

Antitumor Effect and Gut Microbiota Modulation by Quercetin, Luteolin, and Xanthohumol in a Rat Model for Colorectal Cancer Prevention

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Colorectal Cancer Induction and Monitoring
2.3. Tissue Samples
2.4. Genomic DNA Extraction and 16S Ribosomal RNA Sequencing for Gut Microbiota Analysis
2.5. Bioinformatics Analysis
2.6. Statistical Methods
3. Results
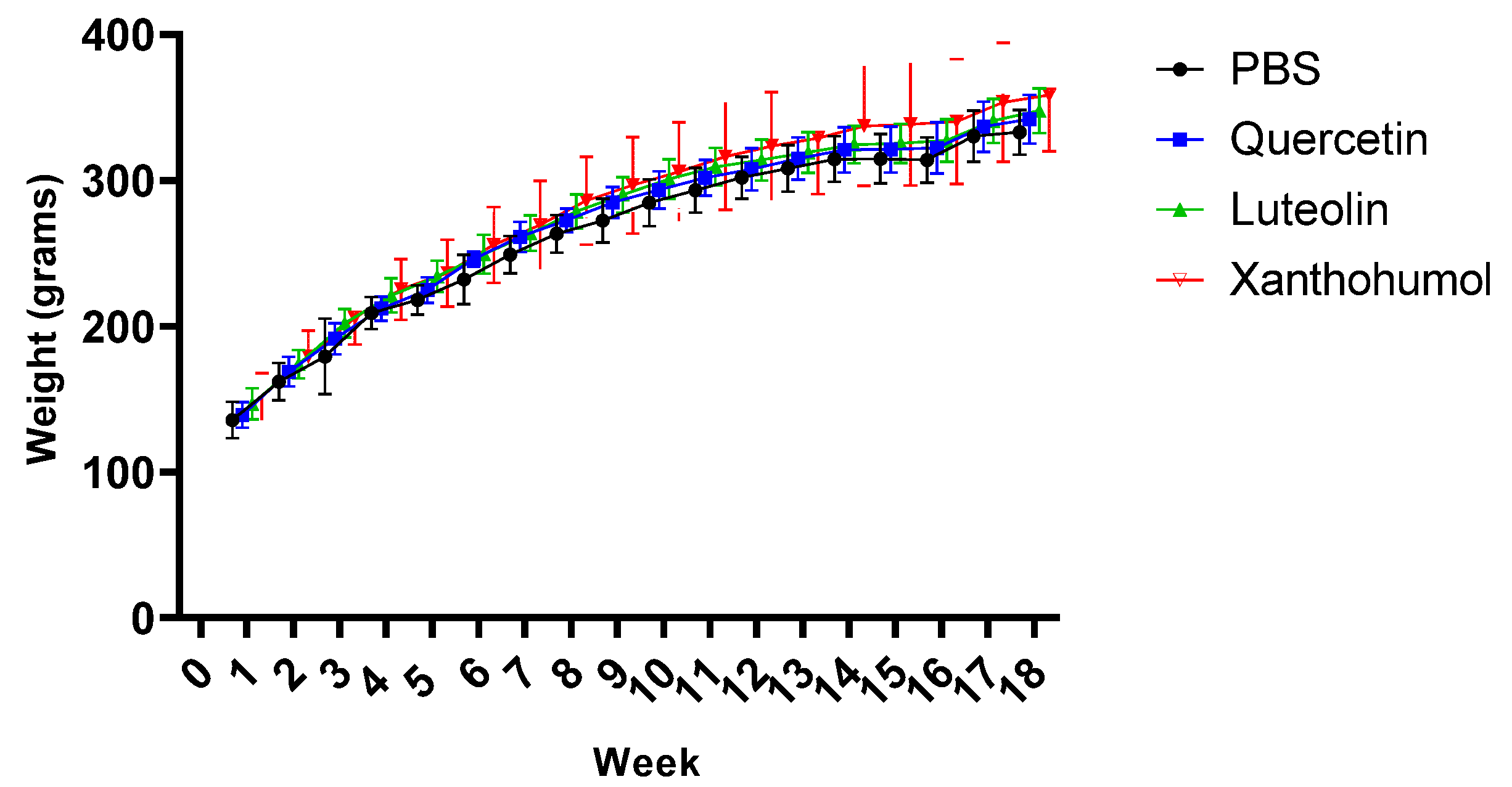
3.1. Effect of Quercetin, Luteolin, and Xanthohumol Administration on Body Weight
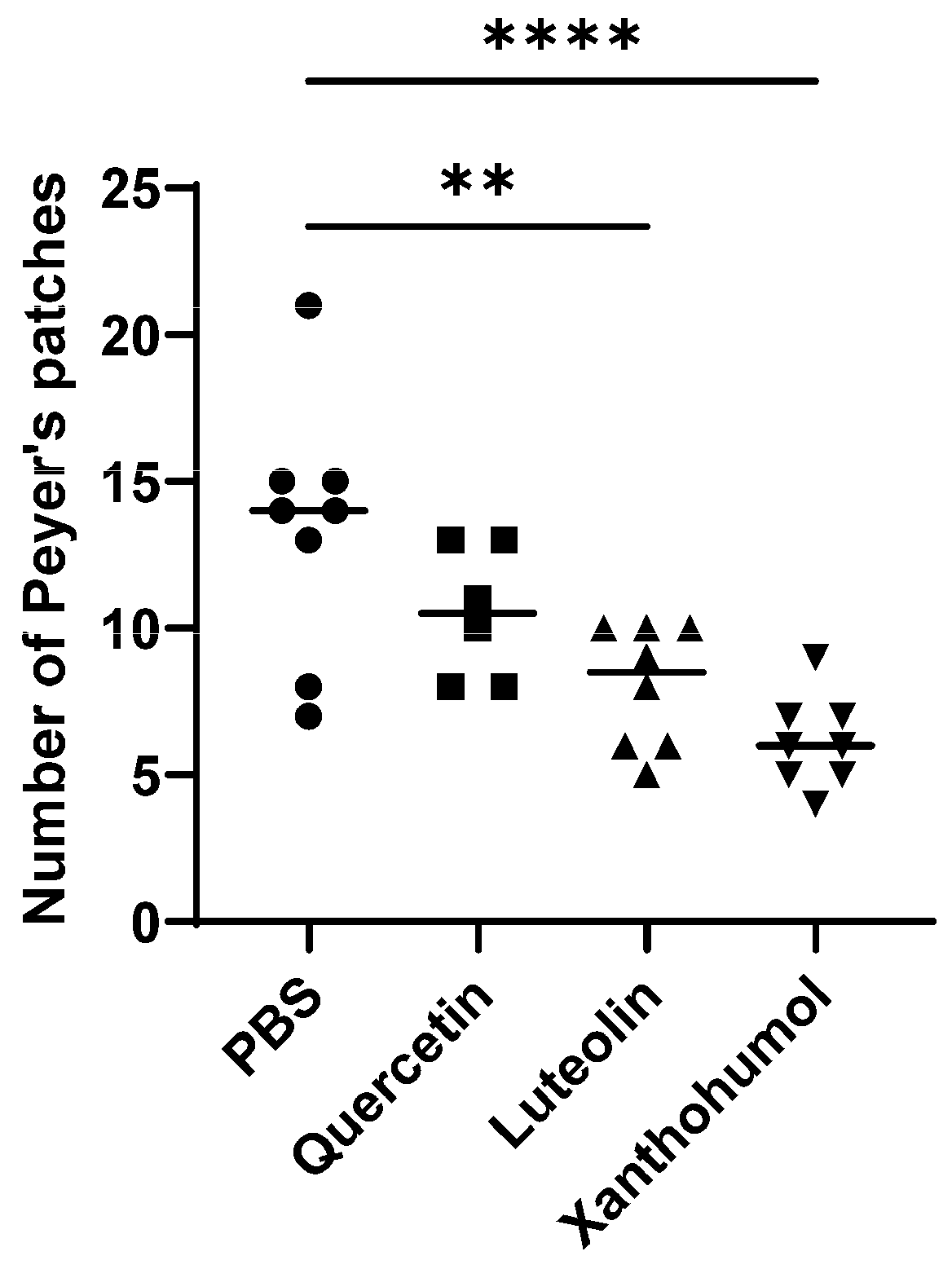
3.2. Effect of Quercetin, Luteolin, and Xanthohumol Administration on Hyperplastic Peyer’s Patches
3.3. Effect of Quercetin, Luteolin, and Xanthohumol Administration on Caecum Weight
3.4. Effect of Quercetin, Luteolin, and Xanthohumol Administration on Colon Length
3.5. Effect of Quercetin, Luteolin, and Xanthohumol Administration on the Number of Tumors
3.6. Effect of Quercetin, Luteolin, and Xanthohumol Administration on the Gut Microbiota
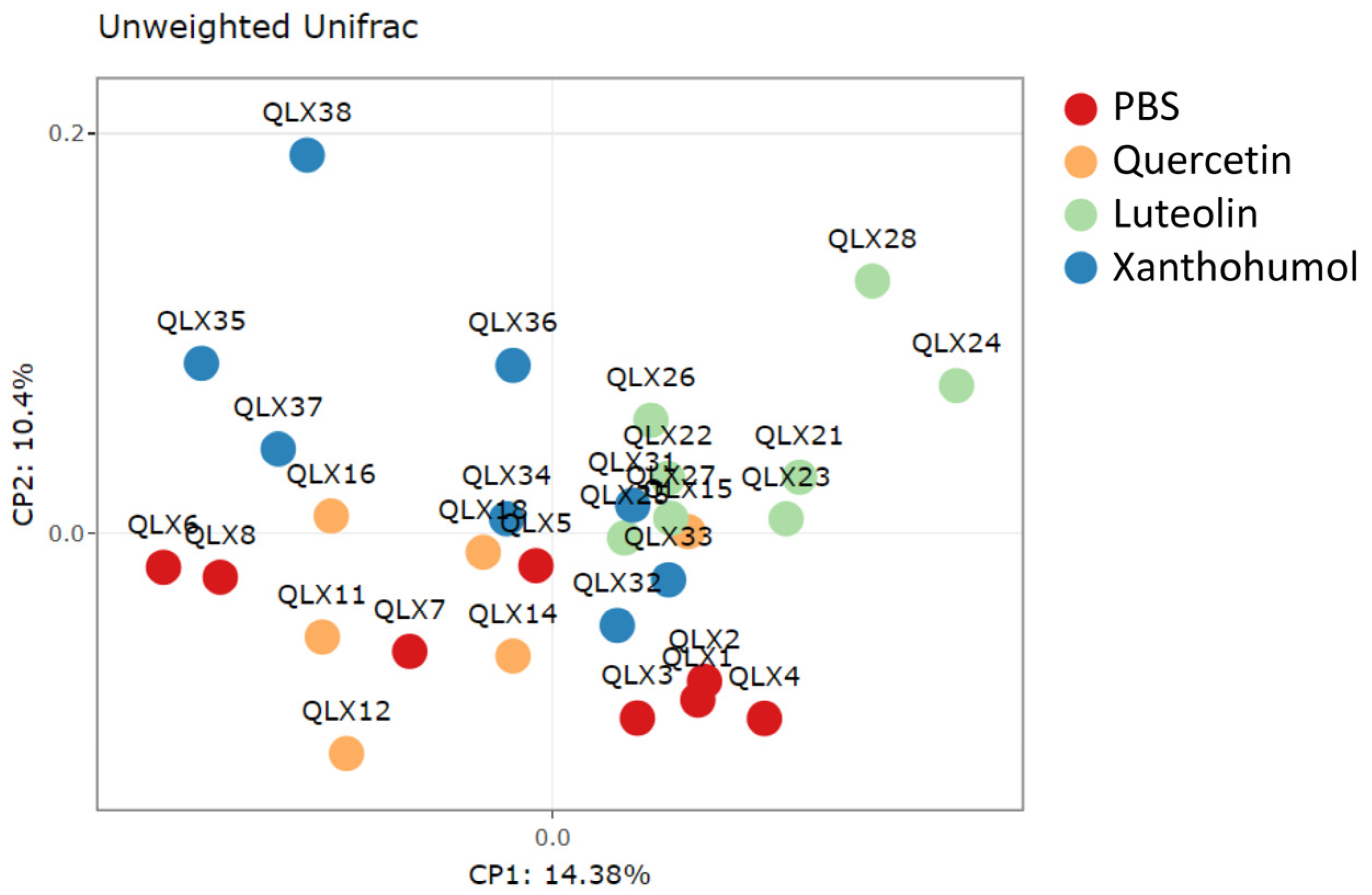
3.6.1. Alpha and Beta Diversity
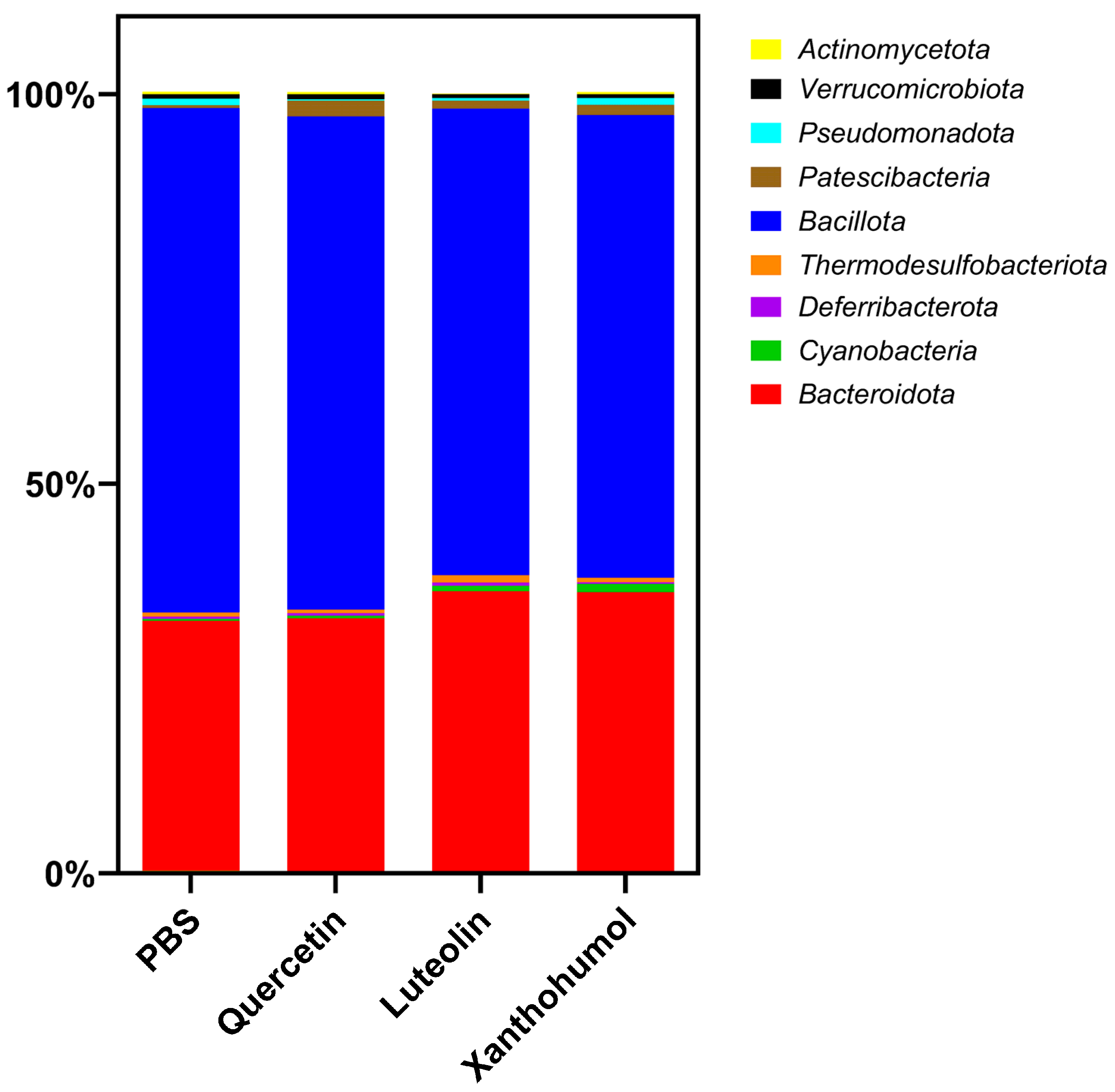
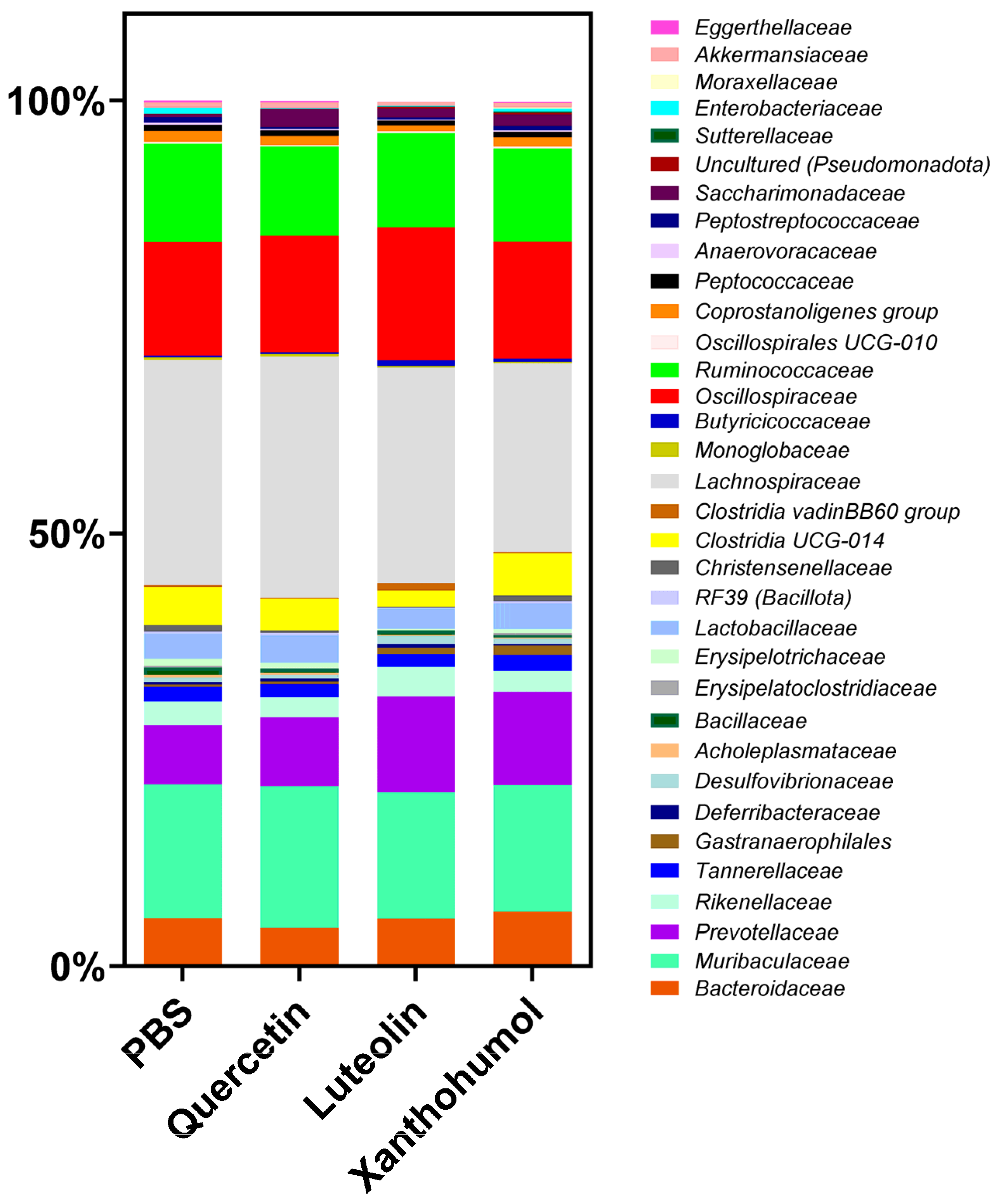
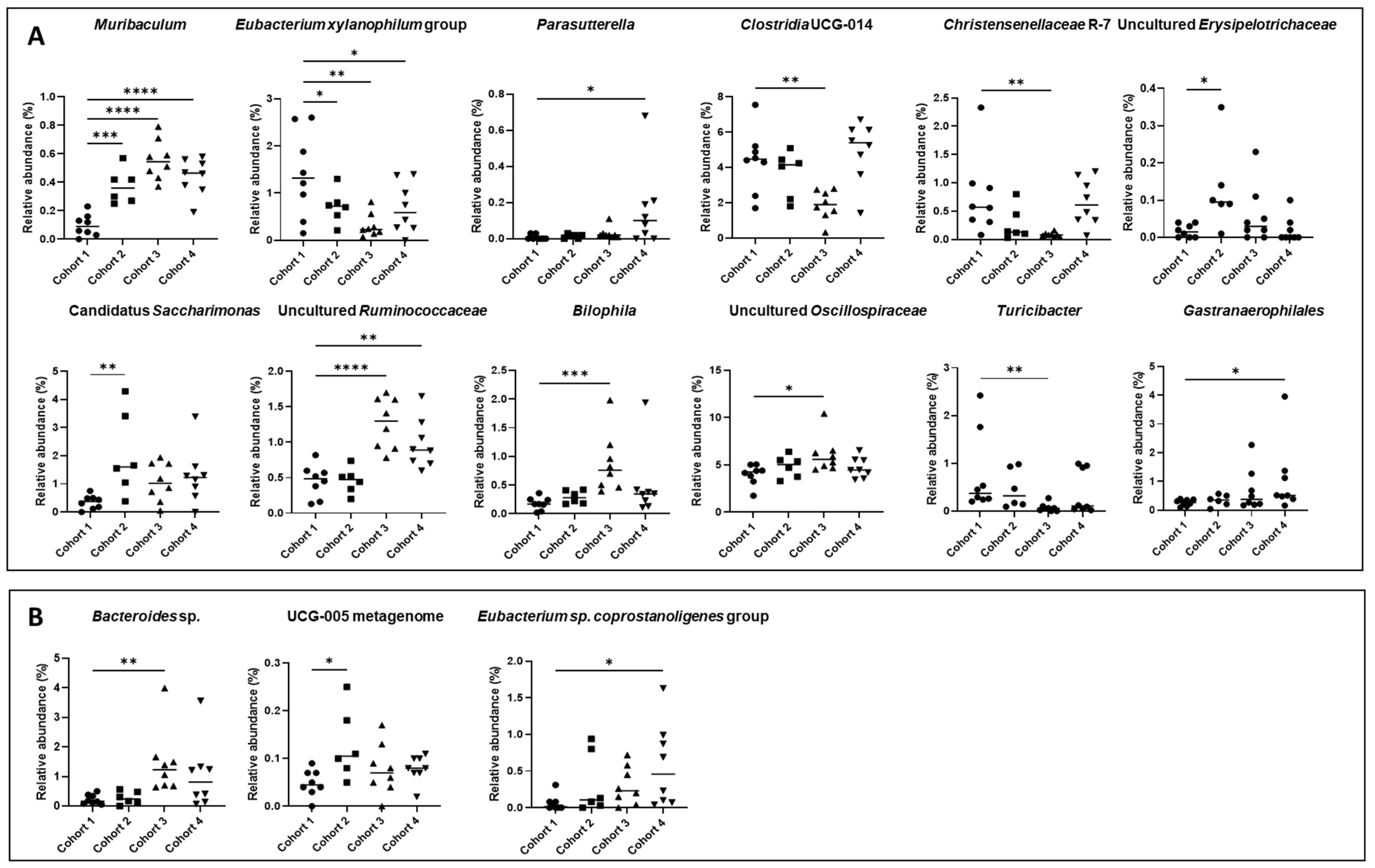
3.6.2. Taxonomic Profile
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Imran, M.; Rauf, A.; Abu-Izneid, T.; Nadeem, M.; Shariati, M.A.; Khan, I.A.; Imran, A.; Orhan, I.E.; Rizwan, M.; Atif, M.; et al. Luteolin, a Flavonoid, as an Anticancer Agent: A Review. Biomed. Pharmacother. 2019, 112, 108612. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.P.; Rajavel, T.; Nabavi, S.F.; Setzer, W.N.; Ahmadi, A.; Mansouri, K.; Nabavi, S.M. Hesperidin: A Promising Anticancer Agent from Nature. Ind. Crops Prod. 2015, 76, 582–589. [Google Scholar] [CrossRef]
- Xi, Y.; Xu, P. Global Colorectal Cancer Burden in 2020 and Projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Constantinou, V.; Constantinou, C. Focusing on Colorectal Cancer in Young Adults (Review). Mol. Clin. Oncol. 2024, 20, 8. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; van de Velde, C.J.H.; Watanabe, T. Colorectal Cancer. Nat. Rev. Dis. Primers 2015, 1, 15065. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global Cancer Statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; García, L.; Monte, J.; Villar, C.J.; Lombó, F. Functional Anthocyanin-Rich Sausages Diminish Colorectal Cancer in an Animal Model and Reduce pro-Inflammatory Bacteria in the Intestinal Microbiota. Genes 2018, 9, 133. [Google Scholar] [CrossRef] [PubMed]
- Parmar, S.; Easwaran, H. Genetic and Epigenetic Dependencies in Colorectal Cancer Development. Gastroenterol. Rep. 2022, 10, goac035. [Google Scholar] [CrossRef]
- Kim, U.; Lee, D.S. Epigenetic Regulations in Mammalian Cells: Roles and Profiling Techniques. Mol. Cells 2023, 46, 86–98. [Google Scholar] [CrossRef]
