_ Germany | double-blind, placebo-controlled, randomized study | Effect of Bifidobacterium lactis Bb12 on gut microbiota | 69 (37 probiotic, 32 placebo) | 1 hospital | ND | chromosomal aberration, human immunodeficiency virus infection in the mother, hydrops fetalis, and inborn malformation of the gastrointestinal tract | From 30 to 35 weeks | 990 to 2750 g | Bifidobacterium lactis Bb12 | 1.6 × 109 cells on day 1 to 3 and 4.8 × 109 cells from day 4 onward | formula-based placebo | Randoma software version 4.3 | first day after birth and continued for 21 days | ND | ND | ND | A higher fecal abundance of Bifidobacterium in the probiotic group, a lower abundance of Enterobacteriaceae and Clostridium spp. in probiotic group |
| [87] | 2007 | Athens_ Greece | prospective randomized case-control | Investigation of the role of probiotic administration on intestinal permeability, growth, sepsis, and NEC rate | 75 (41 probiotic, 34 placebo) | 1 hospital | gestational age between 27 and 37 weeks, stable state, formula-fed | major deformities, such as congenital heart defects or bowel atresia | <36 weeks | <1500 | formula supplemented with Bifidobacterium lactis (Nestlé, Vevey) | 2 × 107 cfu/g of dry milk | same formula without probiotic | randomly assigned (balanced block randomization) | NEC, sepsis | ND | small sample size | Probiotic administration well-tolerated and decreased intestinal permeability and led to increased head growth and Bifidobacterium count | |
| [88] | 2006 | Tokyo_ Japan | randomized controlled study | The role of Bifidobacterium Breve administration on transforming growth factor A1 signaling (TGF-A1) | 19 (11: probiotic, 8: placebo) | 1 neonatal intensive care unit | ND | chromosomal or congenital anomalies or history of intrauterine infection or surgery, Infants who had received or whose mothers had received corticosteroid treatment | <36 weeks | <2500 | B. breve M-16V g: live but not viable bacteria (Morinaga Milk Industry, Kanagawa, Japan) | 1 × 3 × 109 CFU | 5% glucose solution (without any B. breve) | allocated to 1 of 2 groups | starting several hours after birth twice a day | NEC, Respiratory distress syndrome, Infection, Retinopathy of prematurity, Chronic lung disease | No adverse effect | limited analysis of peripheral samples rather than more elements of the mucosal immune system | B. breve administration could up-regulate TGF-A1 signaling which has anti-inflammatory and allergic responses |
| [89] | 2007 | France | prospective study | Colonization of Bifidobacterium in preterm infants | 52 | 2 hospitals | infants with gestational age ranging from 30 to 35 weeks hospitalized in the neonatal intensive care unit | deformities, chromosomal abnormalities, or inappropriate weight for gestational age | From 30 to 35 weeks | 990 to 2750 g | all infants received standard formula (with 2 probiotic strains (ie, B. breve C50 and Streptococcus thermophilus)) with mother milk | ND | all infants received standard formula (with 2 probiotic strains (ie, B. breve C50 and Streptococcus thermophilus)) with mother milk | ND | ND | ND | ND | ND | Gestational age had a significant impact on Bifidobacterium colonization and gut maturation |
| [90] | 2008 | Potsdam_ German | double-blind placebo-controlled randomized prospective clinical trial | Effects of Bifidobacterium lactis Bb12 Supplementation on indicators of health status (fecal pH, acetate, lactate, calprotectin, IgA, and body weight) | 69 (37 Probiotic, 32 Placebo) | 1 hospital | German ethnic background (except one Russian background) | ND | <37 weeks | <1500 | Bifidobacterium lactis | 1.6 × 109 cells per g of powder | human milk fortifier | randomly assigned | daily for 21 days | ND | ND | ND | Early probiotic supplementation resulted in higher body weight, higher concentrations of fecal acetate, lactate, and IgA, and lower fecal pH. Fecal calprotectin was lower in the probiotic group |
