
Dietary Milk or Isolated Legume Proteins Modulate Intestinal Microbiota Composition in Rats


Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Purification
2.2. Animals and Diets
2.3. RT-qPCR Microbiota Composition Analysis
2.4. High-Throughput Analysis of Microbial Community
2.5. Analyses of Predicted Microbial Functions
2.6. Statistical Analysis
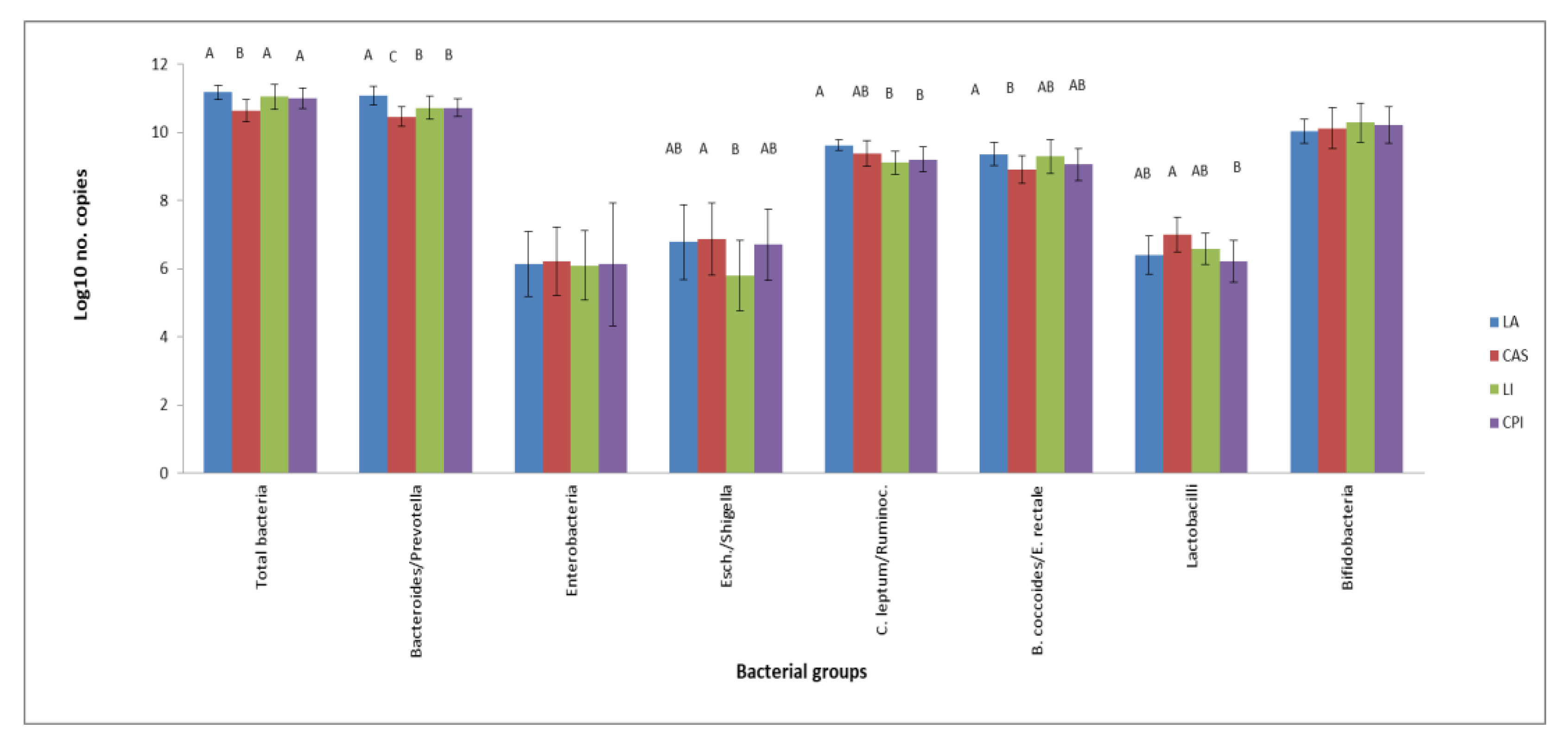
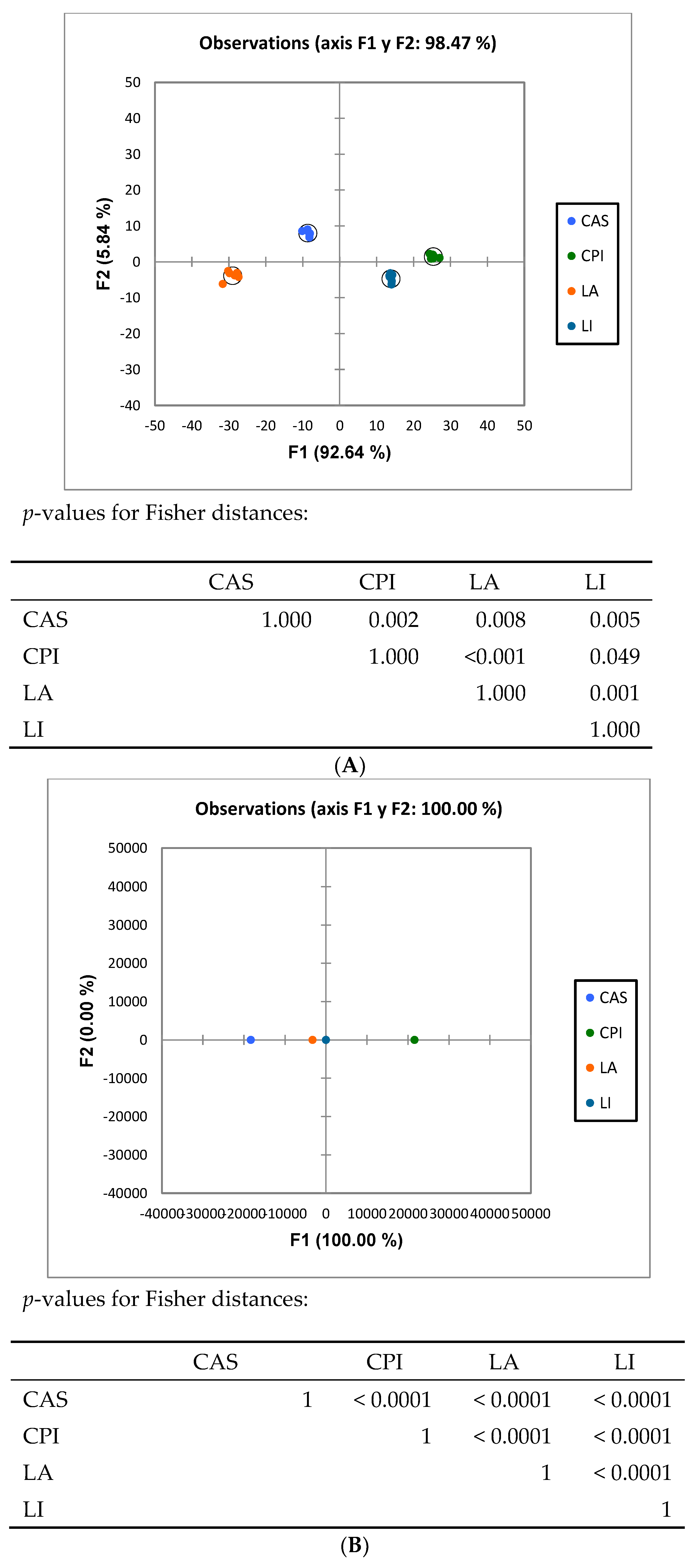
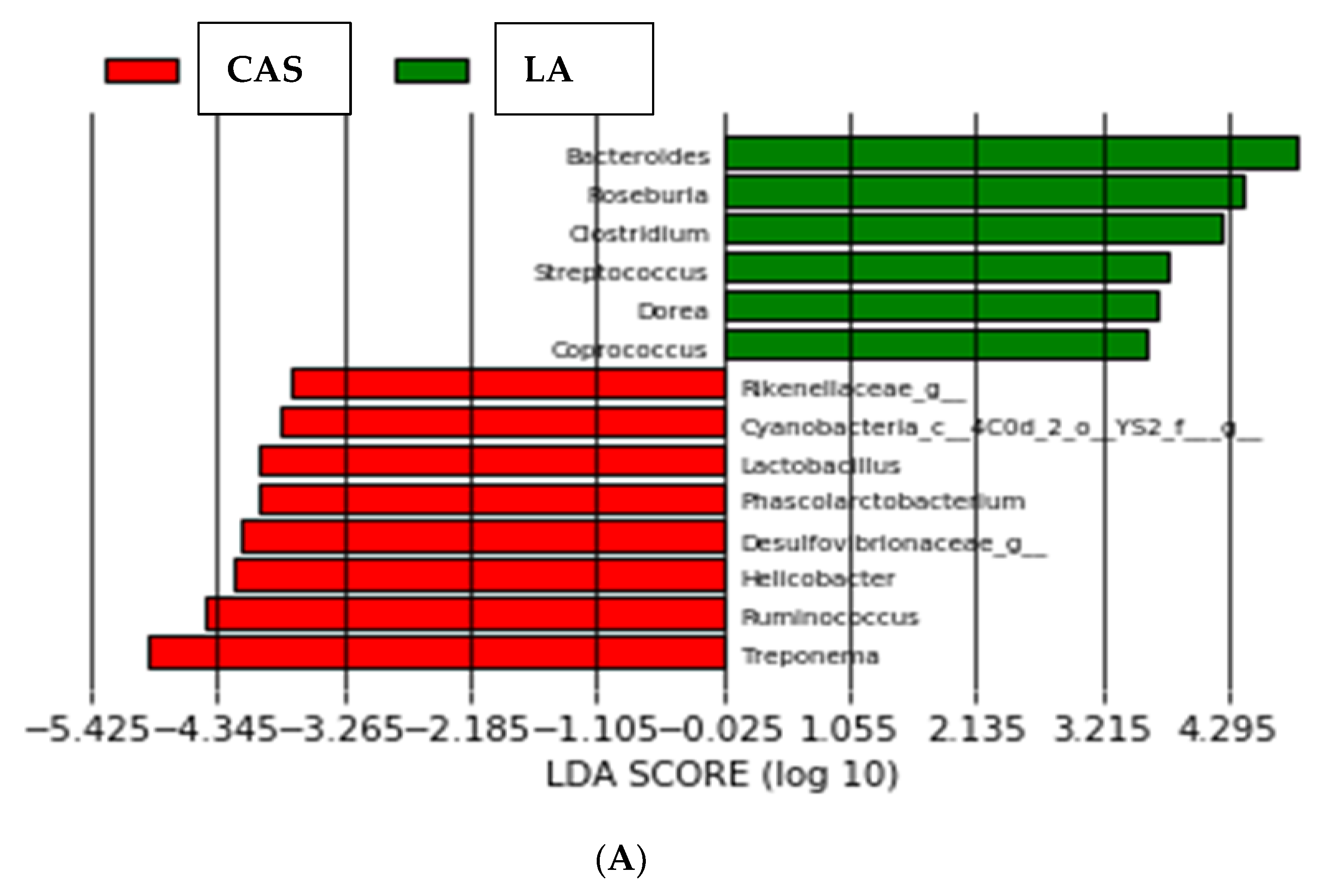
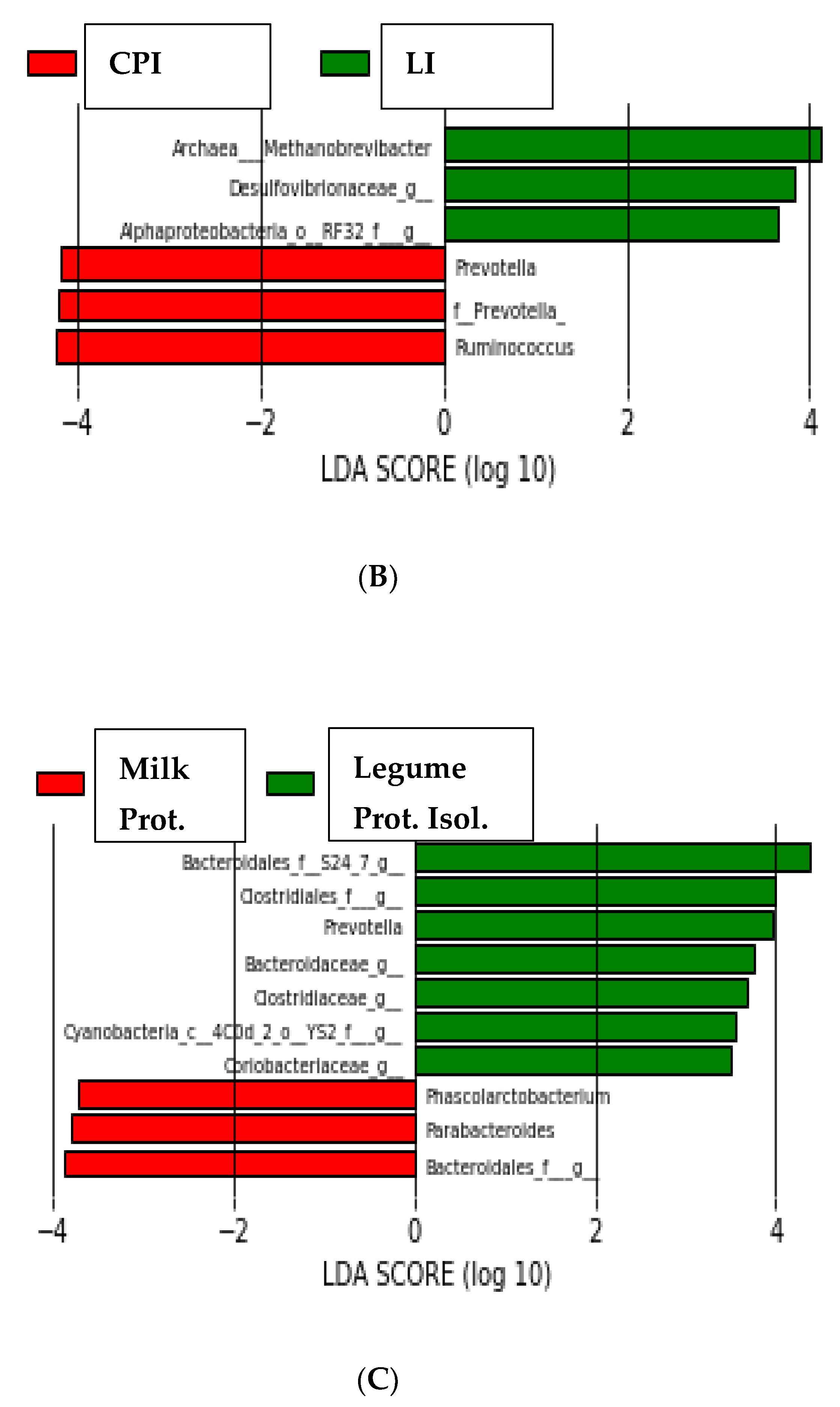
3. Results
3.1. RT-qPCR Microbiota Composition Analysis
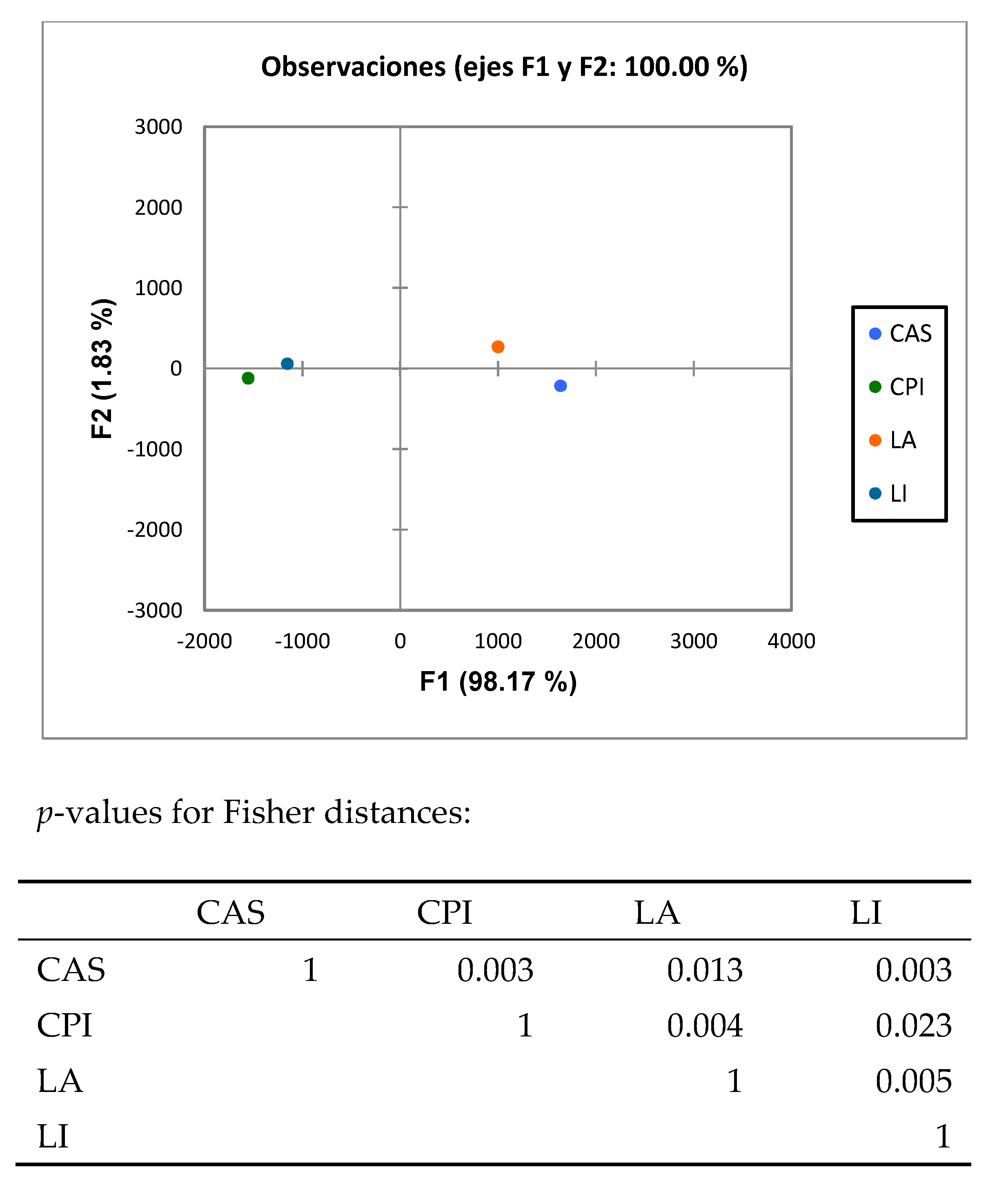
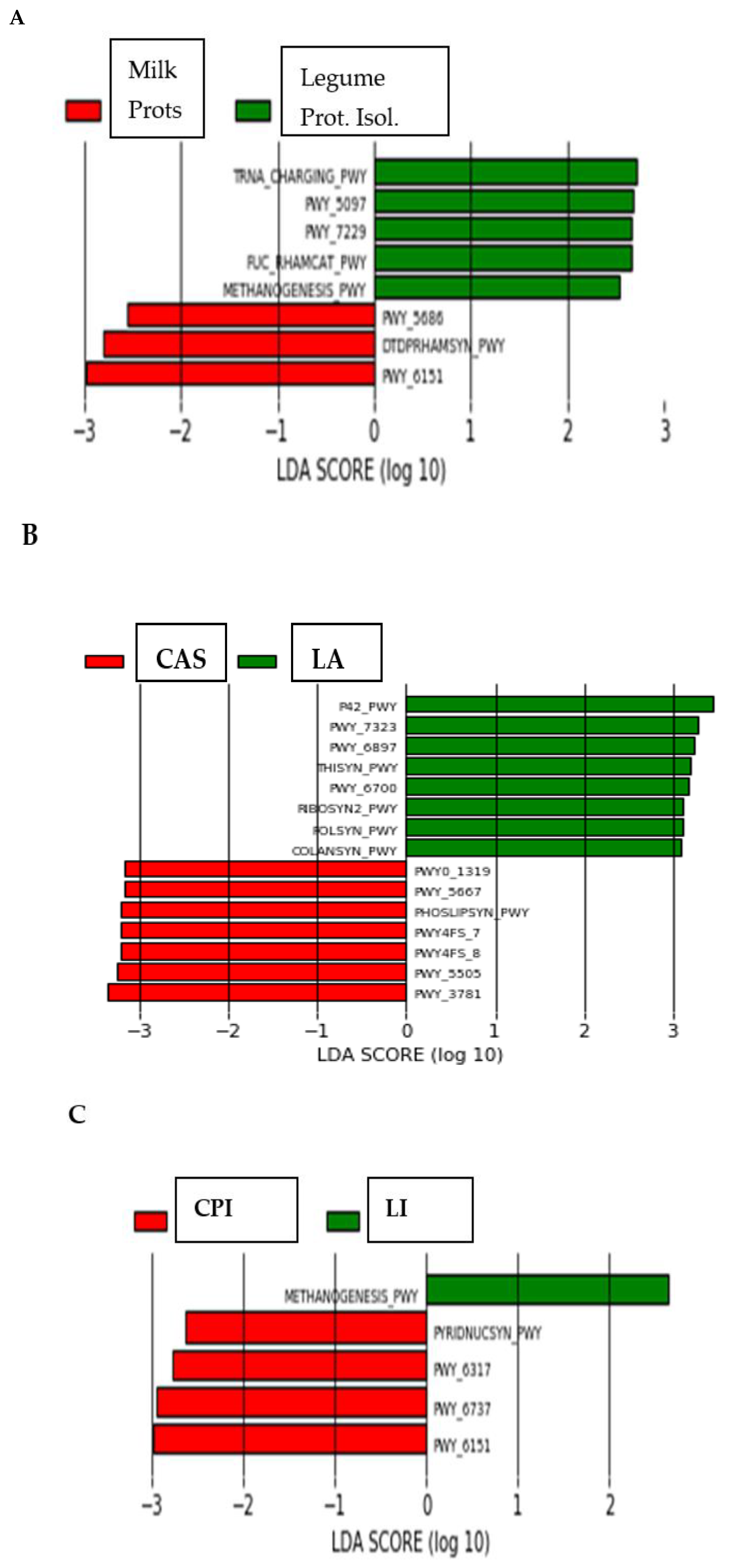
3.2. High-Throughput Analysis of Microbial Community