- Zeki, S.S.; Graham, T.A.; Wright, N.A. Stem Cells and Their Implications for Colorectal Cancer. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Colussi, D.; Brandi, G.; Bazzoli, F.; Ricciardiello, L. Molecular Pathways Involved in Colorectal Cancer: Implications for Disease Behavior and Prevention. Int. J. Mol. Sci. 2013, 14, 16365–16385. [Google Scholar] [CrossRef] [PubMed]
- Grady, W.M.; Carethers, J.M. Genomic and Epigenetic Instability in Colorectal Cancer Pathogenesis. Gastroenterology 2008, 135, 1079–1099. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Chen, W.D.; Parmigiani, G.; Diehl, F.; Beerenwinkel, N.; Antal, T.; Traulsen, A.; Nowak, M.A.; Siegel, C.; Velculescu, V.E.; et al. Comparative Lesion Sequencing Provides Insights into Tumor Evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 4283–4288. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, S.; Sengupta, B.; Taylor, J.; Pahari, B.P.; Sengupta, P.K. Interactions of Dietary Flavonoids with Proteins: Insights from Fluorescence Spectroscopy and Other Related Biophysical Studies. Curr. Drug Metab. 2013, 14, 491–503. [Google Scholar] [CrossRef]
- González-Vallinas, M.; González-Castejón, M.; Rodríguez-Casado, A.; Ramírez de Molina, A. Dietary Phytochemicals in Cancer Prevention and Therapy: A Complementary Approach with Promising Perspectives. Nutr. Rev. 2013, 71, 585–599. [Google Scholar] [CrossRef]
- Li, A.-N.; Li, S.; Zhang, Y.-J.; Xu, X.-R.; Chen, Y.-M.; Li, H.-B. Resources and Biological Activities of Natural Polyphenols. Nutrients 2014, 6, 6020–6047. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef]
- Rodríguez-García, C.; Sánchez-Quesada, C.; Gaforio, J.J. Dietary Flavonoids as Cancer Chemopreventive Agents: An Updated Review of Human Studies. Antioxidants 2019, 8, 137. [Google Scholar] [CrossRef] [PubMed]
- Abotaleb, M.; Samuel, S.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers 2018, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Chirumbolo, S.; Bjørklund, G.; Lysiuk, R.; Vella, A.; Lenchyk, L.; Upyr, T. Targeting Cancer with Phytochemicals via Their Fine Tuning of the Cell Survival Signaling Pathways. Int. J. Mol. Sci. 2018, 19, 3568. [Google Scholar] [CrossRef] [PubMed]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as Anticancer Agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed]
- Gorlach, S.; Fichna, J.; Lewandowska, U. Polyphenols as Mitochondria-Targeted Anticancer Drugs. Cancer Lett. 2015, 366, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Perez-Vizcaino, F.; Fraga, C.G. Research Trends in Flavonoids and Health. Arch. Biochem. Biophys. 2018, 646, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Imran, M.; Khan, I.A.; ur-Rehman, M.; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer Potential of Quercetin: A Comprehensive Review. Phytother. Res. 2018, 32, 2109–2130. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.H.; Sun, T.L.; Xiang, D.X.; Wei, S.S.; Li, W.Q. Anticancer Activity and Mechanism of Xanthohumol: A Prenylated Flavonoid from Hops (Humulus lupulus L.). Front. Pharmacol. 2018, 9, 530. [Google Scholar] [CrossRef]
