
Effects of 4′-Demethylnobiletin and 4′-Demethyltangeretin on Osteoclast Differentiation In Vitro and in a Mouse Model of Estrogen-Deficient Bone Resorption


 ,
,  ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Reagents
2.2. Osteoclast Differentiation in Cocultures of Mouse Primary Osteoblasts and Mouse Bone Marrow Cells
2.3. Osteoclast Differentiation in Cultures of Raw264.7 Cells
2.4. TRAP Staining
2.5. Analysis of mRNA Expression by Quantitative PCR
2.6. Analysis of Protein Expression by Western Blotting
2.7. In Silico Molecular Docking Simulation
2.8. Intraperitoneal Administration of a Mixture of Demethylated Compounds to OVX Mice
2.9. Statistical Analysis
3. Results
3.1. Comparative Effects of NOB, TAN and Its Demethylated Compounds on Osteoclast Differentiation
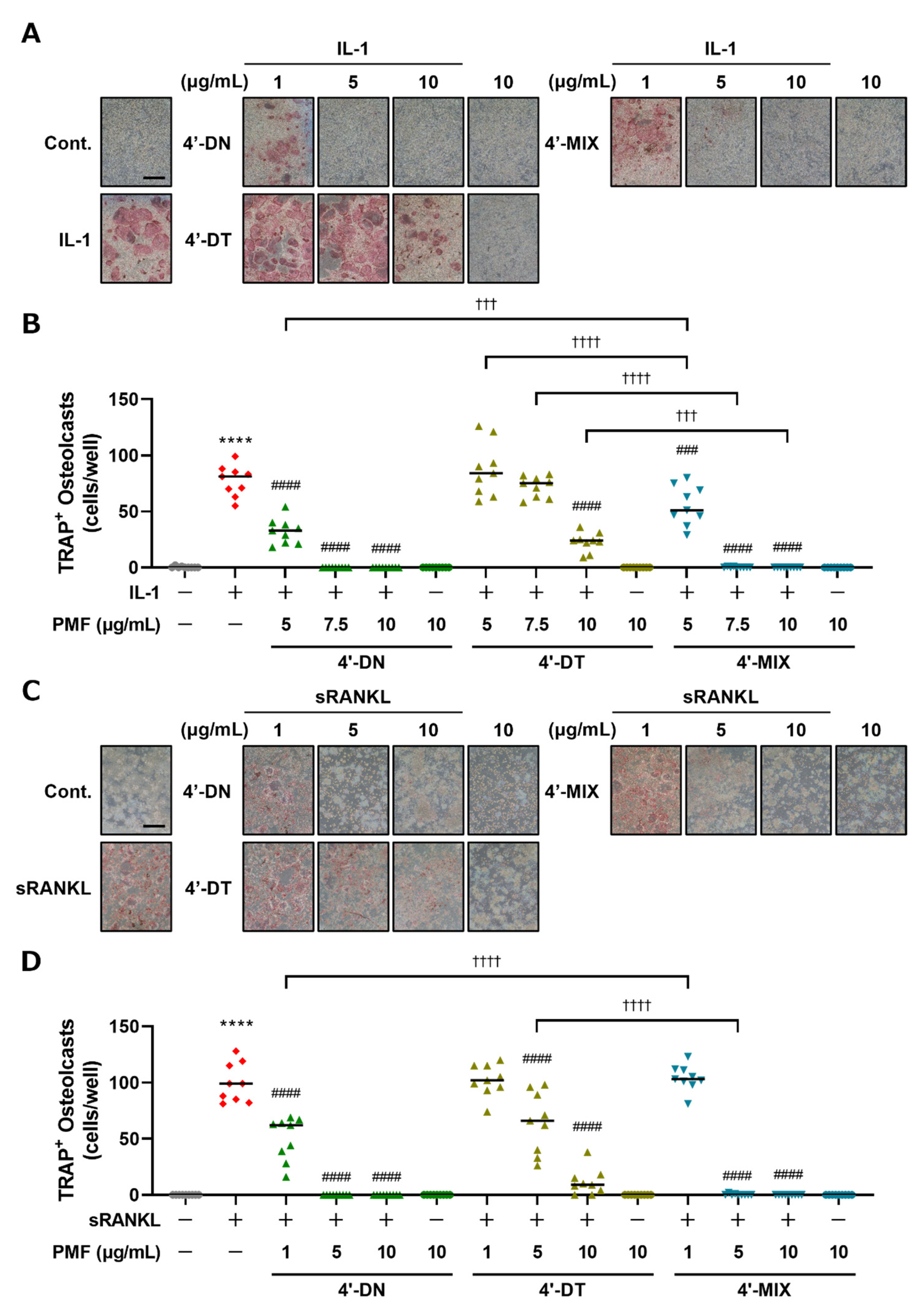
3.2. Mixture of 4′-DN and 4′-DT Suppressed Osteoclast Differentiation in Cocultures of POB and BMC and Raw264.7 Cultures
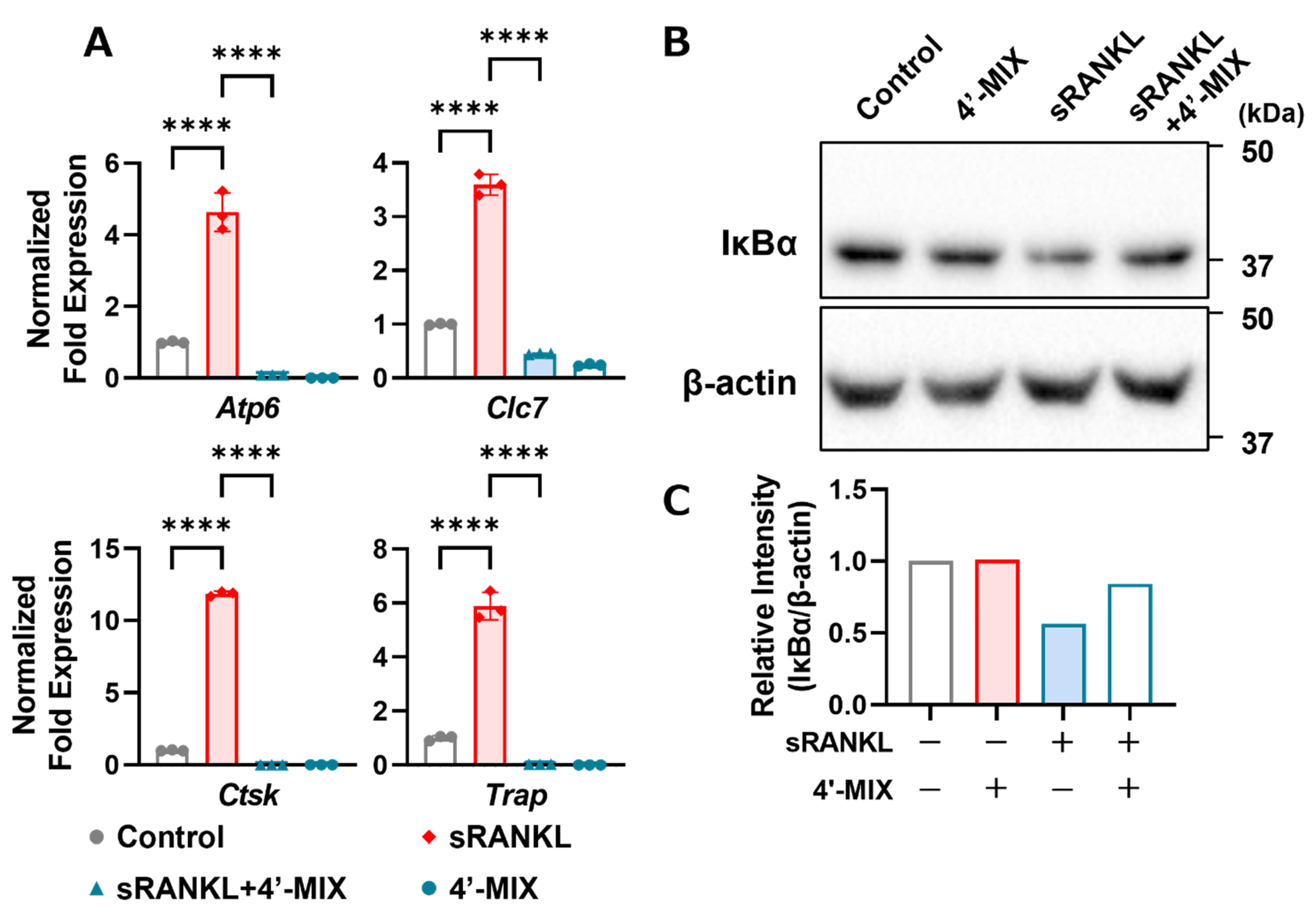
3.3. Mixture of 4′-DN and 4′-DT Inhibits Osteoclast Marker Genes and IκBα Protein Degradation by Adding RANKL
3.4. In Silico Molecular Docking Simulation of 4′-DN and 4′-DT to ATP Pocket of IKKβ Protein
3.5. Intraperitoneal Administration of 4′-MIX Inhibits Estrogen Deficiency-Induced Bone Loss in OVX Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yasuda, H.; Shima, N.; Nakagawa, N.; Yamaguchi, K.; Kinosaki, M.; Mochizuki, S.; Tomoyasu, A.; Yano, K.; Goto, M.; Murakami, A.; et al. Osteoclast Differentiation Factor Is a Ligand for Osteoprotegerin/Osteoclastogenesis-Inhibitory Factor and Is Identical to TRANCE/RANKL. Proc. Natl. Acad. Sci. USA 1998, 95, 3597–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, S.; Tanaka, Y.; Ishiguro, N.; Yamanaka, H.; Takeuchi, T. Rankl: A Therapeutic Target for Bone Destruction in Rheumatoid Arthritis. Mod. Rheumatol. 