
The Interplay between Endocrine-Disrupting Chemicals and the Epigenome towards Metabolic Dysfunction-Associated Steatotic Liver Disease: A Comprehensive Review

Abstract
:1. Introduction
2. Materials and Methods
3. Definitions and Types of EDCs
4. Relation between Dietary Habits, EDCs and MASLD
5. EDCs and Developmental Origins of MASLD
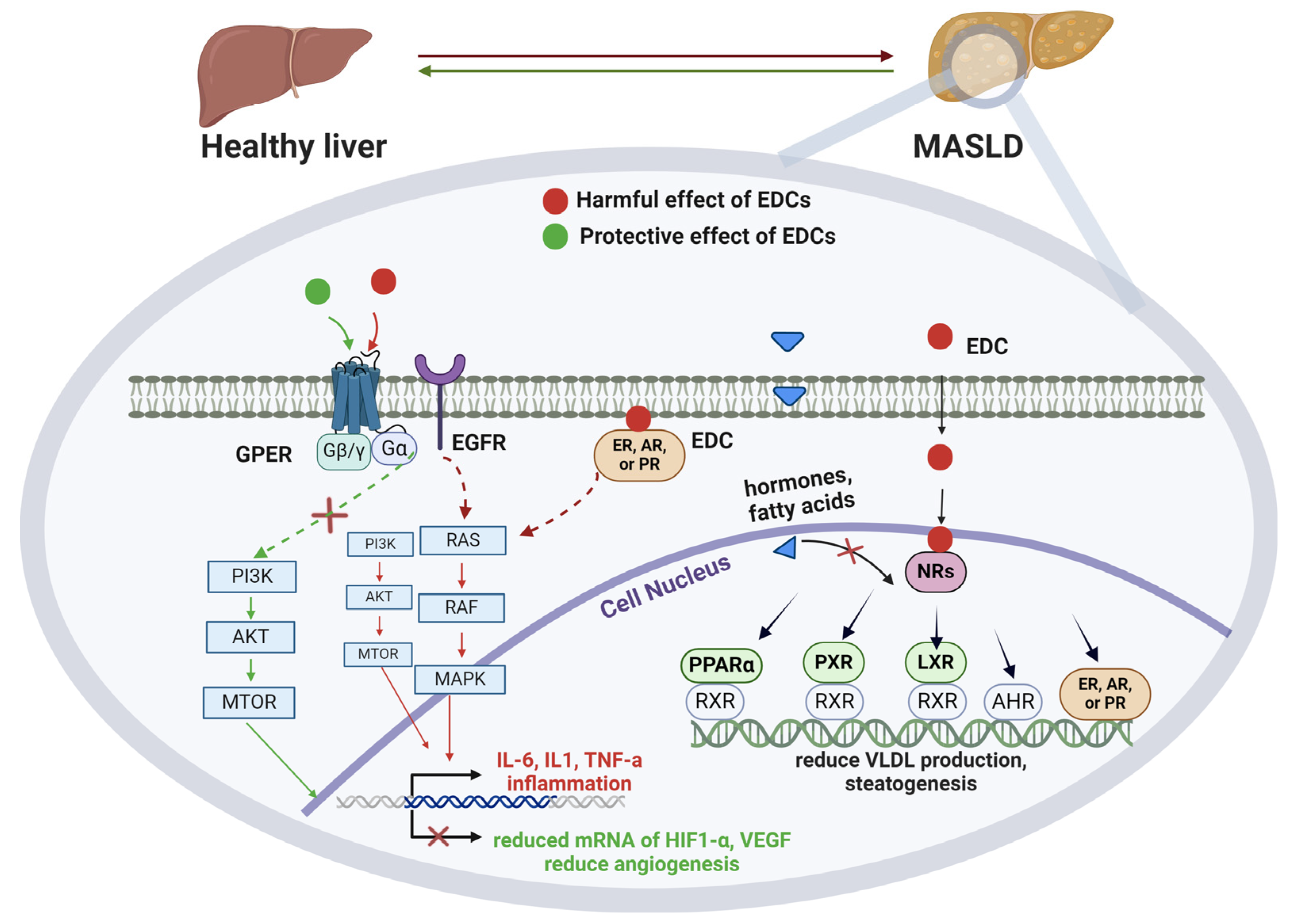
5.1. Molecular Mechanisms
5.2. Impact on Mitochondrial Function
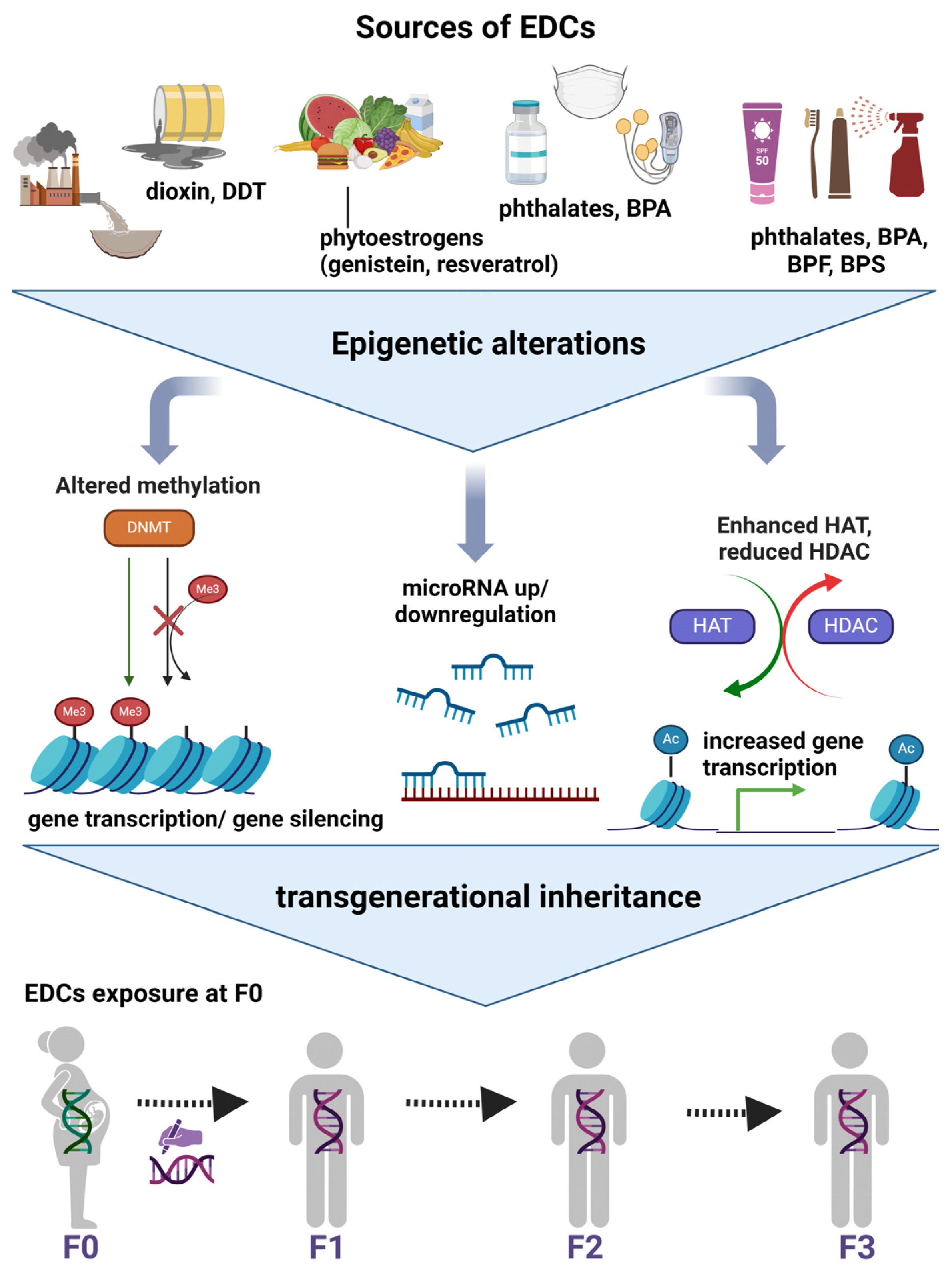
5.3. Epigenetic Regulations and Transgenerational Inheritance
5.4. EDCs and Their Role on MASLD Progression
5.5. Gene–EDCs Interactions in MASLD
6. Practical Recommendations for EDCs Exposure Reduction & MASLD Prevention
7. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MASLD | Metabolic-Dysfunction Associated Steatotic Liver Disease |
| EDCs | endocrine disrupting chemicals |
| NRs | nuclear receptors |
| NAFLD | non-alcoholic fatty liver disease |
| VLDL | very low-density lipoprotein |
| miRNA | microRNA |
| DDT | dichlorodiphenyltrichloroethane |
| BPA | bisphenol A |
| BPF | bisphenol F |
| BPS | bisphenol S |
| DES | diethylstilbestrol |
| PVC | plasticised polyvinyl chloride |
| DEHP | bis (2-ethylhexyl) phthalate |
| MedDiet | Mediterranean diet |
| DASH | Dietary approaches to stop Hypertension |
| NHANES | National Health and Nutrition Examination Survey |
| CI | Confidence Interval |
| TBT | tributyltin |
| PCBs | polychlorinated biphenyls |
| T2DM | type 2 diabetes melitus |
| IR | insulin resistance |
| PXR | pregnane X receptor |
| LXR | liver X receptor |
| AhR | aryl hydrocarbon receptor |
| PPARs | proliferator-activated receptors |
| ER | oestrogen receptor |
| AR | androgen receptor |
| PR | progesterone receptor |
| MAPK | protein kinase |
| PI3K | Phosphoinositide 3-kinase |
| PKB or AKT | Protein kinase B |
| GPER | G protein-coupled estrogen receptor |
| IL | interleukin |
| TNFa | tumor necrosis factor a |
| VEGF | vascular endothelial growth factor |
| ROS | reactive oxygen species |
| NAD | nicotinamide-adenine dinucleotide |
| EGFR | epidermal growth factor receptor |
| StAR | steroidogenic acute regulatory protein |
| DNMT | DNA methyltransferase |
| Stat3 | signal transducer and activator of transcription 3 |
| SREBF1 | sterol regulatory element binding transcription factor 1 |
| HAT | histone acetyltransferases |
| HDAC | histone deacetylases |
| ASD | autism spectrum disorder |
References
- Machado, M.V. MASLD Treatment—A Shift in the Paradigm Is Imminent. Front. Med. 2023, 10, 1316284. [Google Scholar] [CrossRef]