| [91] | 2008 | Taipei_ Taiwan | prospective, blinded, randomized, multicenter controlled trial | Investigation of the efficacy of Bifidobacterium bifidum and Lactobacillus acidophilus probiotics in prevention of NEC | 434 (217 probiotic, 217 placebo) | 7 neonatal intensive care units | Very low birth weight infants who survived to start enteral feeding | severe asphyxia (stage III), fetal chromosomal anomalies, cyanotic congenital heart disease, congenital intestinal atresia, gastroschisis, or omphalocele, infants with exclusive formula feeding, and those who were fasted for more than 3 weeks | <34 weeks | <1500 | Bifidobacterium bifidum and Lactobacillus acidophilus (Infloran) | 109 colony-forming units | breast milk or mixed feeding | assigned randomly to 2 groups using sequential numbers generated at the computer | twice daily, with breast milk or mixed feeding (breast milk and formula) for 6 weeks | NEC (stage ≥ 2) | No adverse effect | ND | NEC rate and mortality were lower in probiotic group |
| [92] | 2010 | Ulm_ Germany | randomized Controlled Trial | Investigation of the role of Bifidobacterium lactis in prevention of nosocomial infections | 103 (93 probiotics, 90 placebo) | 1 Children’s Hospital | infants < 30 weeks of gestation admitted to the Division of Neonatology (Children’s Hospital, University of Ulm, Germany) | early death, congenital malformations or missing parental consent | <30 weeks | <1500 | Bifidobacterium lactis | 6 × 2.0 × 109 CFU/kg/day, 12 billion CFU/kg/day | Human milk fortifier powder | randomly assigned (sealed envelopes, computer-generated, blocked randomization lists, block size of four) | ND | NEC, nosocomial infection | No adverse effect (blood culture) | ND | Probiotic administration did not have a significant impact on nosocomial infection prevention and NEC incidence, B. lactis administration had no adverse effect |
| [93] | 2011 | Recife_ Brazil | prospective, double-blind, randomized, controlled trial | Bifidobacterium breve and Lactobacillus casei supplementation impact on NEC | 231 (119 probiotic, 112 placebo) | 1 neonatal intensive care Unit | Infants born locally and admitted to the Neonatal Intensive Care Unit (NICU) with a birth weight from 750 to 1499 g | major congenital malformations, life-threatening chromosomal alterations, and/or congenital infections | ≅29.35 weeks | 750 to 1499 g | L. casei and B. breve | 3.5 × 107 to 3.5 × 109 CFU (Yakult LB, Brazil) | human milk | randomly assigned using a randomization program (Epi-Info 6.04) | first month of life | NEC (stage ≥2) | no side effects | ND | Probioitc reduced NEC, improved intestinal motility |
| [94] | 2012 | Tokyo_ Japan | prospective randomized control trial | Early Bifidobacterium bifidum OLB6378 supplementation impact on growth and morbidity | 36 (early probiotic supplementation (within 48 h of birth), late probiotic supplementation (more than 48 h after birth)) | 1 maternal and perinatal Center | ND | major congenital malformations, systemic infection, and the lack of parental consent, anticipated feeding problem | <30 weeks | <1500 | B. bifidum/500 mg (Meiji, Tokyo, Japan) | 2.5 × 109 viable cells | ND | random-number generation and a 1:1 allocation | Daily until the bodyweight reached 2 kg | ND | reported safe (monitoring for sepsis with positive blood culture, the length of hospital stay, and the level of B.bifidumin in the fecal samples) | small sample size | Early administration of probiotics had a significant impact on growth (daily body weight gain) and mortality, No significant differences were found in the fecal Bifidobacterium level between the groups (However, it was higher when the supplementation started between 24 and 48 h after birth), |
| [95] | 2013 | Tokyo_ Japan | ND | Comparing single and combined probiotic strains on bifidobacterial abundance | 44 (probiotic (15 one species group, 13 three species group), 16 placebo) | 1 neonatal intensive care unit | infectious diseases, infants treated with antibiotics | <34 weeks | <2000 | Bifidobacterium breve M-16V (one-species group), B. breve M-16V, Bifidobacterium longum subsp. infantis M-63 and B. longum subsp. longum BB536 (mixture of three specie) | 5 × 108 (one-species group), 5 × 108 (of each strain; three-species group) | ND | ND | Daily from the beginning of enteral nutrition for 6 weeks | ND | ND | Significant increase of Bifidobacteria count in the probiotic group, Three species probiotics resulted in earlier formation of bifidobacterial colonization, Lower abundance of Clostridium in the probiotic group, Lower Enterobacteriaceae abundance in the three species, the proportion of bifidobacteria in the three-species group was significantly higher than that in the one-species group, B. breve M-16V and Bifidobacterium infantis M-63 were detected in majority of infants | ||