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Fan, P.; Li, L.; Rezaei, A.; Eslamfam, S.; Che, D.; Ma, X. Metabolites of dietary protein and peptides by intestinal microbes and their impacts on gut. Curr. Prot. Pept. Sci. 2015, 16, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Diether, N.; Willing, B. Microbial fermentation of dietary protein: An important factor in diet–microbe–host interaction. Microorganisms 2019, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Hector, A.J.; Phillips, S.M. Protein recommendations for weight loss in elite athletes: A focus on body composition and performance. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.A.; Clemente, A. In Vivo (rat) and in vitro (Caco-2 cells) absorption of amino acids from legume as compared to animal proteins. Arch. Anim. Nutr. 2009, 63, 413–426. [Google Scholar] [CrossRef]
- Gao, C.; Hou, L. Branched chain amino acids metabolism in heart failure. Front. Nutr. 2003, 10, 1279066. [Google Scholar] [CrossRef]
- Windey, K.; De Preter, V.; Verbeke, K. Relevance of protein fermentation to gut health. Mol. Nutr. Food Res. 2012, 56, 184–196. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef]
- Wu, S.; Bhat, Z.F.; Gounder, R.S.; Ahmed, I.A.M.; Al-Juhaimi, F.Y.; Ding, Y.; Bekhit, A.E.-D.A. Effect of dietary protein and processing on gut microbiota—A systematic review. Nutrients 2022, 14, 453. [Google Scholar] [CrossRef]
- Salonen, A.; de Vos, W.M. Impact of diet on human intestinal microbiota and health. Annu. Rev. Food Sci. Technol. 2014, 5, 239–562. [Google Scholar] [CrossRef]