2017, 28, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Inada, M.; Matsumoto, C.; Uematsu, S.; Akira, S.; Miyaura, C. Membrane-Bound Prostaglandin E Synthase-1-Mediated Prostaglandin E2 Production by Osteoblast Plays a Critical Role in Lipopolysaccharide-Induced Bone Loss Associated with Inflammation. J. Immunol. 2006, 177, 1879–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanabe, N.; Maeno, M.; Suzuki, N.; Fujisaki, K.; Tanaka, H.; Ogiso, B.; Ito, K. IL-1α Stimulates the Formation of Osteoclast-like Cells by Increasing M-CSF and PGE2 Production and Decreasing OPG Production by Osteoblasts. Life Sci. 2005, 77, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Modder, U.; Riggs, B.; Spelsberg, T.; Fraser, D.; Atkinson, E.; Arnold, R.; Khosla, S. Dose-Response of Estrogen on Bone versus the Uterus in Ovariectomized Mice. Eur. J. Endocrinol. 2004, 151, 503–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Hirata, M.; Tominari, T.; Matsumoto, C.; Endo, Y.; Murphy, G.; Nagase, H.; Inada, M.; Miyaura, C. BA321, a Novel Carborane Analog That Binds to Androgen and Estrogen Receptors, Acts as a New Selective Androgen Receptor Modulator of Bone in Male Mice. Biochem. Biophys. Res. Commun. 2016, 478, 279–285. [Google Scholar] [CrossRef]
- Omiya, T.; Hirose, J.; Omata, Y.; Tominari, T.; Inada, M.; Watanabe, H.; Miyamoto, T.; Tanaka, S. Sustained Anti-Osteoporotic Action of Risedronate Compared to Anti-RANKL Antibody Following Discontinuation in Ovariectomized Mice. Bone Rep. 2020, 13, 100289. [Google Scholar] [CrossRef] [PubMed]
- Harada, S.; Tominari, T.; Matsumoto, C.; Hirata, M.; Takita, M.; Inada, M.; Miyaura, C. Nobiletin, a Polymethoxy Flavonoid, Suppresses Bone Resorption by Inhibiting NFκB-Dependent Prostaglandin E Synthesis in Osteoblasts and Prevents Bone Loss Due to Estrogen Deficiency. J. Pharmacol. Sci. 2011, 115, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Tominari, T.; Hirata, M.; Matsumoto, C.; Inada, M.; Miyaura, C. Polymethoxy Flavonoids, Nobiletin and Tangeretin, Prevent Lipopolysaccharide-Induced Inflammatory Bone Loss in an Experimental Model for Periodontitis. J. Pharmacol. Sci. 2012, 119, 390–394. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, C.; Inoue, H.; Tominari, T.; Watanabe, K.; Hirata, M.; Miyaura, C.; Inada, M. Heptamethoxyflavone, a Citrus Flavonoid, Suppresses Inflammatory Osteoclastogenesis and Alveolar Bone Resorption. Biosci. Biotechnol. Biochem. 2014, 79, 155–158. [Google Scholar] [CrossRef]
- Matsumoto, S.; Tominari, T.; Matsumoto, C.; Yoshinouchi, S.; Ichimaru, R.; Watanabe, K.; Hirata, M.; Grundler, F.; Miyaura, C.; Inada, M. Effects of Polymethoxyflavonoids on Bone Loss Induced by Estrogen Deficiency and by LPS-Dependent Inflammation in Mice. Pharmaceuticals 2018, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Ohyama, Y.; Ito, J.; Kitano, V.J.; Shimada, J.; Hakeda, Y. The Polymethoxy Flavonoid Sudachitin Suppresses Inflammatory Bone Destruction by Directly Inhibiting Osteoclastogenesis Due to Reduced ROS Production and MAPK Activation in Osteoclast Precursors. PLoS ONE 2018, 13, e0191192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Sang, S.; Pan, M.-H.H.; Lai, C.-S.S.; Lo, C.-Y.Y.; Yang, C.S.; Ho, C.-T.T. Anti-Inflammatory Property of the Urinary Metabolites of Nobiletin in Mouse. Bioorganic Med. Chem. Lett. 2007, 17, 5177–5181. [Google Scholar] [CrossRef]
- Guo, S.; Wu, X.; Zheng, J.; Smith, S.A.; Dong, P.; Xiao, H. Identification of 4′-Demethyltangeretin as a Major Urinary Metabolite of Tangeretin in Mice and Its Anti-Inflammatory Activities. J. Agric. Food Chem. 2021, 69, 4381–4391. [Google Scholar] [CrossRef]
- Kawahata, I.; Suzuki, T.; Rico, E.G.; Kusano, S.; Tamura, H.; Mimaki, Y.; Yamakuni, T. Fermented Citrus Reticulata (Ponkan) Fruit Squeezed Draff That Contains a Large Amount of 4′-Demethylnobiletin Prevents MK801-Induced Memory Impairment. J. Nat. Med. 2017, 71, 617–631. [Google Scholar] [CrossRef]