| [96] | 2013 | Melbourne_ Australia | prospective multicenter, double-blinded, placebo-controlled, randomized trial | Evaluation of the impact of Probiotics (B. infantis, S. thermophilus, and B. lactis) on Los | 1099 (548 probiotic, 551 placebo) | 10 perinatal hospitals | infants with <32 weeks gestational age and weighing <1500 g within 72 h of birth | major congenital or chromosomal anomalies, if death was considered likely within 72 h of birth if mothers were taking nondietary probiotic supplements | <32 weeks | <1500 | Bifidobacterium infantis, Streptococcus thermophilus, and Bifidobacterium lactis (ABC Dophilus Probiotic Powder for Infants; Solgar, Leonia, New Jersey) | 300 × 106 (Bifidobacterium longum subsp. Infantis BB–02), 350 × 106 (TH–4, Streptococcus thermophilus), 350 × 106 (Bifidobacterium animalis subsp. Lactis BB-12) | maltodextrin | 1:1 allocation using STATA | Daily until discharge from hospital or term corrected age | NEC, Los | reported safe | ND | A significant decrease in NEC rate (stage 2 or more) (but not sepsis, and all-cause mortality) was observed after probiotics administration, No probiotic adverse effect was reported |
| [97] | 2014 | Perth_ Australia | randomized double-blinded placebo-controlled trial | Role of Bifidobacterium breve (B. breve) M-16V supplementation on fecal counts and possible adverse effects | 159 (79 Probiotic, 80 Placebo) | 1 tertiary neonatal intensive care unit | Infants with <32 weeks and 6 days, under 1500 g, ready to commence or on enteral feeds for <12 h | Major congenital malformation, chromosomal aberration, lack of informed parental consent, enteral feeds for ≥12 h, contraindications for enteral feeds, life-threatening illness | <33 weeks | <1500 g | B. breve M-16V | 3 × 109 cfu/day | dextrin | randomly allocated | Daily single dose until the corrected age of 37 weeks | NEC (≥Stage 2) | no side effects: blood culture for B. breve M-16V, monitoring adverse effects such as abdominal distension, vomiting, and diarrhea | Immediate supplementation of probiotic without considering B. breve counts in meconium | Routine use of B. breve M-16V is safe, No adverse effect was reported, Probiotics significantly increased B. breve fecal count |
| [98] | 2014 | France | ND | Comparison of cell surface properties (autoaggregation, hydrophobicity, and Caco-2 cells adhesion) of B. longum and B.breve isolates in preterm and full-term infants | 47 (20 preterm, 27 full term) | ND | ND | ND | <36 weeks | ND | ND | ND | ND | ND | ND | ND | ND | ND | Cell surface properties were different between Bifidobacterium strains isolated from preterm and full-term infants |
| [99] | 2016 | London_ UK | multicentre, randomized controlled phase 3 study | Evaluation of Bifidobacterium breve BBG-001 supplementation impact on NEC, sepsis | 1310 (650 probiotic, 660 placebo) | 24 hospitals | ND | Infants with a potentially lethal malformation or any malformation of the gastrointestinal tract apparent by 48 h and those with no chance of survival | From 23 and 30 weeks | <1000 g | B breve BBG-001 (Yakult Honsha Co Ltd.) | enterally in a daily dose of 8·2 to 9·2 log10 CFU | corn starch | based on date of birth | Daily single dose until the infant reaches a corrected post-menstrual age | NEC (Bell’s stage 2 or 3) | no short-term safety | Possible cross-contamination of placebo and probiotic groups | Probiotic did not have any significant impact on NEC and sepsis, |
| [100] | 2015 | Viçosa, Brazil | pilot study | Evaluation of Bifidobacterial composition in full-term and preterm infants | 49 (24 full term, 25 preterm) | 1 hospital | availability of fecal samples and signed informed consent from the mother. Being residents of Viçosa and neighboring areas | ND | <39 weeks | <3500 | one-month-old preterm infants | ND | one-month-old full-term infants | ND | ND | ND | ND | ND | Bifidobacterium longum colonized in all full-term and pre-term newborns. Variation in fecal counts of Bifidobacterium genus and Bifidobacterium longum between full-term and pre-term infants. Variation of Bifidobacterium lactis abundance between preterm cesarean and pre-term vaginally born infants. |