- Syed-Abdul, M.M. Lipid Metabolism in Metabolic-Associated Steatotic Liver Disease (MASLD). Metabolites 2024, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, B.; Terekeci, H.; Sandal, S.; Kelestimur, F. Endocrine Disrupting Chemicals: Exposure, Effects on Human Health, Mechanism of Action, Models for Testing and Strategies for Prevention. Rev. Endocr. Metab. Disord. 2020, 21, 127–147. [Google Scholar] [CrossRef]
- Foulds, C.E.; Treviño, L.S.; York, B.; Walker, C.L. Endocrine-Disrupting Chemicals and Fatty Liver Disease. Nat. Rev. Endocrinol. 2017, 13, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Plunk, E.C.; Richards, S.M. Epigenetic Modifications Due to Environment, Ageing, Nutrition, and Endocrine Disrupting Chemicals and Their Effects on the Endocrine System. Int. J. Endocrinol. 2020, 2020, 9251980. [Google Scholar] [CrossRef] [PubMed]
- Lei, R.; Xue, B.; Tian, X.; Liu, C.; Li, Y.; Zheng, J.; Luo, B. The Association between Endocrine Disrupting Chemicals and MAFLD: Evidence from NHANES Survey. Ecotoxicol. Environ. Saf. 2023, 256, 114836. [Google Scholar] [CrossRef]
- Zhou, Z.; Goodrich, J.M.; Strakovsky, R.S. Mitochondrial Epigenetics and Environmental Health: Making a Case for Endocrine Disrupting Chemicals. Toxicol. Sci. 2020, 178, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Balaguer, P.; Delfosse, V.; Grimaldi, M.; Bourguet, W. Structural and Functional Evidences for the Interactions between Nuclear Hormone Receptors and Endocrine Disruptors at Low Doses. Comptes Rendus Biol. 2017, 340, 414–420. [Google Scholar] [CrossRef]
- Beausoleil, C.; Ormsby, J.N.; Gies, A.; Hass, U.; Heindel, J.J.; Holmer, M.L.; Nielsen, P.J.; Munn, S.; Schoenfelder, G. Low Dose Effects and Non-Monotonic Dose Responses for Endocrine Active Chemicals: Science to Practice Workshop: Workshop Summary. Chemosphere 2013, 93, 847–856. [Google Scholar] [CrossRef]
- Heindel, J.J.; Blumberg, B.; Cave, M.; Machtinger, R.; Mantovani, A.; Mendez, M.A.; Nadal, A.; Palanza, P.; Panzica, G.; Sargis, R.; et al. Metabolism Disrupting Chemicals and Metabolic Disorders. Reprod. Toxicol. 2017, 68, 3–33. [Google Scholar] [CrossRef]
- Rao, A.; Douglas, S.C.; Hall, J.M. Endocrine Disrupting Chemicals, Hormone Receptors, and Acne Vulgaris: A Connecting Hypothesis. Cells 2021, 10, 1439. [Google Scholar] [CrossRef] [PubMed]