- Harwood, M.; Danielewska-Nikiel, B.; Borzelleca, J.F.; Flamm, G.W.; Williams, G.M.; Lines, T.C. A Critical Review of the Data Related to the Safety of Quercetin and Lack of Evidence of in Vivo Toxicity, Including Lack of Genotoxic/Carcinogenic Properties. Food Chem. Toxicol. 2007, 45, 2179–2205. [Google Scholar] [CrossRef]
- Kawanishi, S.; Oikawa, S.; Murata, M. Evaluation for Safety of Antioxidant Chemopreventive Agents. Antioxid. Redox Signal. 2005, 7, 1728–1739. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, L.; Li, G.; Gao, Z. Xanthohumol Protects against Azoxymethane-Induced Colorectal Cancer in Sprague-Dawley Rats. Environ. Toxicol. 2020, 35, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Vanhoecke, B.W.; Delporte, F.; Van Braeckel, E.; Heyerick, A.; Depypere, H.T.; Nuytinck, M.; De Keukeleire, D.; Bracke, M.E. A Safety Study of Oral Tangeretin and Xanthohumol Administration to Laboratory Mice. In Vivo 2005, 19, 103–107. [Google Scholar] [PubMed]
- Miranda, C.L.; Stevens, J.F.; Helmrich, A.; Henderson, M.C.; Rodriguez, R.J.; Yang, Y.-H.; Deinzer, M.L.; Barnes, D.W.; Buhler, D.R. Antiproliferative and Cytotoxic Effects of Prenylated Flavonoids from Hops (Humulus lupulus) in Human Cancer Cell Lines. Food Chem. Toxicol. 1999, 37, 271–285. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, H.J.; Lee, J.S.; Lee, I.-S.; Kang, B.Y. Inhibition of Topoisomerase I Activity and Efflux Drug Transporters’ Expression by Xanthohumol from Hops. Arch. Pharm. Res. 2007, 30, 1435–1439. [Google Scholar] [CrossRef]
- Fernández, J.; Silván, B.; Entrialgo-Cadierno, R.; Villar, C.J.; Capasso, R.; Uranga, J.A.; Lombó, F.; Abalo, R. Antiproliferative and Palliative Activity of Flavonoids in Colorectal Cancer. Biomed. Pharmacother. 2021, 143, 112241. [Google Scholar] [CrossRef]
- Nejabati, H.R.; Roshangar, L. Kaempferol: A Potential Agent in the Prevention of Colorectal Cancer. Physiol. Rep. 2022, 10, e15488. [Google Scholar] [CrossRef]
- Al Shoyaib, A.; Archie, S.R.; Karamyan, V.T. Intraperitoneal Route of Drug Administration: Should It Be Used in Experimental Animal Studies? Pharm. Res. 2020, 37, 12. [Google Scholar] [CrossRef]
- Murota, K.; Nakamura, Y.; Uehara, M. Flavonoid Metabolism: The Interaction of Metabolites and Gut Microbiota. Biosci. Biotechnol. Biochem. 2018, 82, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Thangaraj, K.; Natesan, K.; Settu, K.; Palani, M.; Govindarasu, M.; Subborayan, V.; Vaiyapuri, M. Orientin Mitigates 1, 2-Dimethylhydrazine Induced Lipid Peroxidation, Antioxidant and Biotransforming Bacterial Enzyme Alterations in Experimental Rats. J. Cancer Res. Ther. 2018, 14, 1379–1388. [Google Scholar] [CrossRef]
- Nowak, B.; Poźniak, B.; Popłoński, J.; Bobak, Ł.; Matuszewska, A.; Kwiatkowska, J.; Dziewiszek, W.; Huszcza, E.; Szeląg, A. Pharmacokinetics of Xanthohumol in Rats of Both Sexes after Oral and Intravenous Administration of Pure Xanthohumol and Prenylflavonoid Extract. Adv. Clin. Exp. Med. 2020, 29, 1101–1109. [Google Scholar] [CrossRef]
