
The Effect of Exercise Prescription on the Human Gut Microbiota and Comparison between Clinical and Apparently Healthy Populations: A Systematic Review
 ,
, 
Abstract
:1. Introduction
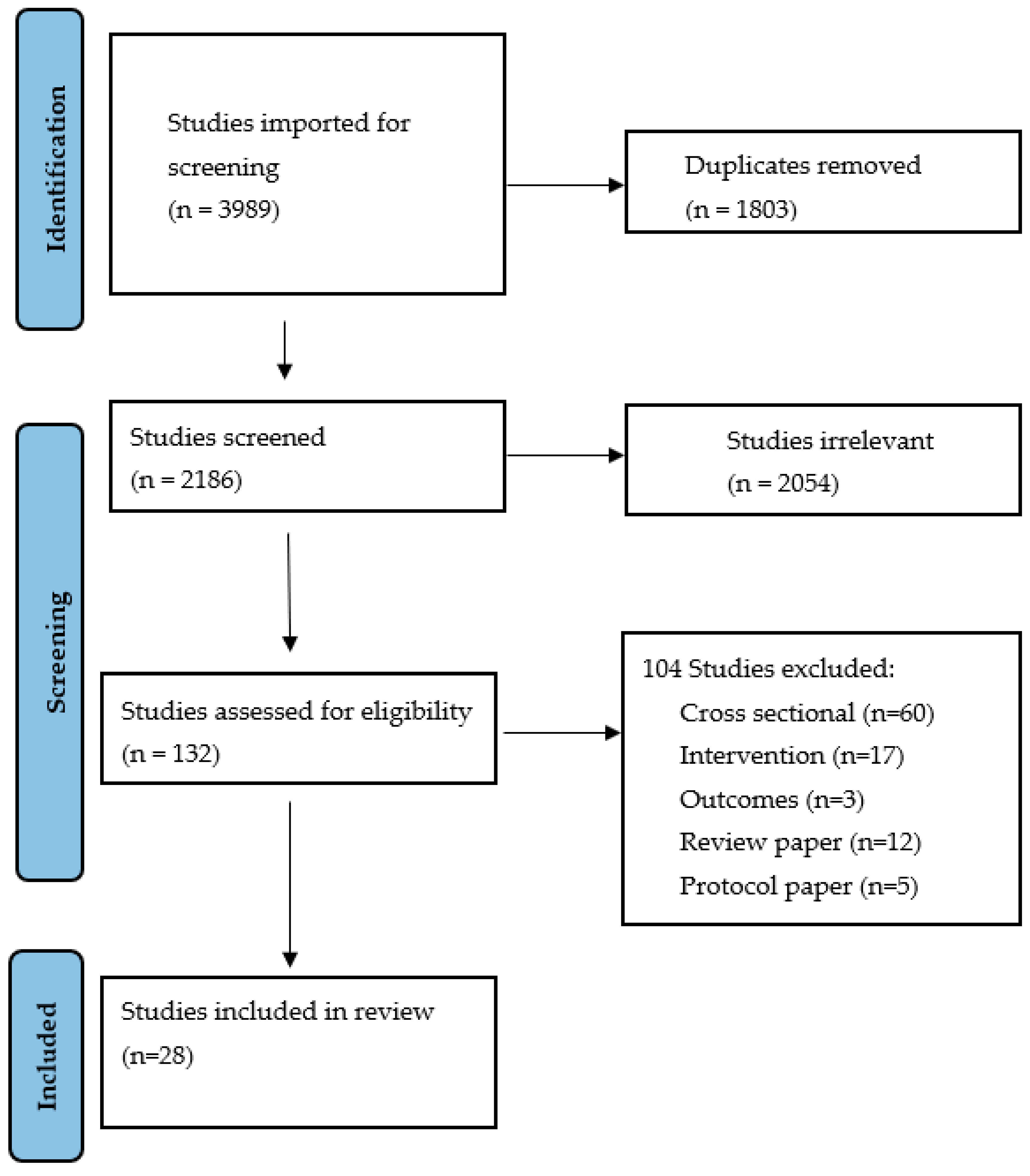
2. Methods
3. Results
4. Participant Characteristics
5. Intervention Characteristics
6. Outcome Measures
7. Influence of Exercise Type on the Gut Microbiota
8. Influence of Exercise Intensity on the Gut Microbiota
8.1. Low-to-Moderate and Moderate-Intensity Exercise
8.2. Moderate-to-High and High-Intensity Exercise
8.3. High Intensity Interval Training (HIIT)
8.4. Sprint Interval Training (SIT) and Maximal Effort
9. Influence of Exercise Frequency on the Gut Microbiota
10. Influence of Intervention Duration on the Gut Microbiota
11. Influence of Time Exercising per Session on the Gut Microbiota
12. Response of the Gut Microbiota to Exercise in Healthy Compared to Clinical Populations
13. Discussion
13.1. Exercise Intervention Characteristics
13.2. Population Influence
13.3. Limitations
14. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bianchi, F.; Duque, A.L.R.F.; Saad, S.M.I.; Sivieri, K. Gut microbiome approaches to treat obesity in humans. Appl. Microbiol. Biotechnol. 2018, 103, 1081–1094. [Google Scholar] [CrossRef] [PubMed]
- Grosicki, G.J.; Fielding, R.A.; Lustgarten, M.S. Gut Microbiota Contribute to Age-Related Changes in Skeletal Muscle Size, Composition, and Function: Biological Basis for a Gut-Muscle Axis. Calcif. Tissue Int. 2017, 102, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badal, V.D.; Vaccariello, E.D.; Murray, E.R.; Yu, K.E.; Knight, R.; Jeste, D.V.; Nguyen, T.T. The Gut Microbiome, Aging, and Longevity: A Systematic Review. Nutrients 2020, 12, 3759. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.A.; Hennet, T. Mechanisms and consequences of intestinal dysbiosis. Cell. Mol. Life Sci. 2017, 74, 2959–2977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beam, A.; Clinger, E.; Hao, L. Effect of Diet and Dietary Components on the Composition of the Gut Microbiota. Nutrients 2021, 13, 2795. [Google Scholar] [CrossRef]
- O’Sullivan, O.; Cronin, O.; Clarke, S.F.; Murphy, E.F.; Molloy, M.G.; Shanahan, F.; Cotter, P. Exercise and the microbiota. Gut Microbes 2015, 6, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef] [Green Version]
- Barton, W.; Penney, N.C.; Cronin, O.; Garcia-Perez, I.; Molloy, M.G.; Holmes, E.; Shanahan, F.; Cotter, P.D.; O’Sullivan, O. The microbiome of professional athletes differs from that of more sedentary subjects in composition and particularly at the functional metabolic level. Gut 2018, 67, 625–633. [Google Scholar] [CrossRef]
- Clarke, S.F.; Murphy, E.F.; O’Sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’Reilly, M.; Jeffery, I.B.; Wood-Martin, R.; et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920. [Google Scholar] [CrossRef] [Green Version]
- Castellanos, N.; Diez, G.G.; Antúnez-Almagro, C.; Bressa, C.; Bailén, M.; González-Soltero, R.; Pérez, M.; Larrosa, M. Key Bacteria in the Gut Microbiota Network for the Transition between Sedentary and Active Lifestyle. Microorganisms 2020, 8, 785. [Google Scholar] [CrossRef]
- Aya, V.; Flórez, A.; Perez, L.; Ramírez, J.D. Association between physical activity and changes in intestinal microbiota composition: A systematic review. PLoS ONE 2021, 16, e0247039. [Google Scholar] [CrossRef]
- Mitchell, C.M.; Davy, B.M.; Hulver, M.W.; Neilson, A.P.; Bennett, B.J.; Davy, K.P. Does Exercise Alter Gut Microbial Composition? A Systematic Review. Med. Sci. Sports Exerc. 2019, 51, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Alvarez, L.; Xu, H.; Martinez-Tellez, B. Influence of Exercise on the Human Gut Microbiota of Healthy Adults: A Systematic Review. Clin. Transl. Gastroenterol. 2020, 11, e00126. [Google Scholar] [CrossRef] [PubMed]
- Cataldi, S.; Bonavolontà, V.; Poli, L.; Clemente, F.M.; De Candia, M.; Carvutto, R.; Silva, A.F.; Badicu, G.; Greco, G.; Fischetti, F. The Relationship between Physical Activity, Physical Exercise, and Human Gut Microbiota in Healthy and Unhealthy Subjects: A Systematic Review. Biology 2022, 11, 479. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.; Gibson, G.R.; Walton, G.E.; Magistro, D.; Kinnear, W.; Hunter, K. Systematic Review of the Effects of Exercise and Physical Activity on the Gut Microbiome of Older Adults. Nutrients 2022, 14, 674. [Google Scholar] [CrossRef] [PubMed]
- Dziewiecka, H.; Buttar, H.S.; Kasperska, A.; Ostapiuk–Karolczuk, J.; Domagalska, M.; Cichoń, J.; Skarpańska-Stejnborn, A. Physical activity induced alterations of gut microbiota in humans: A systematic review. BMC Sports Sci. Med. Rehabilitation 2022, 14, 1–22. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Craven, J.; Cox, A.J.; Bellinger, P.; Desbrow, B.; Irwin, C.; Buchan, J.; McCartney, D.; Sabapathy, S. The influence of exercise training volume alterations on the gut microbiome in highly-trained middle-distance runners. Eur. J. Sport Sci. 2021, 22, 1222–1230. [Google Scholar] [CrossRef]