| [101] | 2015 | Turkey | multicenter, prospective, randomized, double-blind, randomized controlled trial (RCT) | Investigation of the prevention role of probiotics and prebiotics (alone or combined (synbiotic)) on necrotizing enterocolitis | 400 (100: probiotic, 100: prebiotic, 100: synbiotic, 100: placebo) | 5 neonatal intensive care units | gestational age of <32 weeks and a birth weight of <1500 g, born at or transferred to the NICUwithin the first week of life and fed enterally before inclusion | Infants with any disease other than those linked to prematurity or congenital anomalies of the intestinal tract, not fed enterally or who died before the seventh day after birth, whose mothers had taken nondietary probiotic supplements, and whose parents refused to participate | <32 weeks | <1500 | probiotic (Bifidobacterium lactis), prebiotic (inulin), synbiotic (Bifidobacterium lactis) | probiotic (5 × 109 colony-forming units), prebiotic (900 mg), synbiotic (5 × 109 colony-forming units probiotic, 30 mg plus inulin, 900 mg) | breastmilk or formula without the addition of probiotic or prebiotic and received maltodextrin | randomly assigned (balanced blocks using sealed envelope) | maximum of 8 weeks before discharge or death (variables between groups) | Bell stage II-III, bronchopulmonary dysplasia, intraventricular hemorrhage, cystic periventricular leukomalacia, and retinopathy of prematurity | findings cannot be generalized to all probiotics with different doses, limited inclusion to infants who survived beyond the 7 days of life | Probiotic (Bifidobacterium lactis) and synbiotic (Bifidobacterium lactis plus inulin), but not prebiotic (inulin) alone could decrease the NEC rate in the probiotic group | |
| [102] | 2016 | Perth_ Australia | retrospective cohort study | Bifidobacterium breve M-16V supplementation impact on NEC | 1755 (920 probiotic, 835 placebo) | ND | preterm neonates born <34 weeks | major congenital malformations, chromosomal aberrations, and contraindications for enteral feeding, and those with no informed consent | <34 weeks | ≅1340 | Bifidobacterium breve M16V | 3 × 109 (3 billion) cfu/day | ND | ND | Daily single dose continued until the corrected age of 37 weeks | NEC (stage ≥2) | no adverse effects monitoring for sepsis and abdominal distension, vomiting, and diarrhea | It was a retrospective design, which made it difficult to control all confounders | Bifidobacterium breve M-16V was associated with reduced NEC and mortality |
| [103] | 2016 | Perth_ Australia | analysis of a randomized trial | Impact of Bifidobacterium breve M-16V supplementation on fecal Bifidobacterium | 153 (77 probiotic, 76 placebo) | 1 tertiary neonatal intensive care unit | Preterm infants with small for gestational age due to small for gestational age | chromosomal aberrations, congenital malformation | <33 weeks | <825 | B. breve M-16V | 3 × 109 cfu/day | dextrin | randomly allocated | until the corrected age of 37 weeks | ND | reported safe (by monitoring blood culture positive sepsis by B. breve M-16V and adverse effects such as abdominal distension, vomiting, and diarrhea leading to the cessation of the supplementation) | ND | B. breve M-16V supplementation did not change the detectable B. breve counts between infants with small gestational age (SGA) and non-SGA |
| [104] | 2016 | Anhui_ China | ND | Impact of clostridium butyricum and bifidobacterium (LCBBCP) on the expression of B and T lymphocyte attenuator (BTLA) on CD4 cells | 80 | 1 neonatal intensive care unit | ND | neonatal comorbidities (including asphyxia, infection, congenital malformation, respiratory distress syndrome, pneumorrhagia, congenital immunodeficiency, and other related conditions), maternal infectious diseases during pregnancy and autoimmune disorders | <37 weeks | ND | clostridium butyricum and bifidobacterium (Changlekang, China) | ND | simple formula milk | equally randomized (random digit table) | twice a day for 7 days | ND | ND | ND | LCBBCP had inhibitory impact on excessive activation of T lymphocytes |