- Neamtu, A.; Maghiar, T.; Alaya, A.; Olah, N.; Turcus, V.; Pelea, D.; Totolici, B.D.; Neamtu, C.; Maghiar, A.M.; Mathe, E. A Comprehensive View on the Quercetin Impact on Colorectal Cancer. Molecules 2022, 27, 1873. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Tripathi, A.; Chaudhari, B.P.; Dwivedi, P.D.; Pandey, H.P.; Das, M. Deoxynivalenol Induced Mouse Skin Cell Proliferation and Inflammation via MAPK Pathway. Toxicol. Appl. Pharmacol. 2014, 279, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Kim, M.Y.; Cho, J.Y. Anti-Inflammatory Effects of Luteolin: A Review of in Vitro, in Vivo, and in Silico Studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.C.; Kim, H.J.; Kim, Y.J.; Lee, K.Y.; Choi, H.J.; Lee, I.S.; Kang, B.Y. Differential Anti-Inflammatory Pathway by Xanthohumol in IFN-γ and LPS-Activated Macrophages. Int. Immunopharmacol. 2008, 8, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Chiang, M.C.; Tsai, T.Y.; Wang, C.J. The Potential Benefits of Quercetin for Brain Health: A Review of Anti-Inflammatory and Neuroprotective Mechanisms. Int. J. Mol. Sci. 2023, 24, 6328. [Google Scholar] [CrossRef] [PubMed]
- Carbonaro, M.; Grant, G. Absorption of Quercetin and Rutin in Rat Small Intestine. Ann. Nutr. Metab. 2005, 49, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.L.; Alvarado, D.A.; Swanson, K.S.; Holscher, H.D. The Prebiotic Potential of Inulin-Type Fructans: A Systematic Review. Adv. Nutr. 2022, 13, 492–529. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; Ledesma, E.; Monte, J.; Millán, E.; Costa, P.; de la Fuente, V.G.; García, M.T.F.; Martínez-Camblor, P.; Villar, C.J.; Lombó, F. Traditional Processed Meat Products Re-Designed Towards Inulin-Rich Functional Foods Reduce Polyps in Two Colorectal Cancer Animal Models. Sci. Rep. 2019, 9, 14783. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Metzler-Zebeli, B.U.; Zebeli, Q. Gut Function-Enhancing Properties and Metabolic Effects of Dietary Indigestible Sugars in Rodents and Rabbits. Nutrients 2015, 7, 8348–8365. [Google Scholar] [CrossRef]
- Oteiza, P.I.; Fraga, C.G.; Mills, D.A.; Taft, D.H. Flavonoids and the Gastrointestinal Tract: Local and Systemic Effects. Mol. Asp. Med. 2018, 61, 41–49. [Google Scholar] [CrossRef]
- Ashokkumar, P.; Sudhandiran, G. Luteolin Inhibits Cell Proliferation during Azoxymethane-Induced Experimental Colon Carcinogenesis via Wnt/ β-Catenin Pathway. Investig. New Drugs 2011, 29, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Matsukawa, Y.; Nishino, H.; Okuyama, Y.; Matsui, T.; Matsumoto, T.; Matsumura, S.; Shimizu, Y.; Sowa, Y.; Sakai, T. Effects of Quercetin and/or Restraint Stress on Formation of Aberrant Crypt Foci Induced by Azoxymethane in Rat Colons. Oncology 1997, 54, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Deschner, E.E.; Ruperto, J.; Wong, G.; Newmark, H.L. Quercetin and Rutin as Inhibitors of Azoxymethanol-Induced Colonic Neoplasia. Carcinogenesis 1991, 12, 1193–1196. [Google Scholar] [CrossRef] [PubMed]
