
Effect of Rotavirus Infection and 2′-Fucosyllactose Administration on Rat Intestinal Gene Expression

 , , , ,
, , , ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design and Sample Collection
2.3. RNA Extraction and Microarray Procedure
2.4. Microarray Data Analysis
2.5. Validation of Gene Expression by Real-Time PCR
2.6. Statistical Analysis
3. Results
3.1. Clinical Results
3.2. RV Effect on Overall Intestinal Rat Gene Expression
3.3. Gene Expression Changes Due to RV Infection and 2′-FL Supplementation
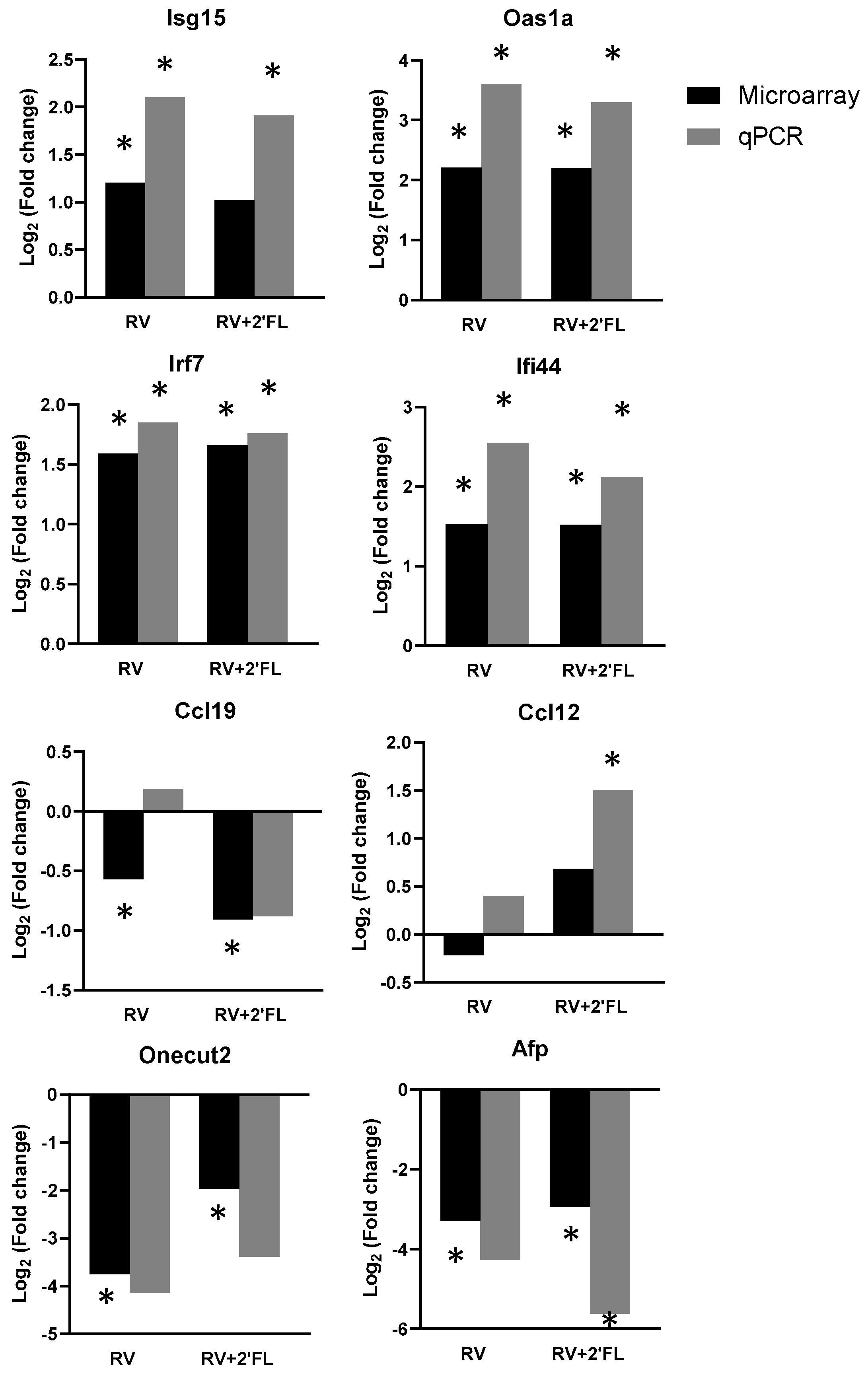
3.4. PCR Confirmation of Key Genes
3.5. Gene Expression Changes with 2′-FL Supplementation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Breastfeeding. Available online: https://www.who.int/health-topics/breastfeeding (accessed on 5 April 2022).