- Sánchez-Vioque, R.; Clemente, A.; Vioque, J.; Bautista, J.; Millán, F. Protein isolates from chickpea (Cicer arietinum L.): Chemical composition, functional properties and protein characterization. Food Chem. 1999, 64, 237–243. [Google Scholar] [CrossRef]
- Ruiz, R.; Rubio, L.A. Lyophilisation improves the extraction of PCR-quality community DNA from pig faecal samples. J. Sci. Food Agric. 2009, 89, 723–727. [Google Scholar] [CrossRef]
- NRC. Nutrients Requirements of Laboratory Animals; National Academies Press: Washington, DC, USA, 1995. [Google Scholar]
- van Barneveld, R.J. Understanding the nutritional chemistry of lupin (Lupinus spp.) seed to improve livestock production efficiency. Nutr. Res. Rev. 1999, 12, 203–230. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.A.; Aranda-Olmedo, I.; Contreras, S.; Góngora, T.; Domínguez, G.; Peralta-Sánchez, J.M.; Martín-Pedrosa, M. Inclusion of limited amounts of extruded legumes plus cereal mixes in normocaloric or obesogenic diets for rats: Effects on intestinal microbiota composition. J. Sci. Food Agric. 2020, 100, 5546–5557. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.I.; Jesse, Z.; Gregory, J.; Daniel, M.D.; Dan, K.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S rRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 7, 5069–5072. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Sato, Y.; Furumichi, M.; Tanabe, M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res. 2012, 40, 109–114. [Google Scholar] [CrossRef]
- XLSTAT. Addinsoft. 2017. Available online: https://www.xlstat.com/es/soluciones/base (accessed on 10 December 2023).