- Tabone, M.; Bressa, C.; García-Merino, J.A.; Moreno-Pérez, D.; Van, E.C.; Castelli, F.A.; Fenaille, F.; Larrosa, M. The effect of acute moderate-intensity exercise on the serum and fecal metabolomes and the gut microbiota of cross-country endurance athletes. Sci. Rep. 2021, 11, 3558. [Google Scholar] [CrossRef]
- Zeppa, S.D.; Amatori, S.; Sisti, D.; Gervasi, M.; Agostini, D.; Piccoli, G.; Pazienza, V.; Gobbi, P.; Rocchi, M.B.L.; Sestili, P.; et al. Nine weeks of high-intensity indoor cycling training induced changes in the microbiota composition in non-athlete healthy male college students. J. Int. Soc. Sports Nutr. 2021, 18, 74. [Google Scholar] [CrossRef]
- Karl, J.P.; Margolis, L.M.; Madslien, E.H.; Murphy, N.E.; Castellani, J.W.; Gundersen, Y.; Hoke, A.V.; Levangie, M.W.; Kumar, R.; Chakraborty, N.; et al. Changes in intestinal microbiota composition and metabolism coincide with increased intestinal permeability in young adults under prolonged physiological stress. Am. J. Physiol. Liver Physiol. 2017, 312, G559–G571. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhang, Z.; Hu, B.; Huang, W.; Yuan, C.; Zou, L. Response of Gut Microbiota to Metabolite Changes Induced by Endurance Exercise. Front. Microbiol. 2018, 9, 765. [Google Scholar] [CrossRef] [Green Version]
- Grosicki, G.J.; Durk, R.P.; Bagley, J.R. Rapid Gut Microbiome Changes in a World-Class Ultramarathon Runner. Physiol. Rep. 2019, 7, e14313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keohane, D.M.; Woods, T.; O’Connor, P.; Underwood, S.; Cronin, O.; Whiston, R.; O’Sullivan, O.; Cotter, P.; Shanahan, F.; Molloy, M.G. Four men in a boat: Ultra-endurance exercise alters the gut microbiome. J. Sci. Med. Sport 2019, 22, 1059–1064. [Google Scholar] [CrossRef]
- Barton, W.; Cronin, O.; Garcia-Perez, I.; Whiston, R.; Holmes, E.; Woods, T.; Molloy, C.B.; Molloy, M.G.; Shanahan, F.; Cotter, P.D.; et al. The effects of sustained fitness improvement on the gut microbiome: A longitudinal, repeated measures case-study approach. Transl. Sports Med. 2020, 4, 174–192. [Google Scholar] [CrossRef]
- Oliveira, C.B.; Marques, C.; Abreu, R.; Figueiredo, P.; Calhau, C.; Brito, J.; Sousa, M. Gut microbiota of elite female football players is not altered during an official international tournament. Scand. J. Med. Sci. Sports 2021, 32 (Suppl. 1), 62–72. [Google Scholar] [CrossRef]
- Bycura, D.; Santos, A.; Shiffer, A.; Kyman, S.; Winfree, K.; Sutliffe, J.; Pearson, T.; Sonderegger, D.; Cope, E.; Caporaso, J. Impact of Different Exercise Modalities on the Human Gut Microbiome. Sports 2021, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Erlandson, K.M.; Liu, J.; Johnson, R.; Dillon, S.; Jankowski, C.M.; Kroehl, M.; Robertson, C.E.; Frank, D.N.; Tuncil, Y.; Higgins, J.; et al. An exercise intervention alters stool microbiota and metabolites among older, sedentary adults. Ther. Adv. Infect. Dis. 2021, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Resende, A.S.; Leite, G.S.F.; Junior, A.H.L. Changes in the Gut Bacteria Composition of Healthy Men with the Same Nutritional Profile Undergoing 10-Week Aerobic Exercise Training: A Randomized Controlled Trial. Nutrients 2021, 13, 2839. [Google Scholar] [CrossRef]
- Huber, Y.; Pfirrmann, D.; Gebhardt, I.; Labenz, C.; Gehrke, N.; Straub, B.K.; Ruckes, C.; Bantel, H.; Belda, E.; Clément, K.; et al. Improvement of non-invasive markers of NAFLD from an individualised, web-based exercise program. Aliment. Pharmacol. Ther. 2019, 50, 930–939. [Google Scholar] [CrossRef]
- Verheggen, R.J.H.M.; Konstanti, P.; Smidt, H.; Hermus, A.R.M.M.; Thijssen, D.H.J.; Hopman, M.T.E. Eight-week exercise training in humans with obesity: Marked improvements in insulin sensitivity and modest changes in gut microbiome. Obesity 2021, 29, 1615–1624. [Google Scholar] [CrossRef] [PubMed]
- Cronin, O.; Barton, W.; Skuse, P.; Penney, N.C.; Garcia-Perez, I.; Murphy, E.F.; Woods, T.; Nugent, H.; Fanning, A.; Melgar, S.; et al. A Prospective Metagenomic and Metabolomic Analysis of the Impact of Exercise and/or Whey Protein Supplementation on the Gut Microbiome of Sedentary Adults. mSystems 2018, 3, e00044-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, S.K.; Cook, D.; Meyer, J.; Vernon, S.D.; Le, T.; Clevidence, D.; Robertson, C.E.; Schrodi, S.; Yale, S.; Frank, D.N. Changes in Gut and Plasma Microbiome following Exercise Challenge in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). PLoS ONE 2015, 10, e0145453. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.M.; Mailing, L.J.; Niemiro, G.M.; Moore, R.; Cook, M.D.; White, B.A.; Holscher, H.D.; Woods, J.A. Exercise Alters Gut Microbiota Composition and Function in Lean and Obese Humans. Med. Sci. Sports Exerc. 2018, 50, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Morita, E.; Yokoyama, H.; Imai, D.; Takeda, R.; Ota, A.; Kawai, E.; Hisada, T.; Emoto, M.; Suzuki, Y.; Okazaki, K. Aerobic Exercise Training with Brisk Walking Increases Intestinal Bacteroides in Healthy Elderly Women. Nutrients 2019, 11, 868. [Google Scholar] [CrossRef] [Green Version]