- Ballard, O.; Morrow, A.L. Human Milk Composition: Nutrients and Bioactive Factors. Pediatr. Clin. N. Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed]
- Rio-Aige, K.; Azagra-Boronat, I.; Castell, M.; Selma-Royo, M.; Collado, M.C.; Rodríguez-Lagunas, M.J.; Pérez-Cano, F.J. The Breast Milk Immunoglobulinome. Nutrients 2021, 13, 1810. [Google Scholar] [CrossRef] [PubMed]
- Picciano, M.F. Nutrient Composition of Human Milk. Pediatr. Clin. N. Am. 2001, 48, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Kirmiz, N.; Robinson, R.C.; Shah, I.M.; Barile, D.; Mills, D.A. Milk Glycans and Their Interaction with the Infant-Gut Microbiota. Annu. Rev. Food Sci. Technol. 2018, 9, 429–450. [Google Scholar] [CrossRef] [PubMed]
- Castanys-Muñoz, E.; Martin, M.J.; Prieto, P.A. 2’-Fucosyllactose: An Abundant, Genetically Determined Soluble Glycan Present in Human Milk. Nutr. Rev. 2013, 71, 773–789. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C.; Fente, C.; Regal, P.; Lamas, A.; Lorenzo, M.P. Human Milk Oligosaccharides (HMOs) and Infant Microbiota: A Scoping Review. Foods 2021, 10, 1429. [Google Scholar] [CrossRef] [PubMed]
- Hegar, B.; Wibowo, Y.; Basrowi, R.W.; Ranuh, R.G.; Sudarmo, S.M.; Munasir, Z.; Atthiyah, A.F.; Widodo, A.D.; Supriatmo; Kadim, M.; et al. The Role of Two Human Milk Oligosaccharides, 2’-Fucosyllactose and Lacto-N-Neotetraose, in Infant Nutrition. Pediatr. Gastroenterol. Hepatol. Nutr. 2019, 22, 330–340. [Google Scholar] [CrossRef]
- Sodhi, C.P.; Wipf, P.; Yamaguchi, Y.; Fulton, W.B.; Kovler, M.; Niño, D.F.; Zhou, Q.; Banfield, E.; Werts, A.D.; Ladd, M.R.; et al. The Human Milk Oligosaccharides 2’-Fucosyllactose and 6’-Sialyllactose Protect against the Development of Necrotizing Enterocolitis by Inhibiting Toll-like Receptor 4 Signaling. Pediatr. Res. 2021, 89, 91–101. [Google Scholar] [CrossRef]
- Azagra-Boronat, I.; Massot-Cladera, M.; Knipping, K.; van’t Land, B.; Stahl, B.; Garssen, J.; Rodríguez-Lagunas, M.J.; Franch, À.; Castell, M.; Pérez-Cano, F.J. Supplementation with 2′-FL and ScGOS/LcFOS Ameliorates Rotavirus-Induced Diarrhea in Suckling Rats. Front. Cell. Infect. Microbiol. 2018, 8, 372. [Google Scholar] [CrossRef]
- Goehring, K.C.; Marriage, B.J.; Oliver, J.S.; Wilder, J.A.; Barrett, E.G.; Buck, R.H. Similar to Those Who Are Breastfed, Infants Fed a Formula Containing 2′-Fucosyllactose Have Lower Inflammatory Cytokines in a Randomized Controlled Trial. J. Nutr. 2016, 146, 2559–2566. [Google Scholar] [CrossRef]
- Berger, B.; Porta, N.; Foata, F.; Grathwohl, D.; Delley, M.; Moine, D.; Charpagne, A.; Siegwald, L.; Descombes, P.; Alliet, P.; et al. Linking Human Milk Oligosaccharides, Infant Fecal Community Types, and Later Risk To Require Antibiotics. mBio 2020, 11, e03196-19. [Google Scholar] [CrossRef] [PubMed]