| [105] | 2017 | Germany | observational study | Impact of Bifidobacterium infantis and Lactobacillus acidophilus supplementation on preterm infant growth under antibiotic exposure | 8534 (6229 probiotic, 2305 placebo) | 54 neonatal intensive care units | birth weight <1500 g, gestational age >22 0/7 and ≤32 6/7 weeks, written informed consent of parents or legal representatives, and discharge to the home environment | lethal malformations, e.g., trisomy 13 and trisomy 18 | ≤33 weeks | <1500 | (Infloran) | 109 | ND | ND | Daily for 28 days | ND | ND | It was an observational study, not a double-blinded, randomized controlled study, which made it difficult to control confounders and interpret findings properly. For a follow-up cohort, the sample size is not sufficient. Variable duration of hospitalization observed in participants. Bias in designing the study since probiotics were more often given to infants <28 weeks of age | Probiotic supplementation had beneficial impact on weight gain and growth rate in infants under antibiotic exposure |
| [106] | 2017 | Melbourne_ Australia | double-blinded, placebo-controlled, randomized trial | Determination of probiotic combination (B. infantis, S. thermophilus, and B. lactis) on neurodevelopmental outcomes in very preterm infants. Follow up | Follow-up 735 (373 probiotics, 362 placeboes) | 10 perinatal hospitals | participants in the ProPrems trial | Children for whom disability status could not be determined | <32 weeks | <1500 | Bifidobacterium infantis, Streptococcus thermophilus, and Bifidobacterium lactis (ABC Dophilus Probiotic Powder for Infants; Solgar, Leonia, New Jersey) | 300 × 106 (Bifidobacterium longum subsp. Infantis BB–02), 350 × 106 (TH–4, Streptococcus thermophilus), 350 × 106 (Bifidobacterium animalis subsp. Lactis BB-12) | maltodextrin | Daily until discharge from hospital or term corrected age | NEC, Los | reported safe | declined and lost follow-up participants since the study was not planned as an outcome of the ProPrems trial, a wide age range among participants, which may impact the power of the study to find differences between two groups | No neurodevelopmental and behavioral adverse effect was detected after combined probiotic administration | |
| [107] | 2018 | Melbourne_ Australia | multi-center, double-blind, placebo-controlled randomized trial | Investigation of the role of Bifidobacterium infantis BB-02, Bifidobacterium lactis BB-12, and Streptococcus thermophilus TH-4 probiotic on gut microbiota composition | 66 (38 probiotic, 28 placebo) | 1 newborn Intensive Care Unit | infants enrolled at The Royal Women’s Hospital, Melbourne, Australia with at least one swab available | ND | <32 weeks | <1500 | Bifidobacterium longum subsp. Infantis, Streptococcus thermophilus, Bifidobacterium animalis subsp. Lactis (ABC Dophilus Probiotic Powder for Infants; Solgar, Leonia, New Jersey) | 300 × 106 (Bifidobacterium longum subsp. Infantis BB–02), 350 × 106 (TH–4, Streptococcus thermophilus), 350 × 106 (Bifidobacterium animalis subsp. Lactis BB-12) | maltodextrin powder | adjusting for age at sampling | once enteral feeds were commenced until discharge or term-corrected age | ND | ND | limited taxonomy classification to the genus level, cross-colonization in the control group, and a small number of ProPrems participants, due to the variable number of samples per infant colonization patterns could not be established for all infants, only 11 specimens collected before supplementation commenced were available (not clear if there was a gut microbial difference between the two allocation groups before supplementation), due to the lower NEC incidence in the selected participants for this study comparison of gut microbiota in NEC and NonNEC infants were not possible | A higher abundance of Bifidobacterium in the probiotic group, lower Enterococcus abundance in the probiotic group, early BB-02, TH-4, and BB-12 supplementation increased the Bifidobacterium abundnace |