- Rettedal, E.A.; Cree, J.M.E.; Adams, S.E.; MacRae, C.; Skidmore, P.M.L.; Cameron-Smith, D.; Gant, N.; Blenkiron, C.; Merry, T.L. Short-term high-intensity interval training exercise does not affect gut bacterial community diversity or composition of lean and overweight men. Exp. Physiol. 2020, 105, 1268–1279. [Google Scholar] [CrossRef]
- Taniguchi, H.; Tanisawa, K.; Sun, X.; Kubo, T.; Hoshino, Y.; Hosokawa, M.; Takeyama, H.; Higuchi, M. Effects of short-term endurance exercise on gut microbiota in elderly men. Physiol. Rep. 2018, 6, e13935. [Google Scholar] [CrossRef]
- Munukka, E.; Ahtiainen, J.P.; Puigbó, P.; Jalkanen, S.; Pahkala, K.; Keskitalo, A.; Kujala, U.M.; Pietilä, S.; Hollmén, M.; Elo, L.; et al. Six-Week Endurance Exercise Alters Gut Metagenome That Is not Reflected in Systemic Metabolism in Over-weight Women. Front. Microbiol. 2018, 9, 2323. [Google Scholar] [CrossRef] [Green Version]
- Cronin, O.; Barton, W.; Moran, C.; Sheehan, D.; Whiston, R.; Nugent, H.; McCarthy, Y.; Molloy, C.B.; O’Sullivan, O.; Cotter, P.D.; et al. Moderate-intensity aerobic and resistance exercise is safe and favorably influences body composition in patients with quiescent Inflammatory Bowel Disease: A randomized controlled cross-over trial. BMC Gastroenterol. 2019, 19, 29. [Google Scholar] [CrossRef]
- Kern, T.; Blond, M.B.; Hansen, T.H.; Rosenkilde, M.; Quist, J.S.; Gram, A.S.; Ekstrøm, C.T.; Hansen, T.; Stallknecht, B. Structured exercise alters the gut microbiota in humans with overweight and obesity—A randomized controlled trial. Int. J. Obes. 2019, 44, 125–135. [Google Scholar] [CrossRef]
- Motiani, K.K.; Collado, M.C.; Eskelinen, J.J.; Virtanen, K.A.; Löyttyniemi, E.; Salminen, S.; Nuutila, P.; Kalliokoski, K.K.; Hannukainen, J.C. Exercise training modulates gut microbiota profile and improves endotoxemia. Med. Sci. Sports Exerc. 2020, 52, 94–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warbeck, C.; Dowd, A.J.; Kronlund, L.; Parmar, C.; Daun, J.T.; Wytsma-Fisher, K.; Millet, G.Y.; Schick, A.; Reimer, R.A.; Fung, T.; et al. Feasibility and effects on the gut microbiota of a 12-week high-intensity interval training plus lifestyle education intervention on inactive adults with celiac disease. Appl. Physiol. Nutr. Metab. 2021, 46, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Dupuit, M.; Rance, M.; Morel, C.; Bouillon, P.; Boscaro, A.; Martin, V.; Vazeille, E.; Barnich, N.; Chassaing, B.; Boisseau, N. Effect of Concurrent Training on Body Composition and Gut Microbiota in Postmenopausal Women with Overweight or Obesity. Med. Sci. Sports Exerc. 2021, 54, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Mahdieh, M.S.; Maryam, J.; Bita, B.; Neda, F.; Motahare, M.; Mahboobeh, B.; LeBris S, Q.; Kalani Behrooz, S. A pilot study on the relationship between Lactobacillus, Bifidibactrium counts and inflammatory factors following exercise training. Arch. Physiol. Biochem. 2021, 1–10. [Google Scholar] [CrossRef]
- Mokhtarzade, M.; Shamsi, M.M.; Abolhasani, M.; Bakhshi, B.; Sahraian, M.A.; Quinn, L.S.; Negaresh, R. Home-based exercise training influences gut bacterial levels in multiple sclerosis. Complement. Ther. Clin. Pr. 2021, 45, 101463. [Google Scholar] [CrossRef]
- Jollet, M.; Nay, K.; Chopard, A.; Bareille, M.-P.; Beck, A.; Ollendorff, V.; Vernus, B.; Bonnieu, A.; Mariadassou, M.; Rué, O.; et al. Does Physical Inactivity Induce Significant Changes in Human Gut Microbiota? New Answers Using the Dry Immersion Hypoactivity Model. Nutrients 2021, 13, 3865. [Google Scholar] [CrossRef]
- Lin, C.-L.; Hsu, Y.-J.; Ho, H.-H.; Chang, Y.-C.; Kuo, Y.-W.; Yeh, Y.-T.; Tsai, S.-Y.; Chen, C.-W.; Chen, J.-F.; Huang, C.-C.; et al. Bifidobacterium longum subsp. Longum olp-01 supplementation during endurance running training improves exercise performance in middle-and long-distance runners: A double-blind controlled trial. Nutrients 2020, 12, 1972. [Google Scholar] [CrossRef]
- Jang, L.-G.; Choi, G.; Kim, S.-W.; Kim, B.-Y.; Lee, S.; Park, H. The combination of sport and sport-specific diet is associated with characteristics of gut microbiota: An observational study. J. Int. Soc. Sports Nutr. 2019, 16, 21. [Google Scholar] [CrossRef] [Green Version]
- Šket, R.; Deutsch, L.; Prevoršek, Z.; Mekjavić, I.B.; Plavec, J.; Rittweger, J.; Debevec, T.; Eiken, O.; Stres, B. Systems View of Deconditioning During Spaceflight Simulation in the PlanHab Project: The Departure of Urine 1 H-NMR Metabolomes From Healthy State in Young Males Subjected to Bedrest Inactivity and Hypoxia. Front. Physiol. 2020, 11, 532271. [Google Scholar] [CrossRef]
- Ticinesi, A.; Nouvenne, A.; Cerundolo, N.; Catania, P.; Prati, B.; Tana, C.; Meschi, T. Gut Microbiota, Muscle Mass and Function in Aging: A Focus on Physical Frailty and Sarcopenia. Nutrients 2019, 11, 1633. [Google Scholar] [CrossRef] [Green Version]