- Torrens-Mas, M.; Alorda-Clara, M.; Martínez-Vigara, M.; Roca, P.; Sastre-Serra, J.; Oliver, J.; Pons, D.G. Xanthohumol Reduces Inflammation and Cell Metabolism in HT29 Primary Colon Cancer Cells. Int. J. Food Sci. Nutr. 2022, 73, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Sastre-Serra, J.; Ahmiane, Y.; Roca, P.; Oliver, J.; Pons, D.G. Xanthohumol, a Hop-Derived Prenylflavonoid Present in Beer, Impairs Mitochondrial Functionality of SW620 Colon Cancer Cells. Int. J. Food Sci. Nutr. 2019, 70, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.R.; Abensur, H. Physiology of Fluid and Solute Transport across the Peritoneal Membrane. J. Bras. Nefrol. 2014, 36, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M. Ellagitannins, Ellagic Acid and Their Derived Metabolites: A Review about Source, Metabolism, Functions and Health. Food Res. Int. 2011, 44, 1150–1160. [Google Scholar] [CrossRef]
- Jaganath, I.B.; Jaganath, I.B.; Mullen, W.; Edwards, C.A.; Crozier, A. The Relative Contribution of the Small and Large Intestine to the Absorption and Metabolism of Rutin in Man. Free Radic. Res. 2006, 40, 1035–1046. [Google Scholar] [CrossRef]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of Dietary Polyphenols and Gut Microbiota Metabolism: Antimicrobial Properties. BioMed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef]
- Selma, M.V.; Espín, J.C.; Tomás-Barberán, F.A. Interaction between Phenolics and Gut Microbiota: Role in Human Health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Feo, V.; Battistelli, A.; Da Cruz, A.G.; Coppola, R. Polyphenols, the New Frontiers of Prebiotics, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Volume 94, ISBN 9780128202180. [Google Scholar]
- Makarewicz, M.; Drożdż, I.; Tarko, T.; Duda-Chodak, A. The Interactions between Polyphenols and Microorganisms, Especially Gut Microbiota. Antioxidants 2021, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Jin, Z.; Wu, W.; Gao, R.; Guo, B.; Gao, Z.; Yang, Y.; Qin, H. Analysis of the Intestinal Lumen Microbiota in an Animal Model of Colorectal Cancer. PLoS ONE 2014, 9, e90849. [Google Scholar] [CrossRef]
- Yazici, C.; Wolf, P.G.; Kim, H.; Cross, T.-W.L.; Vermillion, K.; Carroll, T.; Augustus, G.J.; Mutlu, E.; Tussing-Humphreys, L.; Braunschweig, C.; et al. Race-Dependent Association of Sulfidogenic Bacteria with Colorectal Cancer. Gut 2017, 66, 1983–1994. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, G.; Guttula, K.; Kellingray, L.; Tett, A.J.; ten Hoopen, R.; Kemsley, E.K.; Savva, G.M.; Ibrahim, A.; Narbad, A. Metabolite Quantification of Faecal Extracts from Colorectal Cancer Patients and Healthy Controls. Oncotarget 2018, 9, 33278–33289. [Google Scholar] [CrossRef]
- Burns, M.B.; Montassier, E.; Abrahante, J.; Priya, S.; Niccum, D.E.; Khoruts, A.; Starr, T.K.; Knights, D.; Blekhman, R. Colorectal Cancer Mutational Profiles Correlate with Defined Microbial Communities in the Tumor Microenvironment. PLoS Genet. 2018, 14, e1007376. [Google Scholar] [CrossRef]