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Smith, E.; Macfarlane, G. Enumeration of amino acid fermenting bacteria in the human large intestine: Effects of pH and starch on peptide metabolism and dissimilation of amino acids. FEMS Microbiol. Ecol. 1988, 25, 355–368. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, X.; Liu, H.; Brown, M.A.; Qiao, S. Dietary protein and gut microbiota composition and function. Curr. Prot. Pept. Sci. 2019, 20, 145. [Google Scholar] [CrossRef]
- Wu, S.; Bekhit, A.E.A.; Wu, Q.; Chen, M.; Liao, X.; Wang, J.; Ding, Y. Bioactive peptides and gut microbiota: Candidates for a novel strategy for reduction and control of neurodegenerative diseases. Trends Food Sci. Technol. 2021, 108, 164–176. [Google Scholar] [CrossRef]
- Fujio-Vejar, S.; Vasquez, Y.; Morales, P.; Magne, F.; Vera-Wolf, P.; Ugalde, J.A.; Navarrete, P.; Gotteland, M. The gut microbiota of healthy chilean subjects reveals a high abundance of the phylum Verrucomicrobia. Front. Microbiol. 2017, 8, 1221. [Google Scholar] [CrossRef] [PubMed]
- Antal, G.M.; Lukehart, S.A.; Meheus, A.Z. The endemic treponematoses. Microbes Infect. 2002, 4, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Chan, F.K.L.; Lau, J.Y.W. Gastrointestinal and Liver Disease, 11th ed.; Feldman, M., Friedman, L.S., Brandt, L.J., Eds.; Elsevier: Philadelphia, PA, USA, 2021; Chapter 53. [Google Scholar]
- Tavella, T.; Rampelli, S.; Guidarelli, G.; Bazzocchi, A.; Gasperini, C.; Pujos-Guillot, E.; Comte, B.; Barone, M.; Biagi, E.; Candela, M.; et al. Elevated gut microbiome abundance of Christensenellaceae, Porphyromonadaceae and Rikenellaceae is associated with reduced visceral adipose tissue and healthier metabolic profile in Italian elderly. Gut Microbes 2021, 13, 1880221. [Google Scholar] [CrossRef] [PubMed]
- Hoang, T.; Kim, M.J.; Park, J.W.; Jeong, S.-Y.; Lee, J.; Shin, A. Nutrition-wide association study of microbiome diversity and composition in colorectal cancer patients. BMC Cancer 2022, 22, 656. [Google Scholar] [CrossRef] [PubMed]
- Tipton, K.D. Efficacy and consequences of very-high-protein diets for athletes and exercisers. Proc. Nutr. Soc. 2011, 70, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rist, V.T.S.; Weiss, E.; Sauer, N.; Mosenthin, R.; Eklund, M. Effect of dietary protein supply originating from soybean meal or casein on the intestinal microbiota of piglets. Anaerobe 2014, 25, 72–79. [Google Scholar] [CrossRef]
- Rubio, L.A.; Peinado, M.J. Replacement of soybean meal with lupin or chickpea seed meal in diets for fattening Iberian pigs promotes a healthier ileal microbiota composition. Adv. Microbiol. 2014, 4, 498–503. [Google Scholar] [CrossRef]
- Kim, E.; Kim, D.; Park, J. Changes of mouse gut microbiota diversity and composition by modulating dietary protein and carbohydrate contents: A pilot study. Prev. Nutr. Food Sci. 2016, 21, 57–61. [Google Scholar] [CrossRef]
- Rubio, L.A.; Ruiz, R.; Peinado, M.J.; Martin-Pedrosa, M.; Fernández-Fígares, I.; Porres, J. Carbohydrates digestibility and faecal microbiota composition in rats fed diets based on raw or fermented Vigna unguiculata seed meal as the only protein source. Food Chem. Adv. 2022, 1, 100022. [Google Scholar]