- Fielding, R.A.; Reeves, A.R.; Jasuja, R.; Liu, C.; Barrett, B.B.; Lustgarten, M.S. Muscle strength is increased in mice that are colonized with microbiota from high-functioning older adults. Exp. Gerontol. 2019, 127, 110722. [Google Scholar] [CrossRef]
- Scheiman, J.; Luber, J.M.; Chavkin, T.A.; MacDonald, T.; Tung, A.; Pham, L.-D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.T.; et al. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef] [PubMed]
- Bressa, C.; Bailén-Andrino, M.; Pérez-Santiago, J.; González-Soltero, R.; Pérez, M.; Montalvo-Lominchar, M.G.; Maté-Muñoz, J.L.; Domínguez, R.; Moreno, D.; Larrosa, M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 2017, 12, e0171352. [Google Scholar] [CrossRef] [Green Version]
- Al Bander, Z.; Nitert, M.D.; Mousa, A.; Naderpoor, N. The Gut Microbiota and Inflammation: An Overview. Int. J. Environ. Res. Public Health 2020, 17, 7618. [Google Scholar] [CrossRef]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef]
- Keirns, B.H.; Koemel, N.A.; Sciarrillo, C.M.; Anderson, K.L.; Emerson, S.R. Exercise and intestinal permeability: Another form of exercise-induced hormesis? Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 319, G512–G518. [Google Scholar] [CrossRef] [PubMed]
- Marchbank, T.; Davison, G.; Oakes, J.R.; Ghatei, M.A.; Patterson, M.; Moyer, M.P.; Playford, R.J. The nutriceutical bovine colostrum truncates the increase in gut permeability caused by heavy exercise in athletes. Am. J. Physiol. Liver Physiol. 2011, 300, G477–G484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuhl, M.N.; Lanphere, K.R.; Kravitz, L.; Mermier, C.M.; Schneider, S.; Dokladny, K.; Moseley, P.L. Effects of oral glutamine supplementation on exercise-induced gastrointestinal permeability and tight junction protein expression. J. Appl. Physiol. 2014, 116, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Pires, W.; Veneroso, C.E.; Wanner, S.P.; Pacheco, D.A.S.; Vaz, G.C.; Amorim, F.T.; Tonoli, C.; Soares, D.D.; Coimbra, C.C. Association between Exercise-Induced Hyperthermia and Intestinal Permeability: A Systematic Review. Sports Med. 2016, 47, 1389–1403. [Google Scholar] [CrossRef] [PubMed]
- van Wijck, K.; Lenaerts, K.; Van Loon, L.J.; Peters, W.H.; Buurman, W.A.; Dejong, C.H. Exercise-induced splanchnic hypoperfusion results in gut dysfunction in healthy men. PLoS ONE 2011, 6, e22366. [Google Scholar] [CrossRef] [Green Version]
- McKenna, Z.; Houck, J.; Ducharme, J.; Li, Z.; Berkemeier, Q.; Fennel, Z.; Wells, A.; Mermier, C.; Deyhle, M.; Laitano, O.; et al. The effect of prolonged interval and continuous exercise in the heat on circulatory markers of intestinal barrier integrity. Eur. J. Appl. Physiol. 2022, 122, 2651–2659. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, C.M.; Madigan, S.M.; Garcia-Perez, I.; Rankin, A.; Sullivan, O.O.; Cotter, P. Distinct microbiome composition and metabolome exists across subgroups of elite Irish athletes. J. Sci. Med. Sport 2019, 23, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef]
- Weersma, R.K.; Zhernakova, A.; Fu, J. Interaction between drugs and the gut microbiome. Gut 2020, 69, 1510–1519. [Google Scholar] [CrossRef]
- Nie, K.; Ma, K.; Luo, W.; Shen, Z.; Yang, Z.; Xiao, M.; Tong, T.; Yang, Y.; Wang, X. Roseburia intestinalis: A Beneficial Gut Organism From the Discoveries in Genus and Species. Front. Cell. Infect. Microbiol. 2021, 11, 757718. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Belenguer, A.; Holtrop, G.; Johnstone, A.M.; Flint, H.J.; Lobley, G.E. Reduced Dietary Intake of Carbohydrates by Obese Subjects Results in Decreased Concentrations of Butyrate and Butyrate-Producing Bacteria in Feces. Appl. Environ. Microbiol. 2007, 73, 1073–1078. [Google Scholar] [CrossRef] [Green Version]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottiere, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and functional importance in the gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef]
- Spiljar, M.; Merkler, D.; Trajkovski, M. The Immune System Bridges the Gut Microbiota with Systemic Energy Homeostasis: Focus on TLRs, Mucosal Barrier, and SCFAs. Front. Immunol. 2017, 8, 1353. [Google Scholar] [CrossRef] [Green Version]
- Hou, H.; Chen, D.; Zhang, K.; Zhang, W.; Liu, T.; Wang, S.; Dai, X.; Wang, B.; Zhong, W.; Cao, H. Gut microbiota-derived short-chain fatty acids and colorectal cancer: Ready for clinical translation? Cancer Lett. 2022, 526, 225–235. [Google Scholar] [CrossRef]
- Gloor, G.B.; Macklaim, J.M.; Pawlowsky-Glahn, V.; Egozcue, J.J. Microbiome Datasets Are Compositional: And This Is Not Optional. Front. Microbiol. 2017, 8, 2224. [Google Scholar] [CrossRef] [Green Version]