- Esona, M.D.; Gautam, R. Rotavirus. Clin. Lab Med. 2015, 35, 363–391. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.E.; Burton, A.H.; Boschi-Pinto, C.; Steele, A.D.; Duque, J.; Parashar, U.D. 2008 Estimate of Worldwide Rotavirus-Associated Mortality in Children Younger than 5 Years before the Introduction of Universal Rotavirus Vaccination Programmes: A Systematic Review and Meta-Analysis. Lancet Infect. Dis. 2012, 12, 136–141. [Google Scholar] [CrossRef]
- Frias, A.H.; Vijay-Kumar, M.; Gentsch, J.R.; Crawford, S.E.; Carvalho, F.A.; Estes, M.K.; Gewirtz, A.T. Intestinal Epithelia Activate Anti-Viral Signaling via Intracellular Sensing of Rotavirus Structural Components. Mucosal Immunol. 2010, 3, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Akira, S. Innate Immunity and Adjuvants. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2748–2755. [Google Scholar] [CrossRef]
- Morales-Ferré, C.; Azagra-Boronat, I.; Massot-Cladera, M.; Tims, S.; Knipping, K.; Garssen, J.; Knol, J.; Franch, À.; Castell, M.; Pérez-Cano, F.J.; et al. Preventive Effect of a Postbiotic and Prebiotic Mixture in a Rat Model of Early Life Rotavirus Induced-Diarrhea. Nutrients 2022, 14, 1163. [Google Scholar] [CrossRef]
- Hostetler, M.A.; Nakanishi, A.K.; Whiteman, P.J. Gastroenteritis: An Evidence Based Approach To Typical Vomiting, Diarrhea, and Dehydration. Pediatr. Emerg. Med. Pract. 2004, 1, 1–19. [Google Scholar]
- Pérez-Berezo, T.; Franch, A.; Ramos-Romero, S.; Castellote, C.; Pérez-Cano, F.J.; Castell, M. Cocoa-Enriched Diets Modulate Intestinal and Systemic Humoral Immune Response in Young Adult Rats. Mol. Nutr. Food Res. 2011, 55, S56–S66. [Google Scholar] [CrossRef]
- Massot-Cladera, M.; Franch, À.; Castell, M.; Pérez-Cano, F.J. Cocoa Polyphenols and Fiber Modify Colonic Gene Expression in Rats. Eur. J. Nutr. 2017, 56, 1871–1885. [Google Scholar] [CrossRef]
- Selga, E.; Pérez-Cano, F.J.; Franch, À.; Ramírez-Santana, C.; Rivero, M.; Ciudad, C.J.; Castellote, C.; Noé, V. Gene Expression Profiles in Rat Mesenteric Lymph Nodes upon Supplementation with Conjugated Linoleic Acid during Gestation and Suckling. BMC Genom. 2011, 12, 182. [Google Scholar] [CrossRef]
- Morales-Ferré, C.; Azagra-Boronat, I.; Massot-Cladera, M.; Franch, À.; Castell, M.; Rodríguez-Lagunas, M.J.; Pérez-Cano, F.J. Sexual Dimorphism Has Low Impact on the Response against Rotavirus Infection in Suckling Rats. Vaccines 2020, 8, 345. [Google Scholar] [CrossRef] [PubMed]
- Rigo-Adrover, M.; Saldaña-Ruíz, S.; van Limpt, K.; Knipping, K.; Garssen, J.; Knol, J.; Franch, A.; Castell, M.; Pérez-Cano, F.J. A Combination of ScGOS/LcFOS with Bifidobacterium Breve M-16V Protects Suckling Rats from Rotavirus Gastroenteritis. Eur. J. Nutr. 2017, 56, 1657–1670. [Google Scholar] [CrossRef] [PubMed]