| [108] | 2019 | Norwich_ UK | single- center retrospective observational study | Lactobacillus and Bifidobacterium supplementation impact on NEC, sepsis, and mortality | 982 (pre-probiotic epoch = 469, routine probiotics = 513) | 1 tertiary- level neonatal intensive care unit | <32 weeks’ gestation, plus 32–36 weeks’ gestation VLBW infants. Outborn babies were included if transferred within 72 h of birth | abdominal concerns at referral | <36 weeks | <1500 | Initially Bifidobacterium and Lactobacillus (Infloran capsules), then triple-species Labinic Drops: four drops once daily | Initially 109 colony-forming units (CFU) (Bifidobacterium and Lactobacillus) then ~0.5 × 109 CFU dosage each of L. acidophilus, B. bifidum, and B. longum subspecies infantis daily | donor breast milk (DBM) was available to supplement shortfalls in mother’s own breast milk supply before full feeds. Cow’s milk-based fortifier was added to breast milk between full enteral feeds (≥150 mL/kg/day) and discharge | allocated by date of birth | twice daily on postnatal day 1 until ~34 weeks postmenstrual age | NEC, sepsis | no safety issues | It was a retrospective observational study which made it difficult to control confounders and interpret findings properly | A significant decrease in NEC incidence and sepsis after multispecies probiotic supplementation was observed |
| [109] | 2019 | Japan | ND | Impact of probiotic supplementation and timing of initial colostrum on Bifidobacterium colonization | 98 (group H:37, group L = 30. group N = 31) | 1 neonatal intensive care unit | ND | preterm infants without informed consent, congenital malformations | <36 weeks | <2500 | group H (received non-live bifidobacteria), and group L (received live bifidobacteria). | Group L: a mixture of 20 mg of live OLB6378 powder (containing 10 mg of lyophilized live OLB6378 concentrate with >2.5 × 109 live cells) and 480 mg of dextrin, Group H: a mixture of 20 mg of lyophilized non-live OLB6378 powder (containing 10 mg of lyophilized non-live OLB6378 concentrate with >2.5 × 109 non-live cells) and 480 mg of dextrin | Group N (no intervention) | ND | within 48 h after birth and continued for at least 1 month after birth | ND | ND | different measurement of bifidobacterial colonization in preterm and term infants, not performing multiple regression analysis | Bifidobacterial colonization in preterm infants at 1 month was low compared to term infants, Higher Bifidobacterium colonization was detected after probiotic administration in groups H, and L, Earlier consumption of colostrum had a significant impact on the fecal Bifidobacterium count/abundance |
| [110] | 2020 | Norwich_ UK | observational longitudinal study | Impact of Bifidobacterium and Lactobacillus probiotic on fecal metabolites and gut microbiota | 234 (101 probiotic, 133 placebo) | 4 tertiary-level NICUs | premature infants born at gestational age <34 weeks, and resident in the same NICU for the study duration | necrotizing enterocolitis or severe congenital abnormalities | <28 weeks | <1500 g | Bifidobacterium bifidum, Lactobacillus acidophilus (Infloran, Desma Healthcare, Chiasso, Switzerland): | 109 colony-forming units (CFU) of Bifidobacterium bifidum and 109 CFU of Lactobacillus acidophilus | ND | matched by age, sex, and delivery method | Twice daily from the first enteral colostrum/milk feed until 34 weeks post-conceptual age | ND | It was an observational study, not a double-blinded, randomized controlled study, which made it difficult to control confounders and interpret findings properly, not monitor the impact of feeding diet on microbiota in all infants, not measure absolute abundance of bacterial taxa | Association of probiotics with higher abundance of Bifidobacterium and higher fecal acetate/lactate concentration, and lower fecal pH | |
| [111] | 2020 | Perth_ Australia | follow up of a randomized controlled trial | Evaluation of long-term neuropsychological effects of early probiotic supplementation in preterm infants | 67 (36 probiotics, 31 placebo) | 1 tertiary neonatal intensive care unit | preterm neonates (born <33 weeks) in the original RCT of probiotic for preventing NEC | Major congenital malformation, chromosomal aberration, lack of informed parental consent, enteral feeds for ≥12 h, contraindications for enteral feeds, life-threatening illness | <33 week | <1105 | B. breve M-16V | 3 × 109 cfu/day | dextrin | randomly allocated | Daily single dose until the corrected age of 37 weeks | no side effects: blood culture for B. breve M-16V, monitoring adverse effects such as abdominal distension, vomiting, and diarrhea | low follow-up rate | Probiotics did not have any significant effect on neurodevelopment at the age of 3 to 5 years | |