- Shanahan, F.; Ghosh, T.S.; O’Toole, P.W. The Healthy Microbiome—What Is the Definition of a Healthy Gut Microbiome? Gastroenterology 2021, 164, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Price, J.; Abu-Ali, G.; Huttenhower, C. The healthy human microbiome. Genome Med. 2016, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estaki, M.; Pither, J.; Baumeister, P.; Little, J.P.; Gill, S.K.; Ghosh, S.; Ahmadi-Vand, Z.; Marsden, K.R.; Gibson, D.L. Cardiorespiratory fitness as a predictor of intestinal microbial diversity and distinct metagenomic functions. Microbiome 2016, 4, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisanz, J.E.; Upadhyay, V.; Turnbaugh, J.A.; Ly, K.; Turnbaugh, P.J. Meta-Analysis Reveals Reproducible Gut Microbiome Alterations in Response to a High-Fat Diet. Cell Host Microbe 2019, 26, 265–272. [Google Scholar] [CrossRef]
- Mohajeri, M.H.; Brummer, R.J.M.; Rastall, R.A.; Weersma, R.K.; Harmsen, H.J.M.; Faas, M.; Eggersdorfer, M. The role of the microbiome for human health: From basic science to clinical applications. Eur. J. Nutr. 2018, 57 (Suppl. 1), 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.M. The Influence of the Gut Microbiota on Host Physiology: In Pursuit of Mechanisms. Yale J. Biol. Med. 2016, 89, 285–297. [Google Scholar]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef]


{kind=link}
| Reference | Study Design | Study Quality | Sample Size | Groups, Male/Female (%) | Age (Years) | BMI (kg/m2) | Condition/ Intervention Group | Control Group | Classification | Microbiota Change (Diversity and Relative Abundance of Genera) |
|---|---|---|---|---|---|---|---|---|---|---|
| Craven et al., 2021 [18] | Single-arm | 66.7% | 14 | Male 57% | F: 22 ± 3.4 M: 20.7 ± 3.2 | F: 20.0 M: 21.43 (calculated) | Middle-distance runners (competitive) | NA | Athlete | Alpha-diversity—NR Beta-diversity—NR ↓ Haemophilus |
| Tabone et al., 2021 [19] | Single-arm | 50% | 40 | Male 100% | 35.79 ± 8.01 | 22.75 ± 2.12 | Cross-country runners (elite) | NA | Athletes | ~Alpha-diversity ~Beta-diversity ↑ Blautia, Ruminococcus, Romboutsia ↓ Clostridium |
| Zeppa et al., 2021 [20] | Single-arm | 66.7% | 18 | Male 100% | 22 ± 2 | 22.3 ± 2.7 | Healthy (sedentary) | NA | Healthy | ~Alpha-diversity ↑ Beta-diversity ↑ Dorea, Ruminoccus, Bifidobacterium ↓ Roseburia, Prevotella |
| Karl et al., 2017 [21] | Randomised Controlled Trial (Single-arm for exercise) | 83.3% | 18 | NR | 19 ± 2 | 23.6 ± 1.8 | Healthy (military) | No: 73 participants in remainder of study | Military | ↑ Alpha-diversity Beta-diversity—NR ↑ Dorea, Ruminococcus, Streptococcus,Paraprevotella, Eggerthella, Akkermansia, Fusobacterium ↓ Roseburia, Lachnospira, Blautia, Blautia, Faecalibacterium, Odoribacter, Bacteroides, Collinsella |
| Zhao et al., 2018 [22] | Observational | 66.7% | 20 | Male 80% | 31.6 ± 6.1 | 22.6 ± 2.1 | Runners (amateur) | NA | Athletes | ~Alpha-diversity Beta-diversity—NR ↑ Ruminiclostridium, Coprococcus, Pseudobutyrivibrio, Ruminococcus, Romboutsia, Mitsuokella, Collinsella, Actinobacilus ↓ Roseburia |
| Grosicki et al., 2019 [23] | Observational | 50% | 1 | Male 100% | 32 | 22.14 | Ultra-marathon runner | NA | Athletes | ↑ Alpha-diversity Beta diversity—NR ↑ Faecalibacterium, Streptococcus, Veillonella, Haemophilus ↓ Subdoligranulum, Alloprevotella |
| Keohane et al., 2019 [24] | Observational | 50% | 4 | Male 100% | 25.5 ± 1.3 | 24.4 ± 1.4 | Endurance rowers | NA | Athletes | ↑ Alpha-diversity Beta-diversity—NR ↑ Roseburia, Dorea, Subdolilogranulum, Prevotella ↓ Bacteroides |
| Barton et al., 2020 * [25] | Observational | 83.3% | 2 | Male 100% | 31.5 (30–33) | 30.2 (28.6–31.7) | Marathon participant and triathlete | NA | Athlete | ↑ Alpha-diversity ↑ Beta-diversity ↑ Veillonella, Akkermansia, Bifidobacterium |
| Oliveira et al., 2022 [26] | Observational | 66.7% | 17 | Male 0% | 24.1 ± 3.4 | 21.4 ± 1.7 | Athletes (elite) | NA | Athletes | ~Alpha-diversity ~Beta-diversity |
| Bycura et al., 2021 [27] | Controlled trial | 83.3% | 56 | AT: 28 Male 25% RT: 28 Male 46% | AT: 20.54 ± 1.93 RT: 21.28 ± 3.85 | AT: 24.41 ± 4.20 RT: 23.77 ± 4.15 | Healthy young adults (Aerobic) | Healthy young adults (Resistance) | Healthy | Alpha-diveristy—NR AT: ↑ Beta-diversity |
| Erlandson et al., 2021 * [28] | Controlled trial | 83.3% | 22 | Male 95% | 58 (55, 63.8) | 27.4 (24.6, 31) | Healthy (sedentary) High-intensity exercise | Healthy (sedentary) Moderate intensity exercise | Healthy | ~ Alpha-diversity ↑ Beta-diversity ↑ Oscillospira, Bifidobacterium, Succinivibria ↓ Prevotella, Oribacter |
| Resende et al., 2021 [29] | Randomised Controlled Trial | 100% | 28 | I: 14 Male 100% C: 14 Male 100% | I: 25.58 ± 5.07 C: 25.5 ± 4.66 | I: 25.28 ± 4.11 C: 23.68 ± 3.29 | Healthy (sedentary) | Healthy (sedentary, no exercise intervention) | Healthy | ~Alpha-diversity ~Beta-diversity |
| Huber et al., 2019 * [30] | Single-arm | 66.7% | 44 | Male 65.9% | 41 (24–61) | 31.3 (27.3,34) | NAFLD | NA | Clinical | Alpha-diversity—NR ↑ Beta-diversity Abundance—NR |
| Verheggen et al., 2021 [31] | Single-arm | 66.7% | 14 | Male 50% | 51 ± 11 | 34.9 ± 4.9 | Obese (sedentary) | NA | Clinical | ~Alpha-diversity ~Beta-diversity ↑ Lachnospira, Ruminococcus |
| Cronin et al., 2018 * [32] | Randomised Controlled Trial (Single-arm for exercise) | 100% | 30 | Male 44% | 35 (28, 38) | 27.9 (25.1, 29.2) | Obese (sedentary) | No: 90 participants in remainder of study | Clinical | ↑ Alpha-diversity ↑ Beta-diversty |