- Azagra-Boronat, I.; Massot-Cladera, M.; Knipping, K.; Van‘T Land, B.; Tims, S.; Stahl, B.; Knol, J.; Garssen, J.; Franch, À.; Castell, M.; et al. Oligosaccharides Modulate Rotavirus-Associated Dysbiosis and TLR Gene Expression in Neonatal Rats. Cells 2019, 8, 876. [Google Scholar] [CrossRef] [PubMed]
- Amimo, J.O.; Raev, S.A.; Chepngeno, J.; Mainga, A.O.; Guo, Y.; Saif, L.; Vlasova, A.N. Rotavirus Interactions With Host Intestinal Epithelial Cells. Front. Immunol. 2021, 12, 793841. [Google Scholar] [CrossRef]
- Vlasova, A.N.; Rajashekara, G.; Saif, L.J. Interactions between Human Microbiome, Diet, Enteric Viruses and Immune System: Novel Insights from Gnotobiotic Pig Research. Drug Discov. Today Dis. Models 2018, 28, 95–103. [Google Scholar] [CrossRef]
- Montenegro-Landívar, M.F.; Tapia-Quirós, P.; Vecino, X.; Reig, M.; Valderrama, C.; Granados, M.; Cortina, J.L.; Saurina, J. Polyphenols and Their Potential Role to Fight Viral Diseases: An Overview. Sci. Total Environ. 2021, 801, 149719. [Google Scholar] [CrossRef]
- Collinson, S.; Padua-Zamora, A.; Gv, G.; Li, C.; Lf, D.; Sj, A. Cochrane Library Cochrane Database of Systematic Reviews Probiotics for Treating Acute Infectious Diarrhoea (Review). Cochrane. Database Syst. Rev. 2020, 12, CD003048. [Google Scholar] [CrossRef]
- Nogacka, A.M.; Arboleya, S.; Nikpoor, N.; Auger, J.; Salazar, N.; Cuesta, I.; Mantecón, L.; Solís, G.; Gueimonde, M.; Tompkins, T.A.; et al. Influence of 2’-Fucosyllactose on the Microbiota Composition and Metabolic Activity of Fecal Cultures from Breastfed and Formula-Fed Infants at Two Months of Age. Microorganisms 2021, 9, 1478. [Google Scholar] [CrossRef]
- Li, M.; Monaco, M.H.; Wang, M.; Comstock, S.S.; Kuhlenschmidt, T.B.; Fahey, G.C.; Miller, M.J.; Kuhlenschmidt, M.S.; Donovan, S.M. Human Milk Oligosaccharides Shorten Rotavirus-Induced Diarrhea and Modulate Piglet Mucosal Immunity and Colonic Microbiota. ISME J. 2014, 8, 1609. [Google Scholar] [CrossRef]
- Laucirica, D.R.; Triantis, V.; Schoemaker, R.; Estes, M.K.; Ramani, S. Milk Oligosaccharides Inhibit Human Rotavirus Infectivity in MA104 Cells. J. Nutr. 2017, 147, 1709. [Google Scholar] [CrossRef]
- Colbère-Garapin, F.; Martin-Latil, S.; Blondel, B.; Mousson, L.; Pelletier, I.; Autret, A.; François, A.; Niborski, V.; Grompone, G.; Catonnet, G.; et al. Prevention and Treatment of Enteric Viral Infections: Possible Benefits of Probiotic Bacteria. Microbes Infect. 2007, 9, 1623–1631. [Google Scholar] [CrossRef] [PubMed]
- Sargeant, H.R.; Miller, H.M.; Shaw, M.A. Inflammatory Response of Porcine Epithelial IPEC J2 Cells to Enterotoxigenic E. Coli Infection Is Modulated by Zinc Supplementation. Mol. Immunol. 2011, 48, 2113–2121. [Google Scholar] [CrossRef] [PubMed]