- Davin-Regli, A.; Lavigne, J.P.; Pages, J.M. Enterobacter spp.: Update on Taxonomy, Clinical Aspects, and Emerging Antimicrobial Resistance. Clin. Microbiol. Rev. 2019, 32, e00002-19. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Utrilla, M.P.; Peinado, M.J.; Ruiz, R.; Rodríguez-Nogales, A.; Algieri, F.; Rodríguez-Cabezas, M.E.; Rubio, L.A. Pea (Pisum sativum L.) seed albumin extracts show anti-inflammatory effect in a DSS model of mouse colitis. Mol. Nutr. Food Res. 2015, 59, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Gaci, N.; Borrel, G.; Tottey, W.; O’Toole, P.W.; Brugère, J.F. Archaea and the human gut: New beginning of an old story. World J. Gastroenterol. 2014, 20, 16062–16078. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LA 1 | CAS | CPI | LI | |
|---|---|---|---|---|
| Asp | 98.4 | 58.9 | 92.7 | 93.7 |
| Glu | 142.9 | 184.1 | 123.3 | 185.3 |
| Ser | 39.6 | 49.1 | 39.1 | 43.9 |
| Gly | 18.9 | 16.5 | 27.9 | 34.6 |
| His | 17.2 | 27.5 | 19.1 | 21.4 |
| Arg | 28.5 | 37.4 | 58.9 | 80.4 |
| Thr | 45.3 | 39.6 | 26.4 | 30.3 |
| Ala | 51.2 | 32.6 | 33.5 | 30.7 |
| Pro | 51.7 | 82.8 | 38.9 | 44.5 |
| Tyr | 38.1 | 49.0 | 24.8 | 38.7 |
| Val | 55.2 | 53.3 | 38.5 | 38.9 |
| Met | 15.4 | 24.5 | 15.9 | 6.8 |
| Cys | 23.3 | 3.4 | 12.3 | 10.0 |
| Ile | 54.0 | 40.9 | 40.5 | 44.7 |
| Leu | 118.8 | 74.9 | 68.7 | 70.9 |
| Phe | 37.7 | 46.9 | 55.8 | 42.4 |
| Trp 2 | 16.7 | 10.2 | 5.5 | 8.8 |
| Lys | 89.3 | 97.9 | 50.4 | 41.6 |
| Diet 1 | ||||
|---|---|---|---|---|
| LA | CAS | LI | CPI | |
| Family | ||||
| LA | 0 | 0.0003 | 0.0014 | 0.0002 |
| CAS | 0 | 0.0076 | 0.0009 | |
| LPI | 0 | 0.0438 | ||
| CPI | 0 | |||
| Genus | ||||
| LA | 0 | 0.0007 | 0.0006 | 0.0004 |
| CAS | 0 | 0.0049 | 0.0003 | |
| LPI | 0 | 0.001 | ||
| CPI | 0 | |||
| Diet 1 | |||||
|---|---|---|---|---|---|
| LA 2 | CAS | CPI | LI | p-Values 3 | |
| Phylum | |||||
| Euryarchaeota | 2 b | 0 b | 4 b | 103 a | <0.0001 |
| Actinobacteria | 80 | 216 | 193 | 108 | 0.216 |
| Bacteroidetes | 2196 a | 1208 c | 1658 b | 1393 bc | <0.0001 |
| Firmicutes | 1480 b | 1562 ab | 1470 b | 1717 a | 0.058 |
| Proteobacteria | 177 b | 285 a | 183 b | 236 ab | 0.027 |
| Spirochaetes | 0 b | 661 a | 619 a | 420 a | <0.0001 |
| Family | |||||
| Methanobacteriaceae | 2 b | 0 b | 5 b | 100 a | 0.000 |
| Bifidobacteriaceae | 52 b | 216 a | 165 ab | 113 ab | 0.103 |
| Bacteroidales;f__ | 93 a | 76 a | 48 a | 39 a | 0.144 |
| Bacteroidaceae | 667 a | 179 b | 302 b | 324 b | 0.037 |
| Porphyromonadaceae | 45 | 56 | 37 | 31 | 0.184 |
| Prevotellaceae | 5 c | 0 c | 94 a | 35 b | 0.002 |
| Bacteroidales;f__S24-7 | 670 | 660 | 802 | 792 | 0.592 |
| [Paraprevotellaceae] | 386 a | 177 ab | 196 ab | 129 b | 0.070 |
| Cyanobacteria;f__ | 0 | 7 | 9 | 21 | 0.220 |
| Lactobacillaceae | 12 | 41 | 14 | 33 | 0.175 |
| Clostridiales;f__ | 70 b | 92 ab | 152 a | 142 a | 0.025 |
| Clostridiaceae | 98 a | 12 b | 20 b | 25 b | 0.015 |
| Lachnospiraceae | 342 ab | 139 b | 331 ab | 582 a | 0.013 |
| Ruminococcaceae | 773 b | 996 a | 745 bc | 619 c | 0.000 |
| Veillonellaceae | 90 b | 156 a | 90 b | 84 b | 0.007 |