- Wu, X.; Song, M.; Rakariyatham, K.; Zheng, J.; Guo, S.; Tang, Z.; Zhou, S.; Xiao, H. Anti-Inflammatory Effects of 4′-Demethylnobiletin, a Major Metabolite of Nobiletin. J. Funct. Foods 2015, 19, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Misquitta, Y.R.; Olland, A.; Johnson, M.A.; Kelleher, K.S.; Kriz, R.; Lin, L.L.; Stahl, M.; Mosyak, L. Crystal Structure of a Human IκB Kinase β Asymmetric Dimer. J. Biol. Chem. 2013, 288, 22758–22767. [Google Scholar] [CrossRef] [Green Version]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An Open Chemical Toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Abu-Amer, Y. NF-ΚB Signaling and Bone Resorption. Osteoporos. Int. 2013, 24, 2377–2386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Meng, D.; Zhang, P.; Wang, X.; Du, G.; Brennan, C.; Li, S.; Ho, C.-T.; Zhao, H. Antioxidant Protection of Nobiletin, 5-Demethylnobiletin, Tangeretin, and 5-Demethyltangeretin from Citrus Peel in Saccharomyces Cerevisiae. J. Agric. Food Chem. 2018, 66, 3155–3160. [Google Scholar] [CrossRef]
- Hirata, N.; Tominari, T.; Ichimaru, R.; Taniguchi, K.; Matsumoto, C.; Watanabe, K.; Hirata, M.; Ma, S.; Suzuki, K.; Grundler, F.M.W.; et al. Structure-Activity Relationship of Anthocyanidins as an Inhibitory Effect on Osteoclast Differentiation. BPB Rep. 2019, 2, 1–6. [Google Scholar] [CrossRef]
- Hirata, N.; Ichimaru, R.; Tominari, T.; Matsumoto, C.; Watanabe, K.; Taniguchi, K.; Hirata, M.; Ma, S.; Suzuki, K.; Grundler, F.; et al. Beta-Cryptoxanthin Inhibits Lipopolysaccharide-Induced Osteoclast Differentiation and Bone Resorption via the Suppression of Inhibitor of NF-ΚB Kinase Activity. Nutrients 2019, 11, 368. [Google Scholar] [CrossRef] [Green Version]