- Ingle, H.; Peterson, S.T.; Baldridge, M.T. Distinct Effects of Type I and III Interferons on Enteric Viruses. Viruses 2018, 10, 46. [Google Scholar] [CrossRef] [PubMed]
- Villena, J.; Vizoso-Pinto, M.G.; Kitazawa, H. Intestinal Innate Antiviral Immunity and Immunobiotics: Beneficial Effects against Rotavirus Infection. Front. Immunol. 2016, 7, 563. [Google Scholar] [CrossRef]
- Elkhateeb, E.; Tag-El-Din-Hassan, H.T.; Sasaki, N.; Torigoe, D.; Morimatsu, M.; Agui, T. The Role of Mouse 2′,5′-Oligoadenylate Synthetase 1 Paralogs. Infect. Genet. Evol. 2016, 45, 393–401. [Google Scholar] [CrossRef]
- Dediego, M.L.; Nogales, A.; Martinez-Sobrido, L.; Topham, D.J. Interferon-Induced Protein 44 Interacts with Cellular FK506-Binding Protein 5, Negatively Regulates Host Antiviral Responses, and Supports Virus Replication. mBio 2019, 10, e01839-19. [Google Scholar] [CrossRef]
- Lin, Z.; Wang, J.; Zhao, S.; Li, Y.; Zhang, Y.; Wang, Y.; Yan, Y.; Cheng, Y.; Sun, J. Goose IRF7 Is Involved in Antivirus Innate Immunity by Mediating IFN Activation. Dev. Comp. Immunol. 2022, 133, 104435. [Google Scholar] [CrossRef]
- Sherry, B. Rotavirus and Reovirus Modulation of the Interferon Response. J. Interferon Cytokine Res. 2009, 29, 559–567. [Google Scholar] [CrossRef]
- Santin, I.; Moore, F.; Grieco, F.A.; Marchetti, P.; Brancolini, C.; Eizirik, D.L. USP18 Is a Key Regulator of the Interferon-Driven Gene Network Modulating Pancreatic Beta Cell Inflammation and Apoptosis. Cell. Death Dis. 2012, 3, e419. [Google Scholar] [CrossRef]
- Malakhov, M.P.; Malakhova, O.A.; il Kim, K.; Ritchie, K.J.; Zhang, D.E. UBP43 (USP18) Specifically Removes ISG15 from Conjugated Proteins. J. Biol. Chem. 2002, 277, 9976–9981. [Google Scholar] [CrossRef]
- Ye, H.; Duan, X.; Yao, M.; Kang, L.; Li, Y.; Li, S.; Li, B.; Chen, L. USP18 Mediates Interferon Resistance of Dengue Virus Infection. Front. Microbiol. 2021, 12, 682380. [Google Scholar] [CrossRef] [PubMed]
- Malakhova, O.A.; Kim, K.I.; Luo, J.K.; Zou, W.; Kumar, K.G.S.; Fuchs, S.Y.; Shuai, K.; Zhang, D.E. UBP43 Is a Novel Regulator of Interferon Signaling Independent of Its ISG15 Isopeptidase Activity. EMBO J. 2006, 25, 2358–2367. [Google Scholar] [CrossRef] [PubMed]
- Carolina, D.; Palomino, T.; Marti, L.C. Chemokines and Immunity Quimiocinas e Imunidade. Einstein 2015, 13, 469–473. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, S.; Kou, L.; Tang, C.; Huang, R.; Pei, Z.; Li, Z. Ischemic Stroke Damages the Intestinal Mucosa and Induces Alteration of the Intestinal Lymphocytes and CCL19 MRNA in Rats. Neurosci. Lett. 2017, 658, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Kubo, Y.; Izumida, M.; Takahashi, E.; Kido, H.; Sato, K.; Yamaya, M.; Nishimura, H.; Nakayama, K.; Matsuyama, T. Enterokinase Enhances Influenza A Virus Infection by Activating Trypsinogen in Human Cell Lines. Front. Cell. Infect. Microbiol. 