| [112] | 2020 | Spain | a prospective and observational study | Impact of donated human milk on gut Bifidobacterial profile and metabolism | 42 (28 own mother milk, 13 donated milk) | 1 hospital | ND | Mixed feeding and use of probiotics, prebiotics, or other treatments. NEC, culture-positive early-onset infection, major malformations, or surgery of the intestinal tract | between 24 and 34 weeks | 1334.88 ± 338.64 g (mean ± SD) | human donor milk-fed preterm infants | ND | breastfed preterm infants | ND | At least for the first ten days of life: preterm infants received their own mother’s milk or donated milk. At 30 days of life, half of the babies received formula, with only three babies with their own mother’s milk. the OMM regimen. | ND | ND | mall number of infants and the large interindividual variability, confounding factors, such as antibiotics could influence the findings | A specific bifidobacterial profile was detected based on feeding type. Higher bifidobacterial diversity in the human donor milk group |
| [113] | 2020 | Germany | observational study | Impact of Lactobacillus Acidophilus, Bifidobacterium Infantis probiotic on growth and sepsis | 7516 | 64 neonatal intensive care units | infants with complete documentation for feeding type | lethal malformations or infants treated with comfort (palliative) care | <29 weeks | <1500 | Lactobacillus Acidophilus, Bifidobacterium Infantis | 1 × 3 × 109 CFU (Colony forming units) L. acidophilus and 1 × 1.5 × 109 B. infantis | exclusively fed with own mother‘s and/or donor‘s milk, fed with HM and formula at any time during the primary stay in hospital, Infants who were exclusively fed with formula | ND | once or twice daily in capsules beginning from day 1 to 3 of life until day 28 of life | sepsis, Bronchopulmonary dysplasia, NEC, focal intestinal perforation, Retinopathy of prematurity | ND | observational design, lack of information on the daily type of feeding in the Mix group, indication for supplementation, and timing with the bovine and individual fortification of human milk or formula | Probiotic supplementation had protective and promotive roles on sepsis and growth, respectively |
| [114] | 2020 | Perth_ Australia | Retrospective cohort | Comparing clinical outcomes of Bifidobacterium breve M-16V supplementation in full-term and preterm infants | 1380 (162 preterm, 1218 full term) | 1 neonatal intensive care unit | preterm neonates (gestation <34 weeks) admitted between June 2012 and August 2015 | ND | <34 weeks, subgroup: <29 weeks | <1500 | B. breve M-16 V (Morinaga Milk Industries, Tokyo, Japan) | 3 × 109 cfu/day | ND | gestational age, gender, duration of respiratory support, and patent ductus arteriosus were controlled as confounding factors | twice daily continued till the 37 weeks gestational age or discharge (Supplementation was stopped after suspected NEC or sepsis diagnosis) | NEC (stage ≥ 2), Los | No adverse effect by monitoring blood culture positive sepsis by B. breve M-16V, abdominal distension, vomiting, and diarrhea leading to the cessation of the supplementation | retrospective design that makes it difficult to determine the effects of known and unknown confounders, relatively small sample size | No significant difference was observed between the two groups regarding NEC rate, LOS, and mortality. postnatal age at full feeds was higher in preterm infants |