- Waters, J.L.; Ley, R.E. The Human Gut Bacteria Christensenellaceae Are Widespread, Heritable, and Associated with Health. BMC Biol. 2019, 17, 83. [Google Scholar] [CrossRef]
- Qu, R.; Zhang, Y.; Ma, Y.; Zhou, X.; Sun, L.; Jiang, C.; Zhang, Z.; Fu, W. Role of the Gut Microbiota and Its Metabolites in Tumorigenesis or Development of Colorectal Cancer. Adv. Sci. 2023, 10, e2205563. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; Redondo-Blanco, S.; Gutiérrez-del-Río, I.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Colon Microbiota Fermentation of Dietary Prebiotics towards Short-Chain Fatty Acids and Their Roles as Anti-Inflammatory and Antitumour Agents: A Review. J. Funct. Foods 2016, 25, 511–522. [Google Scholar] [CrossRef]
- Liu, N.; Zou, S.; Xie, C.; Meng, Y.; Xu, X. Effect of the β-Glucan from Lentinus Edodes on Colitis-Associated Colorectal Cancer and Gut Microbiota. Carbohydr. Polym. 2023, 316, 121069. [Google Scholar] [CrossRef]
- Peck, S.C.; Denger, K.; Burrichter, A.; Irwin, S.M.; Balskus, E.P.; Schleheck, D. A Glycyl Radical Enzyme Enables Hydrogen Sulfide Production by the Human Intestinal Bacterium Bilophila Wadsworthia. Proc. Natl. Acad. Sci. USA 2019, 116, 3171–3176. [Google Scholar] [CrossRef]
- Mancabelli, L.; Milani, C.; Lugli, G.A.; Turroni, F.; Cocconi, D.; van Sinderen, D.; Ventura, M. Identification of Universal Gut Microbial Biomarkers of Common Human Intestinal Diseases by Meta-Analysis. FEMS Microbiol. Ecol. 2017, 93, fix153. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nan, X.; Zhao, Y.; Jiang, L.; Wang, H.; Zhang, F.; Hua, D.; Liu, J.; Yao, J.; Yang, L.; et al. Dietary Supplementation of Inulin Ameliorates Subclinical Mastitis via Regulation of Rumen Microbial Community and Metabolites in Dairy Cows. Microbiol. Spectr. 2021, 9, e0010521. [Google Scholar] [CrossRef]
- Lin, H.; Ma, X.; Yang, X.; Chen, Q.; Wen, Z.; Yang, M.; Fu, J.; Yin, T.; Lu, G.; Qi, J.; et al. Natural Shikonin and Acetyl-Shikonin Improve Intestinal Microbial and Protein Composition to Alleviate Colitis-Associated Colorectal Cancer. Int. Immunopharmacol. 2022, 111, 109097. [Google Scholar] [CrossRef]
- Weber, A.M.; Ibrahim, H.; Baxter, B.A.; Kumar, R.; Maurya, A.K.; Kumar, D.; Agarwal, R.; Raina, K.; Ryan, E.P. Integrated Microbiota and Metabolite Changes Following Rice Bran Intake during Murine Inflammatory Colitis-Associated Colon Cancer and in Colorectal Cancer Survivors. Cancers 2023, 15, 2231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, Z.; Lu, Y.; Tu, D.; Zou, F.; Lin, S.; Yu, W.; Miao, M.; Shi, H. A Functional Food Inhibits Azoxymethane/Dextran Sulfate Sodium-Induced Inflammatory Colorectal Cancer in Mice. Onco. Targets Ther. 2021, 14, 1465–1477. [Google Scholar] [CrossRef]