| Erysipelotrichaceae | 77 | 52 | 102 | 116 | 0.204 |
| Alphaproteobacteria;o__RF32;f__ | 5 | 33 | 15 | 34 | 0.241 |
| Alcaligenaceae | 70 | 56 | 45 | 57 | 0.595 |
| Helicobacteraceae | 51 c | 173 a | 96 bc | 115 ab | 0.004 |
| Enterobacteriaceae | 6 a | 12 | 6 | 4 | 0.334 |
| Spirochaetaceae | 0 b | 6 a | 609 a | 428 a | <0.0001 |
| Genera | |||||
| Archaea;__Methanobrevibacter | 00 b | 0 b | 2 b | 71 a | <0.0001 |
| Bifidobacterium | 53 a | 159 a | 168 a | 97 a | 0.103 |
| Bacteroidales;f__;g__ | 104 a | 75 ab | 40 b | 60 ab | 0.134 |
| Bacteroides | 523 a | 193 b | 204 b | 300 b | 0.013 |
| Parabacteroides | 46 a | 55 a | 32 ab | 18 b | 0.028 |
| Prevotella | 0 b | 0 b | 115 a | 35 b | <0.0001 |
| Bacteroidales;f__S24-7;g__ | 676 | 663 | 803 | 784 | 0.642 |
| [Prevotella] | 1 b | 149 a | 191 a | 25 b | <0.0001 |
| Chloroflexi;c__S085;o__;f__;g__ | 24 b | 60 a | 3 b | 12 b | 0.013 |
| Cyanobacteria;c__4C0d;g__ | 0 b | 7 ab | 9 ab | 22 a | 0.143 |
| Lactobacillus | 12 a | 42 a | 14 a | 33 a | 0.128 |
| Clostridiales;f__;g__ | 63 b | 94 b | 152 a | 143 a | 0.002 |
| Lachnospiraceae;g__ | 75 a | 96 a | 204 a | 259 a | 0.256 |
| Blautia | 57 a | 21 b | 32 ab | 17 b | 0.069 |
| Coprococcus | 30 ab | 8 b | 31 ab | 47 a | 0.115 |
| Roseburia | 137 a | 10 b | 23 b | 17 b | <0.0001 |
| Ruminococcaceae;g__ | 306 bc | 240 c | 331 ab | 387 a | 0.006 |
| Oscillospira | 55 | 51 | 51 | 60 | 0.910 |
| Ruminococcus | 452 b | 651 a | 474 b | 226 c | <0.0001 |
| Phascolarctobacterium | 89 b | 147 a | 91 b | 83 b | 0.016 |
| Allobaculum | 73 | 49 | 97 | 114 | 0.197 |
| Alphaproteobacteria;o__RF32;f__;g_ | 3 b | 32 ab | 15 ab | 38 a | 0.105 |
| Sutterella | 69 | 56 | 45 | 58 | 0.626 |
| Helicobacteraceae;g__ | 22 | 24 | 54 | 51 | 0.235 |
| Helicobacter | 29 b | 136 a | 43 b | 30 b | <0.0001 |
| Treponema | 9 c | 644 a | 667 a | 436 b | <0.0001 |
| Diet 1 | |||||
|---|---|---|---|---|---|
| LA 2 | CAS | LI | CPI | p-Values 3 | |
| Family | |||||
| Simpson | 0.829 c | 0.842 bc | 0.865 a | 0.862 ab | 0.003 |
| Shannon | 2.070 c | 2.200 b | 2.330 a | 2.275 ab | 0.000 |
| Evenness | 0.480 | 0.482 | 0.505 | 0.501 | 0.743 |
| Chao1 | 16.571 c | 18.929 b | 20.375 a | 19.750 ab | <0.0001 |
| Genus | |||||
| Simpson | 0.854 | 0.874 | 0.877 | 0.880 | 0.136 |
| Shannon | 2.301 b | 2.438 a | 2.500 a | 2.469 a | 0.008 |
| Evenness | 0.479 | 0.498 | 0.487 | 0.493 | 0.933 |
| Chao1 | 21.143 c | 23.286 b | 25.125 a | 24.167 ab | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubio, L.A. Dietary Milk or Isolated Legume Proteins Modulate Intestinal Microbiota Composition in Rats. Nutrients 2024, 16, 149. https://doi.org/10.3390/nu16010149
Rubio LA. Dietary Milk or Isolated Legume Proteins Modulate Intestinal Microbiota Composition in Rats. Nutrients. 2024; 16(1):149. https://doi.org/10.3390/nu16010149
Chicago/Turabian StyleRubio, Luis A. 2024. "Dietary Milk or Isolated Legume Proteins Modulate Intestinal Microbiota Composition in Rats" Nutrients 16, no. 1: 149. https://doi.org/10.3390/nu16010149
APA StyleRubio, L. A. (2024). Dietary Milk or Isolated Legume Proteins Modulate Intestinal Microbiota Composition in Rats. Nutrients, 16(1), 149. https://doi.org/10.3390/nu16010149