| [115] | 2021 | Iowa_ USA | single-center retrospective chart review | Evaluation of Bifidobacterium and Lactobacillus supplementation impact on NEC | 37 (14 Pre-probiotic, 23 Post-probiotic) | 1 Children’s Hospital | Probiotic group: Infants with at least 3 days old, born at <33 weeks gestational age, with a corrected post-menstrual age of at least 24 0/7 weeks who received intakeof at least 6mL of enteral feedings per day | Infants with major congenital abnormalities, anatomic obstruction of the gastrointestinal tract, inguinal hernia repair, G-tube placement, or peritoneal dialysiscatheter placement | <33 weeks | <1500 g | multispecies probiotic (Bifidobacterium breve, bifidum, infantis, and longum) plus Lactobacillus rhamnosus GG (Ultimate Flora Baby Probiotic) | 2 × 109 colony forming units per 0.5 g. | ND | randomly assigned (1:1) using a randomization program | Daily until the infant reaches a corrected post-menstrual age | NEC (Bell’s stage ≥2a) | NA | It was a retrospective study, which made it difficult to control confounders and interpret findings properly, single-center design | Probiotic supplementation did not have significant impact on the NEC rate and mortality |
| [116] | 2021 | USA | non-concurrent, retrospective cohort study | Bifidobacterium infantis EVC001 supplementation impact on NEC rate | 483 (182 probiotics, 301 placeboes) | 1 hospital | weight <1500g, received full resuscitation, and survived until day-of-life 3, (the earliest time at which very low birth weight infants received at least one feed of EVC001, fed human milk-based diets consisting of either mother’s milk, donor milk, or a combination, fed according to institutional guidelines incorporating best practices for NEC prevention, including a human milk-based diet, an initial period of trophic feeding and gradual feeding advancements, did not have hemodynamically significant congenital heart disease | underwent palliative delivery or unsuccessful resuscitation, died prior to day-of-life 4, were fed a non-human milk-based diet prior to 34 weeks PMA, immunodeficiency, received less than two feeds of EVC001 in EVC001 group | ≅28 weeks | <1500 | active B. infantis EVC001 | 8 billion colony forming units (CFU) in 0.5 mL of medium chain triglyceride oil | human milk-based diet of mother’s milk, donor milk, or both | adjusted for sex, birth weight, gestational age at birth, and mode of delivery | Daily prior to morning feed until 34 weeks post-menstrual age or for a minimum of two weeks, whichever duration was longer | NEC (≥Stage 2) | reported safe | observational design, absence of fecal sampling to confirm that B. infantis EVC001 supplementation led to successful modulation of the preterm intestinal microbiota | Probiotic administration led to a significant reduction in the NEC rate and mortality |
| [117] | 2022 | Paris_ France | prospective longitudinal study | Characterization of Bifidobacteria strains isolated from preterm infants | 26 | 1 pediatric hospital | Preterm infants with at least 2 fecal samples with bifidobacterial colonization at different times | Preterm infants with malformations or metabolic diseases | <37 weeks | From 710 to 2610 g | ND | ND | ND | ND | ND | ND | ND | Low genotype resolution | Environmental factors can affect phenotypes in Bifidobacterium strains. Phenotypes and genotypes of Bifidobacteria species were unstable during the first year of life. Twin infants have a more similar microbiota compared to other infants |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadeghpour Heravi, F.; Hu, H. Bifidobacterium: Host–Microbiome Interaction and Mechanism of Action in Preventing Common Gut-Microbiota-Associated Complications in Preterm Infants: A Narrative Review. Nutrients 2023, 15, 709. https://doi.org/10.3390/nu15030709
Sadeghpour Heravi F, Hu H. Bifidobacterium: Host–Microbiome Interaction and Mechanism of Action in Preventing Common Gut-Microbiota-Associated Complications in Preterm Infants: A Narrative Review. Nutrients. 2023; 15(3):709. https://doi.org/10.3390/nu15030709
Chicago/Turabian StyleSadeghpour Heravi, Fatemah, and Honghua Hu. 2023. "Bifidobacterium: Host–Microbiome Interaction and Mechanism of Action in Preventing Common Gut-Microbiota-Associated Complications in Preterm Infants: A Narrative Review" Nutrients 15, no. 3: 709. https://doi.org/10.3390/nu15030709
APA StyleSadeghpour Heravi, F., & Hu, H. (2023). Bifidobacterium: Host–Microbiome Interaction and Mechanism of Action in Preventing Common Gut-Microbiota-Associated Complications in Preterm Infants: A Narrative Review. Nutrients, 15(3), 709. https://doi.org/10.3390/nu15030709