- Swedenborg, E.; Rüegg, J.; Mäkelä, S.; Pongratz, I. Endocrine Disruptive Chemicals: Mechanisms of Action and Involvement in Metabolic Disorders. J. Mol. Endocrinol. 2009, 43, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.L. Minireview: Epigenomic Plasticity and Vulnerability to EDC Exposures. Mol. Endocrinol. 2016, 30, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Janesick, A.S.; Shioda, T.; Blumberg, B. Transgenerational Inheritance of Prenatal Obesogen Exposure. Mol. Cell Endocrinol. 2014, 398, 31–35. [Google Scholar] [CrossRef]
- Ribeiro, E.; Ladeira, C.; Viegas, S. EDCs Mixtures: A Stealthy Hazard for Human Health? Toxics 2017, 5, 5. [Google Scholar] [CrossRef]
- Melough, M.M.; Maffini, M.V.; Otten, J.J.; Sathyanarayana, S. Diet Quality and Exposure to Endocrine-Disrupting Chemicals among US Adults. Environ. Res. 2022, 211, 113049. [Google Scholar] [CrossRef] [PubMed]
- Serrano, S.E.; Braun, J.; Trasande, L.; Dills, R.; Sathyanarayana, S. Phthalates and Diet: A Review of the Food Monitoring and Epidemiology Data. Environ. Health 2014, 13, 43. [Google Scholar] [CrossRef]
- Pacyga, D.C.; Sathyanarayana, S.; Strakovsky, R.S. Dietary Predictors of Phthalate and Bisphenol Exposures in Pregnant Women. Adv. Nutr. 2019, 10, 803–815. [Google Scholar] [CrossRef]
- Koch, H.M.; Lorber, M.; Christensen, K.L.Y.; Pälmke, C.; Koslitz, S.; Brüning, T. Identifying Sources of Phthalate Exposure with Human Biomonitoring: Results of a 48h Fasting Study with Urine Collection and Personal Activity Patterns. Int. J. Hyg. Env. Health 2013, 216, 672–681. [Google Scholar] [CrossRef]
- Rolfo, A.; Nuzzo, A.M.; De Amicis, R.; Moretti, L.; Bertoli, S.; Leone, A. Fetal–Maternal Exposure to Endocrine Disruptors: Correlation with Diet Intake and Pregnancy Outcomes. Nutrients 2020, 12, 1744. [Google Scholar] [CrossRef]
- Montemayor, S.; Mascaró, C.M.; Ugarriza, L.; Casares, M.; Llompart, I.; Abete, I.; Zulet, M.Á.; Martínez, J.A.; Tur, J.A.; Bouzas, C. Adherence to Mediterranean Diet and NAFLD in Patients with Metabolic Syndrome: The FLIPAN Study. Nutrients 2022, 14, 3186. [Google Scholar] [CrossRef] [PubMed]
- Anania, C.; Massimo Perla, F.; Olivero, F.; Pacifico, L.; Chiesa, C. Mediterranean Diet and Nonalcoholic Fatty Liver Disease. World J. Gastroenterol. 2018, 24, 2083–2094. [Google Scholar] [CrossRef]
- Tabb, M.M.; Blumberg, B. New Modes of Action for Endocrine-Disrupting Chemicals. Mol. Endocrinol. 2006, 20, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, N.; Plaz Torres, M.C.; Petta, S.; Valenti, L.; Giannini, E.G.; Aghemo, A. Is There an ‘Ideal’ Diet for Patients with NAFLD? Eur. J. Clin. Invest. 2022, 52, e13659. [Google Scholar] [CrossRef]
- Endocrine Society. What You Can Do about EDCs. Available online: https://www.endocrine.org/topics/edc/what-you-can-do (accessed on 10 March 2024).