- Murakami, A.; Song, M.; Katsumata, S.-I.; Uehara, M.; Suzuki, K.; Ohigashi, H. Citrus Nobiletin Suppresses Bone Loss in Ovariectomized DdY Mice and Collagen-Induced Arthritis in DBA/1J Mice: Possible Involvement of Receptor Activator of NF-KappaB Ligand (RANKL)-Induced Osteoclastogenesis Regulation. BioFactors 2007, 30, 179–192. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, J.; Ai, Z.; Su, J. Nobiletin-Loaded Micelles Reduce Ovariectomy-Induced Bone Loss by Suppressing Osteoclastogenesis. Int. J. Nanomed. 2019, 14, 7839–7849. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirata, M.; Tominari, T.; Ichimaru, R.; Takiguchi, N.; Tanaka, Y.; Takatoya, M.; Arai, D.; Yoshinouchi, S.; Miyaura, C.; Matsumoto, C.; et al. Effects of 4′-Demethylnobiletin and 4′-Demethyltangeretin on Osteoclast Differentiation In Vitro and in a Mouse Model of Estrogen-Deficient Bone Resorption. Nutrients 2023, 15, 1403. https://doi.org/10.3390/nu15061403
Hirata M, Tominari T, Ichimaru R, Takiguchi N, Tanaka Y, Takatoya M, Arai D, Yoshinouchi S, Miyaura C, Matsumoto C, et al. Effects of 4′-Demethylnobiletin and 4′-Demethyltangeretin on Osteoclast Differentiation In Vitro and in a Mouse Model of Estrogen-Deficient Bone Resorption. Nutrients. 2023; 15(6):1403. https://doi.org/10.3390/nu15061403
Chicago/Turabian StyleHirata, Michiko, Tsukasa Tominari, Ryota Ichimaru, Naruhiko Takiguchi, Yuki Tanaka, Masaru Takatoya, Daichi Arai, Shosei Yoshinouchi, Chisato Miyaura, Chiho Matsumoto, and et al. 2023. "Effects of 4′-Demethylnobiletin and 4′-Demethyltangeretin on Osteoclast Differentiation In Vitro and in a Mouse Model of Estrogen-Deficient Bone Resorption" Nutrients 15, no. 6: 1403. https://doi.org/10.3390/nu15061403
APA StyleHirata, M., Tominari, T., Ichimaru, R., Takiguchi, N., Tanaka, Y., Takatoya, M., Arai, D., Yoshinouchi, S., Miyaura, C., Matsumoto, C., Ma, S., Suzuki, K., Grundler, F. M. W., & Inada, M. (2023). Effects of 4′-Demethylnobiletin and 4′-Demethyltangeretin on Osteoclast Differentiation In Vitro and in a Mouse Model of Estrogen-Deficient Bone Resorption. Nutrients, 15(6), 1403. https://doi.org/10.3390/nu15061403