| Shukla et al., 2015 [33] | Observational | 66.7% | 20 | ME/CFS: 10 Male 20% C: 10 Male 20% | ME: 48.6 ± 10.5 C: 46.5 ± 13 | ME: 23.9 ± 4.3 C: 24.6 ± 3.3 | Myalgic Encephalomyelitis/Chronic Fatigue Syndrome | Healthy Control | Clinical | Alpha-diversity—NR Beta-diversity—NR ↑ Lachnospira |
| Allen et al., 2018 [34] | Controlled trial | 83.3% | 32 | L: Male 50% O: Male 21% | L: 25.1 ± 6.52 O: 31.14 ± 8.57 | L: 22.21 ± 2.76 O: 35.71 ± 5.11 | Obese (sedentary) | Lean (sedentary) | Clinical | ~Alpha-diversity ↑ Beta-diversity O: ↑ Lachnospira, Bacteroides, Collinsella ↓ Faecalibacterium L: ↑ Lachnospira, Faecalibacterium, “Butyrate producers” ↓ Bacteroides |
| Morita et al., 2019 * [35] | Controlled trial | 100% | 32 | I: 15 Male 100% C: 14 Male 100% | I: 70 (66–75) C: 70 (66–77) | I: 21.7 (18.9–23.1) C: 20.6 (18.7–24) | Elderly women (sedentary) Aerobic exercise | Elderly women (Sedentary) Trunk exercise | Clinical | Alpha-diveristy—NR Beta-diversity—NR ↑ Bacteoides ↓ Closdrium |
| Rettedal et al., 2020 [36] | Controlled trial | 83.3% | 32 | I: 15 Male 100% C: 14 Male 100% | 20–45 | I: 29.6 ± 2.7 C: 22.7 ± 2.1 | Overweight men (sedentary) | Lean men | Clinical | ~Alpha-diversity ~Beta-diversity I: ↓ Subdoligranulum C: ↑ Subdoligranulum |
| Taniguchi et al., 2018 [37] | Randomised Controlled Trial | 83.3% | 33 | I: 15 Male 100% C: 17 Male 100% | 62–76 | I: 22.9 ± 2.5 C: 22.9 ± 2.5 | Diabetic (T2D) and pre diabetic | Diabetic (T2D) and prediabetic (Crossover) | Clinical | ~Alpha-diversity ~Beta-diversity ↑ Oscilllospira ↓ Clostridium |
| Munukka et al., 2018 [38] | Randomised Controlled Trial | 83.3% | 22 | Male 0% | 36.8 ± 3.9 | 31.8 ± 4.4 | Overweight (sedentary) | Overweight (sedentary) (Waitlist) | Clinical | ~Alpha-diversity ↑ Beta-diversity ↑ Dorea, Akkermansia ↓ Odoribacter |
| Cronin et al., 2019 * [39] | Randomised Controlled Trial | 83.3% | 17 | I: 8 Male 68.2% C: 9 Male 85.7% | I: 33 (31,36) C: 31 (31,36) | I: 28.1 (26.2, 32.4) C: 27.2 (24.5, 33.7) | Inflammatory bowel disease (Chron’s and UC) | Inflammatory bowel disease (crossover) | Clinical | ~Alpha-diversity ~beta-diversity |
| Kern et al., 2020 * [40] | Randomised Controlled Trial | 66.7% | 130 | Bike: 19 Male 42% Mod: 31 Male 55% Vig: 24 Male 50% C: 14 Male 57% | Bike: 35 (28, 43) Mod: 33 (27, 38) Vig: 39 (33, 42) C: 38 (30, 42) | Bike: 30.0 (28.3, 33.9) Mod: 29.3 (27.4, 30.5) Vig: 29.9 (28.2, 32.1) C: 29.9 (27.6, 32.3) | Overweight/obesity (sedentary) (Exercise Intensity x 3 groups) | Overweight/obesity (sedentary) (Usual care) | Clinical | ↑ Alpha-diversity ~Beta-diversity |
| Motiani et al., 2020 [41] | Randomised Controlled Trial | 66.7% | 26 | SIT: 13 MICT: 13 Male 61% | 40–55 | NR | Diabetic (T2D) and prediabetic (sedentary) SIT | Diabetic (T2D) and prediabetic (sedentary) MICT | Clinical | ~Alpha-diversity Beta-diversity—NR SIT: ↑ Lachnospira ↓ Blautia, Clostridium MICT: ↑ Faecalibacterium, Veillonella ↓ Blautia, Clostridium |
| Warbeck et al., 2020 [42] | Randomised Controlled Trial | 100% | 41 | I: 20 Male 20% C: 21 Male 10% | I: 42 ± 12.3 C: 36.2 ± 10.2 | I: 27.0 ± 5.2 C: 28.7 ± 6.1 | Celiac (sedentary) | Celiac (sedentary) waitlist | Clinical | ~Alpha-diversity ↑ Beta-diversity I: ↑ Roseburia, Adlercretzia C (waitlist): ↑ Veillonella, Bifidobacterium |
| Dupuit et al., 2021 [43] | Randomised Controlled Trial | 100% | 29 | I: 14 Male 0% C: 15 Male 0% | I: 58.8 ± 5.3 C: 60.9 ± 4.8 | I: 30.3 ± 3.5 C: 31.5 ± 3.4 | Post-menopausal women with overweight or obesity (sedentary) | Post-menopausal women with overweight or obesity (sedentary, no intervention) | Clinical | ~Alpha-diversity ↑ Beta-diversity |
| Mahdieh et al., 2021 [44] | Randomised Controlled Trial (pilot study) | 83.3% | 18 | I: 9 Male 0% C: 9 Male 0% | I: 23.87 ± 3.13 C: 26.37 ± 1.68 | I: 27.76 ± 1.60 C: 28.41 ± 2.81 | Overweight Women | Overweight Women (no exercise intervention) | Clinical | Alpha-diversity—NR Beta-diversity—NR I: ↑ Lactobacillus, Bifidobacterium C: ↑ Lactobacillus |
| Mokhtarzade et al., 2021 [45] | Randomised Controlled Trial | 83.3% | 42 | I: 21 Male 0% C: 21 Male 0% | I: 35.06 ± 8.18 C: 36.38 ± 9.13 | I: 23.47 ± 2.61 C: 22.62 ± 2.00 | Multiple Sclerosis | Multiple Sclerosis (no exercise intervention) | Clinical | Alpha-diversity—NR Beta-diveristy—NR I: ↑ Prevotella ~Bacteroides |
| Reference | Dropout Rate (%) | Aerobic/Resistance | Type | Duration of Intervention | Intensity | Time per Session | Frequency per Week | Adherence |
|---|---|---|---|---|---|---|---|---|
| Craven et al., 2021 [18] | NR | Aerobic | Running | 7 weeks | Reporte as volume: 3 weeks of normal training, 3 weeks of high-volume training (+30% training volume), one week taper | NR | Prescribed per participant | NR |
| Tabone et al., 2021 [19] | 0% | Aerobic | Treadmill and running | NA | Maximal intensity | Treadmill: until volitional fatigue Track: max pace 1 km | Single effort | 100% |