- Rivas, A.; Monteagudo, C.; Heras-Gonzalez, L.; Mariscal-Arcas, M.; Lorenzo-Tovar, M.L.; Olea-Serrano, F. Association of Bisphenol A Exposure with Dietary Quality Indices in Spanish Schoolchildren. Food Chem. Toxicol. 2016, 94, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Van Woerden, I.; Payne-Sturges, D.C.; Whisner, C.M.; Bruening, M. Dietary Quality and Bisphenols: Trends in Bisphenol A, F, and S Exposure in Relation to the Healthy Eating Index Using Representative Data from the NHANES 2007–2016. Am. J. Clin. Nutr. 2021, 114, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Derghal, A.; Djelloul, M.; Trouslard, J.; Mounien, L. An Emerging Role of Micro-RNA in the Effect of the Endocrine Disruptors. Front. Neurosci. 2016, 10, 318. [Google Scholar] [CrossRef] [PubMed]
- Papalou, O.; Kandaraki, E.A.; Papadakis, G.; Diamanti-Kandarakis, E. Endocrine Disrupting Chemicals: An Occult Mediator of Metabolic Disease. Front. Endocrinol. 2019, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Xia, W.; Zhu, Y.; Li, X.; Wang, D.; Liu, J.; Chang, H.; Li, G.; Xu, B.; Chen, X.; et al. Mitochondrial Dysfunction in Early Life Resulted from Perinatal Bisphenol A Exposure Contributes to Hepatic Steatosis in Rat Offspring. Toxicol. Lett. 2014, 228, 85–92. [Google Scholar] [CrossRef]
- Zhou, X.; Afzal, S.; Zheng, Y.F.; Münch, G.; Li, C.G. Synergistic Protective Effect of Curcumin and Resveratrol against Oxidative Stress in Endothelial EAhy926 Cells. Evid.-Based Complement. Altern. Med. 2021, 2021, 2661025. [Google Scholar] [CrossRef]
- Puengel, T.; Liu, H.; Guillot, A.; Heymann, F.; Tacke, F.; Peiseler, M. Nuclear Receptors Linking Metabolism, Inflammation, and Fibrosis in Nonalcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2022, 23, 2668. [Google Scholar] [CrossRef] [PubMed]
- Treviño, L.S.; Katz, T.A. Endocrine Disruptors and Developmental Origins of Nonalcoholic Fatty Liver Disease. Endocrinology 2018, 159, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Dallio, M.; Diano, N.; Masarone, M.; Gravina, A.G.; Patanè, V.; Romeo, M.; Di Sarno, R.; Errico, S.; Nicolucci, C.; Abenavoli, L.; et al. Chemical Effect of Bisphenol a on Non-Alcoholic Fatty Liver Disease. Int. J. Environ. Res. Public Health 2019, 16, 3134. [Google Scholar] [CrossRef]
- Grasselli, E.; Cortese, K.; Voci, A.; Vergani, L.; Fabbri, R.; Barmo, C.; Gallo, G.; Canesi, L. Direct Effects of Bisphenol A on Lipid Homeostasis in Rat Hepatoma Cells. Chemosphere 2013, 91, 1123–1129. [Google Scholar] [CrossRef]
- Gerona, R.R.; Woodruff, T.J.; Dickenson, C.A.; Pan, J.; Schwartz, J.M.; Sen, S.; Friesen, M.W.; Fujimoto, V.Y.; Hunt, P.A. Bisphenol-A (BPA), BPA Glucuronide, and BPA Sulfate in Midgestation Umbilical Cord Serum in a Northern and Central California Population. Environ. Sci. Technol. 2013, 47, 12477–12485. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, H.; Lin, S.; Chen, T.; Chang, D.; Sun, Y.; Wang, C.; Liu, Y.; Lu, Y.; Song, J.; et al. Advanced Effect of Curcumin and Resveratrol on Mitigating Hepatic Steatosis in Metabolic Associated Fatty Liver Disease via the PI3K/AKT/MTOR and HIF-1/VEGF Cascade. Biomed. Pharmacother. 2023, 165, 115279. [Google Scholar] [CrossRef]
- Messerlian, C.; Martinez, R.M.; Hauser, R.; Baccarelli, A.A. “Omics” and Endocrine-Disrupting Chemicals-New Paths Forward. Nat. Rev. Endocrinol. 2017, 13, 740–748. [Google Scholar] [CrossRef]
- Muftuoglu, M.; Mori, M.P.; Souza-Pinto, N.C. de Formation and Repair of Oxidative Damage in the Mitochondrial DNA. Mitochondrion 2014, 17, 164–181. [Google Scholar] [CrossRef]
- Kim, J.T.; Lee, H.K. Metabolic Syndrome and the Environmental Pollutants from Mitochondrial Perspectives. Rev. Endocr. Metab. Disord. 2014, 15, 253–262. [Google Scholar] [CrossRef]
- Khan, S.; Beigh, S.; Chaudhari, B.P.; Sharma, S.; Aliul Hasan Abdi, S.; Ahmad, S.; Ahmad, F.; Parvez, S.; Raisuddin, S. Mitochondrial Dysfunction Induced by Bisphenol A Is a Factor of Its Hepatotoxicity in Rats. Environ. Toxicol. 2016, 31, 1922–1934. [Google Scholar] [CrossRef]