- Bosshard, P.P.; Zbinden, R.; Altwegg, M. Turicibacter Sanguinis Gen. Nov., Sp. Nov., a Novel Anaerobic, Gram-Positive Bacterium. Int. J. Syst. Evol. Microbiol. 2002, 52, 1263–1266. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Ryu, Y.; An, B.C.; Yoon, Y.S.; Choi, O.; Kim, T.Y.; Yoon, J.; Ahn, J.Y.; Park, H.J.; Kwon, S.K.; et al. A Synthetic Probiotic Engineered for Colorectal Cancer Therapy Modulates Gut Microbiota. Microbiome 2021, 9, 122. [Google Scholar] [CrossRef] [PubMed]
- Zackular, J.P.; Baxter, N.T.; Iverson, K.D.; Sadler, W.D.; Petrosino, J.F.; Chen, G.Y.; Schloss, P.D. The Gut Microbiome Modulates Colon Tumorigenesis. mBio 2013, 4, e00692-13. [Google Scholar] [CrossRef] [PubMed]
- Jahani-Sherafat, S.; Alebouyeh, M.; Moghim, S.; Amoli, H.A.; Ghasemian-Safaei, H. Role of Gut Microbiota in the Pathogenesis of Colorectal Cancer: A Review Article. Gastroenterol. Hepatol. Bed Bench 2018, 11, 101. [Google Scholar]
- Cipe, G.; Idiz, U.O.; Firat, D.; Bektasoglu, H. Relationship Between Intestinal Microbiota and Colorectal Cancer. World J. Gastrointest. Oncol. 2015, 7, 233–240. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort 1 | Cohort 2 | Cohort 3 | |
|---|---|---|---|
| Cohort 2 | 0.037 | ||
| Cohort 3 | 0.002 | 0.002 | |
| Cohort 4 | 0.0084 | 0.0075 | 0.002 |
| Cohort 1 (%) | Cohort 2 (%) | Cohort 3 (%) | Cohort 4 (%) | |
|---|---|---|---|---|
| Actinomycetota | 0.33 | 0.30 | 0.10 **** | 0.26 |
| Bacteroidota | 32.01 | 32.40 | 36.08 ** | 35.80 ** |
| Cyanobacteria | 0.27 | 0.34 | 0.69 | 1.06 * |
| Deferribacterota | 0.31 | 0.34 | 0.43 | 0.19 |
| Thermodesulfobacteriota | 0.53 | 0.43 | 0.95 | 0.60 |
| Bacillota | 64.78 | 63.33 | 59.87 * | 59.41 * |
| Patescibacteria | 0.35 | 2.06 ** | 1.07 | 1.28 * |
| Pseudomonadota | 0.80 | 0.15 ** | 0.34 | 0.87 |
| Verrucomicrobiota | 0.62 | 0.67 | 0.48 | 0.52 |
| Cohort 1 (%) | Cohort 2 (%) | Cohort 3 (%) | Cohort 4 (%) | |
|---|---|---|---|---|
| Eggerthellaceae | 0.24 | 0.16 | 0.05 **** | 0.18 |
| Bacteroidaceae | 5.26 | 4.21 | 5.42 | 6.12 |
| Muribaculaceae | 15.50 | 16.42 | 14.61 | 14.61 |
| Prevotellaceae | 6.83 | 7.93 | 11.09 **** | 10.80 **** |
| Rikenellaceae | 2.76 | 2.34 | 3.40 | 2.38 |
| Tannerellaceae | 1.63 | 1.49 | 1.49 | 1.88 |
| Gastranaerophilales | 0.27 | 0.34 | 0.69 | 1.06 ** |
| Deferribacteraceae | 0.31 | 0.34 | 0.43 | 0.19 |
| Desulfovibrionaceae | 0.53 | 0.43 | 0.95 | 0.60 |
| Acholeplasmataceae | 0.30 | 0.17 | 0.15 | 0.14 |
| Bacillaceae | 0.85 | 0.53 | 0.49 | 0.30 |
| Erysipelatoclostridiaceae | 0.17 | 0.05 | 0.04 | 0.24 |
| Erysipelotrichaceae | 0.82 | 0.60 | 0.13 *** | 0.43 |
| Lactobacillaceae | 2.93 | 3.20 | 2.35 | 2.97 |
| RF39 (Bacillota) | 0.25 | 0.29 | 0.11 | 0.24 |
| Christensenellaceae | 0.77 | 0.28 | 0.09 **** | 0.67 |
| Clostridia UCG-014 | 4.37 | 3.64 | 1.88 | 4.94 |
| Clostridia vadin BB60 group | 0.19 | 0.09 | 0.88 * | 0.11 |
| Lachnospiraceae | 26.07 | 27.94 | 24.93 | 21.84 |
| Monoglobaceae | 0.25 | 0.23 | 0.20 | 0.13 |
| Butyricicoccaceae | 0.23 | 0.26 | 0.61 ** | 0.37 |
| Oscillospiraceae | 13.11 | 13.43 | 15.38 | 13.48 |
| Ruminococcaceae | 11.36 | 10.31 | 10.86 | 10.80 |