2018, 8, 91. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, D.; Fan, C.; Zhou, X.; Liu, Z.; Zheng, B.; Zhu, L.; Jin, Y. Novel Compound Heterozygous TMPRSS15 Gene Variants Cause Enterokinase Deficiency. Front. Genet. 2020, 11, 538778. [Google Scholar] [CrossRef]
- Shen, M.; Dong, C.; Ruan, X.; Yan, W.; Cao, M.; Pizzo, D.; Wu, X.; Yang, L.; Liu, L.; Ren, X.; et al. Chemotherapy-Induced Extracellular Vesicle MiRNAs Promote Breast Cancer Stemness by Targeting ONECUT2. Cancer Res. 2019, 79, 3608. [Google Scholar] [CrossRef] [PubMed]
- Kropp, P.A.; Gannon, M. Onecut Transcription Factors in Development and Disease. Trends. Dev. Biol. 2016, 9, 43–57. [Google Scholar]
- Christians, J.K.; Hoeflich, A.; Keightley, P.D. PAPPA2, an Enzyme That Cleaves an Insulin-Like Growth-Factor-Binding Protein, Is a Candidate Gene for a Quantitative Trait Locus Affecting Body Size in Mice. Genetics 2006, 173, 1547. [Google Scholar] [CrossRef]
- Andrew, M.; Liao, L.; Fujimoto, M.; Khoury, J.; Hwa, V.; Dauber, A. PAPPA2 as a Therapeutic Modulator of IGF-I Bioavailability: In Vivo and in Vitro Evidence. J. Endocr. Soc. 2018, 2, 646–656. [Google Scholar] [CrossRef]
- Azagra-Boronat, I.; Massot-Cladera, M.; Knipping, K.; Garssen, J.; ben Amor, K.; Knol, J.; Franch, À.; Castell, M.; Rodríguez-Lagunas, M.J.; Pérez-Cano, F.J. Strain-Specific Probiotic Properties of Bifidobacteria and Lactobacilli for the Prevention of Diarrhea Caused by Rotavirus in a Preclinical Model. Nutrients 2020, 12, 498. [Google Scholar] [CrossRef] [PubMed]
- Wong, Q.W.L.; Li, J.; Ng, S.R.; Lim, S.G.; Yang, H.; Vardy, L.A. RPL39L Is an Example of a Recently Evolved Ribosomal Protein Paralog That Shows Highly Specific Tissue Expression Patterns and Is Upregulated in ESCs and HCC Tumors. RNA Biol. 2014, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Sun, P.; Liu, X.-Y.; Dong, D.; Du, J.; Gu, L.; Ge Min Chen, Y.-B.; Yun Gang Higher Vocational, L.; Ge, Y.-B. α-Fetoprotein Involvement during Glucocorticoid-Induced Precocious Maturation in Rat Colon. Available World J. Gastroenterol. 2011, 17, 2933–2940. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (A) Upregulated | (B) Downregulated | ||||
|---|---|---|---|---|---|
| Gene | RV | RV+2′-FL | Gene | RV | RV+2′-FL |
| Oas1a | 2.21 | 2.04 | Ccl19 | −0.57 * | - |
| Oas1k | 2.1 | 1.87 | Slpi | −0.77 | −1.06 |
| Usp18 | 2.06 | 1.75 | Lpin1 | −0.82 | - |
| Zbp1 | 1.71 | 1.79 | Alpk3 | −0.83 | −1.02 |
| Irf7 | 1.59 | 1.51 | LOC103691469 | −0.84 | −1.08 |
| Tmigd1 | 1.53 | 1.89 | Kng2l1 | −0.85 | −1.01 |
| Ifi44 | 1.52 | 1.38 | Gkap1 | −0.87 | −0.96 |