- Pu, Y.; Pearl, S.; Gingrich, J.; Jing, J.; Martin, D.; Murga-Zamalloa, C.A.; Veiga-Lopez, A. Multispecies Study: Low-Dose Tributyltin Impairs Ovarian Theca Cell Cholesterol Homeostasis through the RXR Pathway in Five Mammalian Species Including Humans. Arch. Toxicol. 2019, 93, 1665–1677. [Google Scholar] [CrossRef] [PubMed]
- Buñay, J.; Larriba, E.; Patiño-Garcia, D.; Cruz-Fernandes, L.; Castañeda-Zegarra, S.; Rodriguez-Fernandez, M.; del Mazo, J.; Moreno, R.D. Differential Effects of Exposure to Single versus a Mixture of Endocrine-Disrupting Chemicals on Steroidogenesis Pathway in Mouse Testes. Toxicol. Sci. 2018, 161, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Pan, J.; Zhou, W.; Ji, G.; Dang, Y. Recent Advances in Lean NAFLD. Biomed. Pharmacother. 2022, 153, 113331. [Google Scholar] [CrossRef]
- Sun, Q.F.; Tang, L.J.; Wang, M.J.; Zhu, P.W.; Li, Y.Y.; Ma, H.L.; Huang, O.Y.; Hong, L.; Li, G.; Byrne, C.D.; et al. Potential Blood DNA Methylation Biomarker Genes for Diagnosis of Liver Fibrosis in Patients with Biopsy-Proven Non-Alcoholic Fatty Liver Disease. Front. Med. 2022, 9, 864570. [Google Scholar] [CrossRef] [PubMed]
- Cano, R.; Pérez, J.L.; Dávila, L.A.; Ortega, Á.; Gómez, Y.; Valero-Cedeño, N.J.; Parra, H.; Manzano, A.; Castro, T.I.V.; Díaz Albornoz, M.P.; et al. Role of Endocrine-Disrupting Chemicals in the Pathogenesis of Non-Alcoholic Fatty Liver Disease: A Comprehensive Review. Int. J. Mol. Sci. 2021, 22, 4807. [Google Scholar] [CrossRef] [PubMed]
- Qin, T.; Zhang, X.; Guo, T.; Yang, T.; Gao, Y.; Hao, W.; Xiao, X.F. Epigenetic Alteration Shaped by the Environmental Chemical Bisphenol A. Front. Genet. 2021, 11, 618966. [Google Scholar] [CrossRef] [PubMed]
- Kundakovic, M.; Gudsnuk, K.; Franks, B.; Madrid, J.; Miller, R.L.; Perera, F.P.; Champagne, F.A. Sex-Specific Epigenetic Disruption and Behavioral Changes Following Low-Dose in Utero Bisphenol a Exposure. Proc. Natl. Acad. Sci. USA 2013, 110, 9956–9961. [Google Scholar] [CrossRef] [PubMed]
- Ke, Z.H.; Pan, J.X.; Jin, L.Y.; Xu, H.Y.; Yu, T.T.; Ullah, K.; Rahman, T.U.; Ren, J.; Cheng, Y.; Dong, X.Y.; et al. Bisphenol A Exposure May Induce Hepatic Lipid Accumulation via Reprogramming the DNA Methylation Patterns of Genes Involved in Lipid Metabolism. Sci. Rep. 2016, 6, 31331. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; Gallego-Durán, R.; Gallego, P.; Grande, L. Genetic and Epigenetic Regulation in Nonalcoholic Fatty Liver Disease (NAFLD). Int. J. Mol. Sci. 2018, 19, 991. [Google Scholar] [CrossRef]
- Montjean, D.; Neyroud, A.S.; Yefimova, M.G.; Benkhalifa, M.; Cabry, R.; Ravel, C. Impact of Endocrine Disruptors upon Non-Genetic Inheritance. Int. J. Mol. Sci. 2022, 23, 3350. [Google Scholar] [CrossRef]
- Nakanishi, T.; Kohroki, J.; Suzuki, S.; Ishizaki, J.; Hiromori, Y.; Takasuga, S.; Watanabe, Y.; Utoguchi, N.; Tanaka, K. Trialkyltin Compounds Enhance Human CG Secretion and Aromatase Activity in Human Placental Choriocarcinoma Cells. J. Clin. Endocrinol. Metab. 2002, 87, 2830–2837. [Google Scholar] [CrossRef] [PubMed]
- Azumi, K.; Fujie, M.; Usami, T.; Miki, Y.; Satoh, N. A CDNA Microarray Technique Applied for Analysis of Global Gene Expression Profiles in Tributyltin-Exposed Ascidians. Mar. Environ. Res. 2004, 58, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Osada, S.; Nishikawa, J.I.; Nakanishi, T.; Tanaka, K.; Nishihara, T. Some Organotin Compounds Enhance Histone Acetyltransferase Activity. Toxicol. Lett. 2005, 155, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, M.; Zobeiry, M.; Abdolahi, S.; Hatami, B.; Zali, M.R.; Baghaei, K. A Crosstalk between Epigenetic Modulations and Non-Alcoholic Fatty Liver Disease Progression. Pathol. Res. Pract. 2023, 251, 154809. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Chen, Y.; Ning, Z.; Li, Q.; Han, B.; Zhu, C.; Chen, Y.; Xia, F.; Jiang, B.; Wang, B.; et al. Exposure to Famine in Early Life and Nonalcoholic Fatty Liver Disease in Adulthood. J. Clin. Endocrinol. Metab. 2016, 101, 2218–2225. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, S.R.; Kasmi, K.C.E.; Jonscher, K.R.; Friedman, J.E. Developmental Origins of NAFLD: A Womb with a Clue. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Liu, Y.; Xue, X.; Yuan, C.; Wang, Z. BPA’s Transgenerational Disturbance to Transcription of Ovarian Steroidogenic Genes in Rare Minnow Gobiocypris Rarus via DNA and Histone Methylation. Sci. Total Environ. 2021, 762, 143055. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, E.E.; Ben Maamar, M.; Skinner, M.K. Role of Epigenetic Transgenerational Inheritance in Generational Toxicology. Environ. Epigenetics 2022, 8, dvac001. [Google Scholar] [CrossRef] [PubMed]
- Chamorro-García, R.; Sahu, M.; Abbey, R.J.; Laude, J.; Pham, N.; Blumberg, B. Transgenerational Inheritance of Increased Fat Depot Size, Stem Cell Reprogramming, and Hepatic Steatosis Elicited by Prenatal Exposure to the Obesogen Tributyltin in Mice. Environ. Health Perspect. 2013, 121, 359–366. [Google Scholar] [CrossRef]
- Ma, Y.; Xia, W.; Wang, D.Q.; Wan, Y.J.; Xu, B.; Chen, X.; Li, Y.Y.; Xu, S.Q. Hepatic DNA Methylation Modifications in Early Development of Rats Resulting from Perinatal BPA Exposure Contribute to Insulin Resistance in Adulthood. Diabetologia 2013, 56, 2059–2067. [Google Scholar] [CrossRef]
- Gjorgjieva, M.; Sobolewski, C.; Dolicka, D.; Correia De Sousa, M.; Foti, M. MiRNAs and NAFLD: From Pathophysiology to Therapy. Gut 2019, 68, 2065–2079. [Google Scholar] [CrossRef]
- Liu, C.H.; Ampuero, J.; Gil-Gómez, A.; Montero-Vallejo, R.; Rojas, Á.; Muñoz-Hernández, R.; Gallego-Durán, R.; Romero-Gómez, M. MiRNAs in Patients with Non-Alcoholic Fatty Liver Disease: A Systematic Review and Meta-Analysis. J. Hepatol. 2018, 69, 1335–1348. [Google Scholar] [CrossRef]
- De Felice, B.; Manfellotto, F.; Palumbo, A.; Troisi, J.; Zullo, F.; Di Carlo, C.; Sardo, A.D.S.; De Stefano, N.; Ferbo, U.; Guida, M.; et al. Genome-Wide MicroRNA Expression Profiling in Placentas from Pregnant Women Exposed to BPA. BMC Med. Genom. 2015, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Craig, P.M.; Trudeau, V.L.; Moon, T.W. Profiling Hepatic MicroRNAs in Zebrafish: Fluoxetine Exposure Mimics a Fasting Response That Targets AMP-Activated Protein Kinase (AMPK). PLoS ONE 2014, 9, e95351. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.Q.; Young, M.R.; Diwan, B.A.; Coogan, T.P.; Waalkes, M.P. Association of Arsenic-Induced Malignant Transformation with DNA Hypomethylation and Aberrant Gene Expression. Proc. Natl. Acad. Sci. USA 1997, 94, 10907–10912. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, S.F.; Liu, J.; Diwan, B.A.; Barrett, J.C.; Waalkes, M.P. Chronic Inorganic Arsenic Exposure Induces Hepatic Global and Individual Gene Hypomethylation: Implications for Arsenic Hepatocarcinogenesis. Carcinogenesis 2004, 25, 1779–1786. [Google Scholar] [CrossRef] [PubMed]
- Weinhouse, C.; Bergin, I.L.; Harris, C.; Dolinoy, D.C. Stat3 Is a Candidate Epigenetic Biomarker of Perinatal Bisphenol A Exposure Associated with Murine Hepatic Tumors with Implications for Human Health. Epigenetics 2015, 10, 1099–1110. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Ling, Y.; Jiang, J.; Wang, D.; Wang, J.; Li, J.; Wang, X.; Wang, H. Differential Mechanisms Regarding Triclosan vs. Bisphenol A and Fluorene-9-Bisphenol Induced Zebrafish Lipid-Metabolism Disorders by RNA-Seq. Chemosphere 2020, 251, 126318. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, L.S.; Oliveira, K.M.; Freitas, I.N.; Silva, J.A.; Silva, J.N.; Favero-Santos, B.C.; Bonfleur, M.L.; Carneiro, E.M.; Ribeiro, R.A. Bisphenol-A Exposure Worsens Hepatic Steatosis in Ovariectomized Mice Fed on a High-Fat Diet: Role of Endoplasmic Reticulum Stress and Fibrogenic Pathways. Life Sci. 2020, 256, 118012. [Google Scholar] [CrossRef]