| Zeppa et al., 2021 [20] | 5.50% | Aerobic | Cycle ergometer | 9 weeks | HIIT mixed with LIT (each session had HI at 20% of session) | 55 min, 60 min, 70 min (3 weeks each) | 3× 55 min first 3 weeks, 4× 60 min 3 Weeks, 5× 70 min for last 3 weeks | NR |
| Karl et al., 2017 [21] | 0% | Aerobic | Cross-country ski/march | 4 days | 50:10 min work:rest | NR (51 km total distance) | NA | 100% |
| Zhao et al., 2018 [22] | 0% | Aerobic | Running | Single effort | Moderate to vigorous intensity | 92–160 min | NA | 100% |
| Grosicki et al., 2019 [23] | 0% | Aerobic (>80%) Resistance (<20%) | Running “Strength” | 23 weeks | Moderate to high | ~666 min per week | 115–124 km per week | 100% |
| Keohane et al., 2019 [24] | 0% | Aerobic | Rowing | 33 days, 22 h | Moderate to high | 2 h increments, totalling 349.9 h each | Average: 151.8 km/day (12 h) | 100% |
| Barton et al., 2020 [25] | 0% | Aerobic (n = 1) Concurrent (n = 1) | Sport specific | 26 weeks | NR | 1–8 h | NR | NR |
| Oliveira et al., 2022 [26] | NA | Sport specific | Sport specific | 3 days | 3–6 RPE | 666 min | 10 sessions over 3 days | NR |
| Bycura et al., 2021 [27] | 0% | AT: aerobic RT: resistance | AT: 2× group cycling sessions + 1× rotating CRE Activity RT: 3–6 sets of 6 12 reps full body exercise | 8 weeks | AT: 60–90% HRmax RT: 70–85% 1RM | 60 min | 3 sessions | 100% |
| Erlandson et al., 2021 [28] | 32% | Concurrent | Treadmill Four weight based exercises | 24 weeks | Periodised RT and AT until week 12 then randomised to moderate (40–50% VO2 max and 60–70% 1RM) or high intensity (60–70% VO2max and >80% 1RM) with same intervention structure | 20 min to 50 min AT, 4 exercises, 3 sets 8 reps | 3 sessions | NR |
| Resende et al., 2021 [29] | 14% | Aerobic | Cycle ergometer | 10 weeks | Moderate intensity (steady state weeks 1 and 2, 65% VO2 progressive load weekly for weeks 3–10) | 50 min | 3 | 100% compliance |
| Huber et al., 2019 [30] | 6.80% | Web based concurrent | AT: MICT, Treadmill interval RT: 10 Strength exercise | 8 weeks | Individualised moderate | NR | 3× per week for first 4 weeks. 5× per week for weeks 4–8 | 63.4% |
| Verheggen et al., 2021 [31] | 0% | Aerobic | Cycle ergometer | 8 weeks | 65–85% HRR (increased over 55 min intervention) | 55 min | 2–4 | 98% compliance |
| Cronin et al., 2018 [32] | 17% | Concurrent | AT: NR RT: 7 exercises | 8 weeks | AT: RPE 5–7/10 RT: >70% 1RM | NR | 3× per week | 88% |
| Shukla et al., 2015 [33] | 0% | Aerobic | Cycle ergometer | Single effort | Maximal intensity | ME = 11.72 ± 2.6 min C = 13.1 ± 3.4 min | NA | 100% |
| Allen et al., 2018 [34] | 22% | Aerobic | Cycle ergometer Treadmill | 6 weeks | 60–75% HRR | 30–60 min | 3 sessions | 100% Compliance |
| Morita et al., 2019 [35] | 9% | I = Aerobic C = ‘trunk muscle training’ | I: brisk walk C: trunk exercise | 12 weeks | I: >3 METs C: NR | 60 min per session | I: daily C: 1× group session per week + daily home sessions | I: 97.1% attendance C: >90% |
| Rettedal et al., 2020 [36] | 9% | Aerobic | Cycle ergometer | 3 weeks | High-intensity | 8–12 × 60 s bouts @ VO2 peak with 75 s recovery | 9 sessions in total on non-consecutive days | 100% |
| Taniguchi et al., 2018 [37] | 6% | Aerobic | Cycle ergometer | 5 weeks | 60–75% VO2peak | 30 min for weeks 1–2; 45 min for weeks 3–5 | 3 sessions | NR |
| Munukka et al., 2018 [38] | 11% | Aerobic (interval) | Cycle ergometer | 6 weeks | Low to moderate | 40–60 min | 3 sessions | NR |
| Cronin et al., 2019 [39] | 12% | Concurrent | AT: NR RT: 7 exercises | 8 weeks | AT: RPE 5–7/10 RT: >70% 1RM | NR | 3 sessions | 85% |
| Kern et al., 2020 [40] | 32% | Aerobic | Bike: bike commute Mod: NR Vig: NR C: Habitual living | 24 weeks | Bike: Not prescribed (commute) Mod: 50% VO2peak reserve Vig: 70% VO2peak reserve C: Not prescribed | Weekly energy expenditure of 1600 kcal for women and 2100 kcal for men | 5 sessions | 93% |
| Motiani et al., 2020 [41] | 19% | Aerobic | Cycle ergometer | 2 weeks | SIT: maximal effort interval MICT: 60% VO2peak | SIT: 4–6× 30 s bouts with 4 min recovery MICT: 40–60 min | 3 sessions | NR |
| Warbeck et al., 2020 [42] | 17% | Aerobic | Cycle ergometer Ellipticals Treadmills | 12 weeks | HIIT (30 s of vigorous effort followed by 2 min of recovery) | 60 min per session (HIIT for 15–35 min) | 2 sessions | 74.83% (attendance) |
| Dupuit et al., 2021 [43] | NR | Concurrent | Wattbike (HIIT) 10 resistance exercises (targeted whole body in circuit format) | 12 weeks | AT: >85% HRmax (8 s high, 12 s recovery) RT: 8–12 rep max | AT: 20 min RT: ~25 min | 3 sessions | 97.5% attendance 99% compliance |
| Mahdieh et al., 2021 [44] | 11% | Aerobic | Treadmill | 10 weeks | Moderate (55–60% HRR in week 1) gradually increasing to high intensity by week 10 (70–75% HRR) | 30 min in week one progressing to 45 min in week 10 | 3 sessions | 89% |
| Mokhtarzade et al., 2021 [45] | 17% | Concurrent | Aerobic: Jogging, running, cycling Resistance: Home based, 10 exercises | 6 months | AT: periodised 50–65% HRR to 60–75% HRR RT: Periodised RPE 5–6 to 7–8 | NR | 5× per week (2× RT, 3× Aer) | 90% |