| Oscillospirales UCG-010 | 0.21 | 0.18 | 0.25 | 0.22 |
| Coprostanoligenes group | 1.26 | 1.06 | 0.67 | 1.09 |
| Peptococcaceae | 0.72 | 0.65 | 0.52 | 0.62 |
| Anaerovoracaceae | 0.26 | 0.16 | 0.10 **** | 0.16 |
| Peptostreptococcaceae | 0.58 | 0.21 | 0.25 | 0.53 |
| Saccharimonadaceae | 0.35 | 2.06 * | 1.07 | 1.28 * |
| Uncultured (Rhodospirillales) | 0.03 | 0.02 | 0.17 | 0.22 |
| Sutterellaceae | 0.01 | 0.02 | 0.03 | 0.16 *** |
| Enterobacteriaceae | 0.69 | 0.07 * | 0.08 * | 0.31 |
| Moraxellaceae | 0.00 | 0.00 | 0.00 | 0.12 |
| Akkermansiaceae | 0.62 | 0.67 | 0.48 | 0.52 |
| Genus | Cohort 1 (%) | Cohort 2 (%) | Cohort 3 (%) | Cohort 4 (%) |
|---|---|---|---|---|
| Muribaculum | 0.24 | 0.37 *** | 0.56 **** | 0.44 **** |
| Bilophila | 0.18 | 0.29 | 0.80 *** | 0.49 |
| Christensenellaceae R-7 | 0.76 | 0.27 | 0.08 ** | 0.66 |
| Clostridia UCG-014 | 4.37 | 3.64 | 1.88 ** | 4.94 |
| Eubacterium xylanophilum group | 1.40 | 0.71 * | 0.31 ** | 0.69 * |
| Uncultured (Oscillospiraceae) | 3.99 | 4.84 | 6.06 * | 4.72 |
| Uncultured (Ruminococcaceae) | 0.45 | 0.46 | 1.27 **** | 0.98 ** |
| “Candidatus Saccharimonas” | 0.35 | 2.06 ** | 1.07 | 1.28 |
| Parasutterella | 0.01 | 0.02 | 0.03 | 0.16 * |
| Turicibacter | 0.77 | 0.46 | 0.07 ** | 0.40 |
| Gastranaerophilales | 0.20 | 0.25 | 0.22 | 0.71 * |
| Uncultured (Erysipelotrichaceae) | 0.02 | 0.13 * | 0.06 | 0.02 |
| Species | ||||
| Bacteroides sp. | 0.23 | 0.27 | 1.45 ** | 1.04 |
| Eubacterium sp. | 0.06 | 0.33 | 0.30 | 0.58 * |
| Coprostanoligenes group | ||||
| UCG-005 metagenome | 0.05 | 0.13 * | 0.08 | 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Valero, Á.; Magadán-Corpas, P.; Ye, S.; Serna-Diestro, J.; Sordon, S.; Huszcza, E.; Popłoński, J.; Villar, C.J.; Lombó, F. Antitumor Effect and Gut Microbiota Modulation by Quercetin, Luteolin, and Xanthohumol in a Rat Model for Colorectal Cancer Prevention. Nutrients 2024, 16, 1161. https://doi.org/10.3390/nu16081161
Pérez-Valero Á, Magadán-Corpas P, Ye S, Serna-Diestro J, Sordon S, Huszcza E, Popłoński J, Villar CJ, Lombó F. Antitumor Effect and Gut Microbiota Modulation by Quercetin, Luteolin, and Xanthohumol in a Rat Model for Colorectal Cancer Prevention. Nutrients. 2024; 16(8):1161. https://doi.org/10.3390/nu16081161
Chicago/Turabian StylePérez-Valero, Álvaro, Patricia Magadán-Corpas, Suhui Ye, Juan Serna-Diestro, Sandra Sordon, Ewa Huszcza, Jarosław Popłoński, Claudio J. Villar, and Felipe Lombó. 2024. "Antitumor Effect and Gut Microbiota Modulation by Quercetin, Luteolin, and Xanthohumol in a Rat Model for Colorectal Cancer Prevention" Nutrients 16, no. 8: 1161. https://doi.org/10.3390/nu16081161
APA StylePérez-Valero, Á., Magadán-Corpas, P., Ye, S., Serna-Diestro, J., Sordon, S., Huszcza, E., Popłoński, J., Villar, C. J., & Lombó, F. (2024). Antitumor Effect and Gut Microbiota Modulation by Quercetin, Luteolin, and Xanthohumol in a Rat Model for Colorectal Cancer Prevention. Nutrients, 16(8), 1161. https://doi.org/10.3390/nu16081161