- Massart, J.; Begriche, K.; Corlu, A.; Fromenty, B. Xenobiotic-Induced Aggravation of Metabolic-Associated Fatty Liver Disease. Int. J. Mol. Sci. 2022, 23, 1062. [Google Scholar] [CrossRef]
- Lin, Y.; Ding, D.; Huang, Q.; Liu, Q.; Lu, H.; Lu, Y.; Chi, Y.; Sun, X.; Ye, G.; Zhu, H.; et al. Downregulation of MiR-192 Causes Hepatic Steatosis and Lipid Accumulation by Inducing SREBF1: Novel Mechanism for Bisphenol A-Triggered Non-Alcoholic Fatty Liver Disease. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Li, Q.; Zhang, H.; Zou, J.; Feng, X. Bisphenol A Induces Cholesterol Biosynthesis in HepG2 Cells via SREBP-2/HMGCR Signaling Pathway. J. Toxicol. Sci. 2019, 44, 481–491. [Google Scholar]
- Albhaisi, S.; Sanyal, A.J. Gene-Environmental Interactions as Metabolic Drivers of Nonalcoholic Steatohepatitis. Front. Endocrinol. 2021, 12, 665987. [Google Scholar] [CrossRef] [PubMed]
- Kalafati, I.P.; Dimitriou, M.; Revenas, K.; Kokkinos, A.; Deloukas, P.; Dedoussis, G.V. TM6SF2-Rs58542926 Genetic Variant Modifies the Protective Effect of a “Prudent” Dietary Pattern on Serum Triglyceride Levels. Nutrients 2023, 15, 1112. [Google Scholar] [CrossRef] [PubMed]
- Kalafati, I.P.; Dimitriou, M.; Borsa, D.; Vlachogiannakos, J.; Revenas, K.; Kokkinos, A.; Ladas, S.D.; Dedoussis, G.V. Fish Intake Interacts with TM6SF2 Gene Variant to Affect NAFLD Risk: Results of a Case–Control Study. Eur. J. Nutr. 2019, 58, 1463–1473. [Google Scholar] [CrossRef]
- Wallis, D.J.; Truong, L.; La Du, J.; Tanguay, R.L.; Reif, D.M. Uncovering Evidence for Endocrine-Disrupting Chemicals That Elicit Differential Susceptibility through Gene-Environment Interactions. Toxics 2021, 9, 77. [Google Scholar] [CrossRef]
- Dunaway, K.W.; Islam, M.S.; Coulson, R.L.; Lopez, S.J.; Vogel Ciernia, A.; Chu, R.G.; Yasui, D.H.; Pessah, I.N.; Lott, P.; Mordaunt, C.; et al. Cumulative Impact of Polychlorinated Biphenyl and Large Chromosomal Duplications on DNA Methylation, Chromatin, and Expression of Autism Candidate Genes. Cell Rep. 2016, 17, 3035–3048. [Google Scholar] [CrossRef]
- Zhong, R.; He, H.; Jin, M.; Lu, Z.; Deng, Y.; Liu, C.; Shen, N.; Li, J.; Wang, H.; Ying, P.; et al. Genome-Wide Gene-Bisphenol A, F and Triclosan Interaction Analyses on Urinary Oxidative Stress Markers. Sci. Total Environ. 2022, 807, 150753. [Google Scholar] [CrossRef]
- Brokken, L.J.S.; Giwercman, Y.L. Gene-Environment Interactions in Male Reproductive Health: Special Reference to the Aryl Hydrocarbon Receptor Signaling Pathway. Asian J. Androl. 2014, 16, 89–96. [Google Scholar] [CrossRef]
- Jin, W.; Otake, M.; Eguchi, A.; Sakurai, K.; Nakaoka, H.; Watanabe, M.; Todaka, E.; Mori, C. Dietary Habits and Cooking Methods Could Reduce Avoidable Exposure to PCBs in Maternal and Cord Sera. Sci. Rep. 2017, 7, 17357. [Google Scholar] [CrossRef]
- ISS. Knowing, Reducing, Preventing Endocrine Disruptors: A Decalogue for Citizens; Italian Ministry for the Environment, Land and Sea National Istitute of Health: Rome, Italy, 2014. [Google Scholar]
- Martin, L.; Zhang, Y.; First, O.; Mustieles, V.; Dodson, R.; Rosa, G.; Coburn-Sanderson, A.; Adams, C.D.; Messerlian, C. Lifestyle Interventions to Reduce Endocrine-Disrupting Phthalate and Phenol Exposures among Reproductive Age Men and Women: A Review and Future Steps. Environ. Int. 2022, 170, 107576. [Google Scholar] [CrossRef] [PubMed]
- Feroe, A.; Broene, R.; Albuquerque, D.; Ruiz, P. Endocrine Disrupting Chemicals, Transgenerational Epigenetics and Metabolic Diseases. EC Endocrinol. Metab. Res. 2017, 21, 31. [Google Scholar] [PubMed]
- Torres, M.C.P.; Aghemo, A.; Lleo, A.; Bodini, G.; Furnari, M.; Marabotto, E.; Miele, L.; Giannini, E.G. Mediterranean Diet and NAFLD: What We Know and Questions That Still Need to Be Answered. Nutrients 2019, 11, 2971. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Dietary recommendations |