| Cfb | 1.51 | 1.58 | S100g | −0.88 | −0.97 |
| Rpl39l | 1.45 * | - | Xcl1 | −0.9 | - |
| Dhx58 | 1.4 | 1.2 | Abca8a | −0.94 | −1.14 |
| Ifi27 | 1.32 | 1.44 | Gpcpd1 | −0.95 | −0.93 |
| LOC679368 | 1.24 | 1.89 | Apoa4 | −0.95 | −0.95 |
| Isg15 | 1.21 | - | Ribc2 | −0.96 | −0.9 |
| LOC690082 | 1.19 | 1.03 | Selenop | −0.97 | −0.96 |
| Aqp3 | 1.17 | 1.36 | Aoah | −1.01 | - |
| Capn3 | 1.12 | 1.16 | Cyp3a62 | −1.03 | −1.2 |
| Rasa4 | 1.11 | 1.07 | Sla2 | −1.03 | - |
| Tmem52 | 1.1 | - | Fcgrt | −1.08 | −1.21 |
| Fyb2 | 1.07 | 1.05 | LOC102555026 | −1.09 | −1.67 |
| Chdh | 1.07 | 1.24 | Igfals | −1.11 | −1.35 |
| Samd9 | 1.06 | 1 | Gpx3 | −1.13 | −1.32 |
| Upk1b | 1.02 | 1.21 | Ephb6 | −1.13 | −1.2 |
| Samd9 | 0.99 | - | Cd36 | −1.17 | −1.1 |
| Ces1e | 0.93 | 0.92 | LOC691352 | −1.2 | −1.11 |
| MGC108823 | 0.91 | 0.92 | Ankrd29 | −1.23 | −1.44 |
| Oas1i | 0.9 | - | Ptprr | −1.23 | −1.28 |
| Mcpt4 | 0.9 | - | Hoxc11 | −1.26 | −1.41 |
| LOC100912658 | 0.86 | - | Tacr3 | −1.31 | −1.63 |
| Lgals3bp | 0.83 | - | Srd5a2 | −1.32 | - |
| Slc37a4 | 0.73 | 0.83 | C8g | −1.52 | −1.53 |
| Dapl1 | −1.54 | −1.82 | |||
| Ggh | −1.58 | −1.49 | |||
| Slc3a1 | −1.63 | −1.09 | |||
| Pdx1 | −1.66 | −1.8 | |||
| Kcne1 | −2.33 | −2.63 | |||
| Aqp8 | −2.35 | −2.85 | |||
| RGD1562699 | −2.66 | - | |||
| Afp | −3.29 * | −5.39 | |||
| Onecut2 | −3.75 | −3.71 | |||
| Tmprss15 | −5.13 | −5.86 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sáez-Fuertes, L.; Azagra-Boronat, I.; Massot-Cladera, M.; Knipping, K.; Garssen, J.; Franch, À.; Castell, M.; Pérez-Cano, F.J.; Rodríguez-Lagunas, M.J. Effect of Rotavirus Infection and 2′-Fucosyllactose Administration on Rat Intestinal Gene Expression. Nutrients 2023, 15, 1996. https://doi.org/10.3390/nu15081996
Sáez-Fuertes L, Azagra-Boronat I, Massot-Cladera M, Knipping K, Garssen J, Franch À, Castell M, Pérez-Cano FJ, Rodríguez-Lagunas MJ. Effect of Rotavirus Infection and 2′-Fucosyllactose Administration on Rat Intestinal Gene Expression. Nutrients. 2023; 15(8):1996. https://doi.org/10.3390/nu15081996
Chicago/Turabian StyleSáez-Fuertes, Laura, Ignasi Azagra-Boronat, Malén Massot-Cladera, Karen Knipping, Johan Garssen, Àngels Franch, Margarida Castell, Francisco J. Pérez-Cano, and María J. Rodríguez-Lagunas. 2023. "Effect of Rotavirus Infection and 2′-Fucosyllactose Administration on Rat Intestinal Gene Expression" Nutrients 15, no. 8: 1996. https://doi.org/10.3390/nu15081996
APA StyleSáez-Fuertes, L., Azagra-Boronat, I., Massot-Cladera, M., Knipping, K., Garssen, J., Franch, À., Castell, M., Pérez-Cano, F. J., & Rodríguez-Lagunas, M. J. (2023). Effect of Rotavirus Infection and 2′-Fucosyllactose Administration on Rat Intestinal Gene Expression. Nutrients, 15(8), 1996. https://doi.org/10.3390/nu15081996