| Author | Method of Analysis | Number of Samples per Participant | Sample Collection Timepoints (Weeks) | Dietary Control (for Study Period) | Dietary Assessment | Taxonomic Labelling Tool Used |
|---|---|---|---|---|---|---|
| Craven et al., 2021 [18] | 16S | 4 | 0 (×2), 6 (AFT), 7 (Taper) | No | 3-day diet diary at each testing point | NCBI database |
| Tabone et al., 2021 [19] | 16S | 2 | 0, <1 | No | FFQ and 3 × 24-h food diary recall | Silva reference database |
| Zeppa et al., 2021 [20] | 16S | 2 | 0, 9 | No | Daily diaries for duration of study (plus two weeks prior) | GreenGenes and UCLUST |
| Karl et al., 2017 [21] | 16S | 2 | 0, <1 | Yes | No | RDP classifier |
| Zhao et al., 2018 [22] | 16S | 2 | 0, <1 | Yes (type of food) | Questionnaire | Not Reported |
| Grosicki et al., 2019 [23] | 16S | 4 | 1, 19, 21 (after competition), 23 | No | No | PAST: Paleontological Statistics Software |
| Keohane et al., 2019 [24] | metagenomic | 4 | 0 (BEF), ~2, <5 (AFT), +3 months | No | FFQ (baseline), daily diet diary | MetaPhlAn2.0 |
| Barton et al., 2020 [25] | metagenomic | 14 | Fortnightly (0–26) | No | daily diary—My Fitness Pal App | MetaPhlAn2 database |
| Oliveira et al., 2022 [26] | 16S | 2 | 0, <1 | 24-h food records | Kraken taxonomy + Bracken custom data base (GutHealth_DB) | |
| Bycura et al., 2021 [27] | 16S | 28 | −3, 0 (BEF), 8 (AFT), 11 (two samples per week) | No | No | Bayes classifier in q2-feature Classifier Genome Taxonomy Database |
| Erlandson et al., 2021 [28] | 16S | 2 | 0, 24 | No | 3-day diet diary | SINA |
| Resende et al., 2021 [29] | 16S | 2 | 0, 10 | No | 48 h food record, FFQ and 3-day food diary | Greengenes |
| Huber et al., 2019 [30] | NR | 2 (n = 9) | 0, 8 | No | No | NR |
| Verheggen et al., 2021 [31] | 16S | 2 | 0, 8 | No (recommended to not change dietary pattern) | 24-h diary prior to sample, FFQ | NG-Tax |
| Cronin et al., 2018 [32] | metagenomic | 2 | 0, 8 | No | FFQ | Kraken taxonomy |
| Shukla et al., 2015 [33] | 16S | 3 | 0 (BEF), 48 h (AFT), 72 h | No | RDP classifier | |
| Allen et al., 2018 [34] | 16S | 3 | 0 (BEF), 6 (AFT), 12 weeks | 3-days prior to sample collection | 3-day food menu was followed prior to each faecal collection. Menu organised from 7-day diet diary | RDP classifier |
| Morita et al., 2019 [35] | 16S | 2 | 0, 12 | No | FFQ | Human Faecal Microbiota T RFLP profiling (10 groups) |
| Rettedal et al., 2020 [36] | 16S | 4 | 0 (x 2), 3 | Recommended to not change dietary pattern | FFQ | SILVA database v.132 |
| Taniguchi et al., 2018 [37] | 16S | 3 | 0, 5 (AFT), 10 | No | Yes (Diet history questionnaire) | UCLUST |
| Munukka et al., 2018 [38] | Metagenomic and 16S | 3 | 0, 6 (BEF), 12 (AFT) | No | 3-day food diary | Silva 123.4 database |
| Cronin et al., 2019 [39] | Metagenomic | 3 | 0, 8 (AFT), 16 | No | No | Kaiju taxonomic assignment |
| Kern et al., 2020 [40] | 16S | 3 | 0 (BEF), 12, 24 (AFT) | No | 3-day food diary | RDP classifier |
| Motiani et al., 2020 [41] | 16S | 2 | 0, 2 | No | No | Greengenes GG 13.8 Database |
| Warbeck et al., 2020 [42] | 16S | 3 | 0, 12 (AFT, BEF for WLC), 24 (Follow up, AFT for WLC) | No | 3-day diet diary | Silva 136 database |
| Dupuit et al., 2021 [43] | 16S | 2 | 0, 12 | Recommended to not change dietary pattern | 5-day food intake diary | Greengenes GG 13.8 Database |
| Mahdieh et al., 2021 [44] | 16S | 2 | 0, 10 | No | 72 h recall | Other: Targeted analysis of Lactobacillus and Bifidobacterium |
| Mokhtarzade et al., 2021 [45] | QPCR | 2 | 0, 26 | No | 72 h recall | Other: Targeted analysis of Prevotella, Akkermansia mucinophila, Faecalibacterium prausnitzii and Bacteroides |
| Findings and Recommendations | |
|---|---|
| Exercise to modify the gut microbiota |
|
| Population influence |
|
| Future research |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boytar, A.N.; Skinner, T.L.; Wallen, R.E.; Jenkins, D.G.; Dekker Nitert, M. The Effect of Exercise Prescription on the Human Gut Microbiota and Comparison between Clinical and Apparently Healthy Populations: A Systematic Review. Nutrients 2023, 15, 1534. https://doi.org/10.3390/nu15061534
Boytar AN, Skinner TL, Wallen RE, Jenkins DG, Dekker Nitert M. The Effect of Exercise Prescription on the Human Gut Microbiota and Comparison between Clinical and Apparently Healthy Populations: A Systematic Review. Nutrients. 2023; 15(6):1534. https://doi.org/10.3390/nu15061534
Chicago/Turabian StyleBoytar, Alexander N., Tina L. Skinner, Ruby E. Wallen, David G. Jenkins, and Marloes Dekker Nitert. 2023. "The Effect of Exercise Prescription on the Human Gut Microbiota and Comparison between Clinical and Apparently Healthy Populations: A Systematic Review" Nutrients 15, no. 6: 1534. https://doi.org/10.3390/nu15061534
APA StyleBoytar, A. N., Skinner, T. L., Wallen, R. E., Jenkins, D. G., & Dekker Nitert, M. (2023). The Effect of Exercise Prescription on the Human Gut Microbiota and Comparison between Clinical and Apparently Healthy Populations: A Systematic Review. Nutrients, 15(6), 1534. https://doi.org/10.3390/nu15061534