| Prefer fresh fruits, vegetables, fish and preferably seasonal products. |
| Buy tomato sauce stored in glass jars and not plastic bottles. |
| Avoid using popcorn bags in the microwave, instead cook the popcorn in the pot. |
| Choose products that are not stored in plastic containers or cans, as ready-to-eat and fast-food products. |
| Prefer sandwiches or bakery products unwrapped, just freshly exposed at the countertop. |
| Avoid burned, smoked or pre-fried foods and, when possible, remove the burned parts (such as meat, meat by-products and canned fish). |
| Avoid frozen fish, pizza, and other foods. |
| Avoid packed meals such as instant soups and noodles. |
| Avoid the use of plastic coffee or tea bags, when not certified as EDC free and use loose tea/coffee. |
| Cooking, kitchen articles & food storage |
| Avoid using non-stick cooking utensils if the coating is worn. |
| Avoid grilling, barbecuing, deep frying and overheating of food. |
| Consult the manufacturer’s instructions when packaging food and use grease-proof paper or film. |
| Use plastic food containers only in perfect condition. |
| Always follow the manufacturer’s instructions and use only plastic bottles or cans in perfect condition, when heating food and/or milk for babies. |
| When using plastic containers, not labelled as appropriate for high temperatures, only pour cold food or beverages. |
| Avoid using polycarbonate baby bottles for milk/water. |
| Don’t put plastic containers to be heated in the microwave/oven. |
| Products, proper ventilation and house practices |
| Avoid buying materials with soft PVC that contain di (2-ethylhexyl) phthalate or DEHP. |
| Avoid the use of soft PVC floors for children. Instead use a carpet made of untreated fibres (i.e., wool, cotton). |
| Avoid PVC objects/toys for children. |
| Substitute worn out or damaged wrappings of objects having foam padding such as car seats and mattresses. |
| Avoid candle smoke or cigarette smoke in your living environment. |
| Make sure of proper house ventilation to avoid dust accumulation indoors and properly maintain your vacuum cleaner (change filters, empty the dust bag, etc.). |
| Buy clothing products with known origins and composition instead of waterproof or anti-stain labels. |
| Check product labels and avoid beauty products with parabens and/or phthalates. |
| Use water-based sunscreen to avoid the use of chemicals. |
| Avoid fragrances, body sprays, perfumes and the unnecessary use of cosmetics, cleaning, dental care products. |
| Avoid using scented or antibacterial hand soaps, candles, or air fresheners. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mentsiou Nikolaou, E.; Kalafati, I.P.; Dedoussis, G.V. The Interplay between Endocrine-Disrupting Chemicals and the Epigenome towards Metabolic Dysfunction-Associated Steatotic Liver Disease: A Comprehensive Review. Nutrients 2024, 16, 1124. https://doi.org/10.3390/nu16081124
Mentsiou Nikolaou E, Kalafati IP, Dedoussis GV. The Interplay between Endocrine-Disrupting Chemicals and the Epigenome towards Metabolic Dysfunction-Associated Steatotic Liver Disease: A Comprehensive Review. Nutrients. 2024; 16(8):1124. https://doi.org/10.3390/nu16081124
Chicago/Turabian StyleMentsiou Nikolaou, Evangelia, Ioanna Panagiota Kalafati, and George V. Dedoussis. 2024. "The Interplay between Endocrine-Disrupting Chemicals and the Epigenome towards Metabolic Dysfunction-Associated Steatotic Liver Disease: A Comprehensive Review" Nutrients 16, no. 8: 1124. https://doi.org/10.3390/nu16081124
APA StyleMentsiou Nikolaou, E., Kalafati, I. P., & Dedoussis, G. V. (2024). The Interplay between Endocrine-Disrupting Chemicals and the Epigenome towards Metabolic Dysfunction-Associated Steatotic Liver Disease: A Comprehensive Review. Nutrients, 16(8), 1124. https://doi.org/10.3390/nu16081124