Interactions between C-Reactive Protein Genotypes with Markers of Nutritional Status in Relation to Inflammation

 ,
, 
Abstract
:1. Introduction
2. Experimental Section
2.1. Study Design, Population Selection and Demographic Characteristics
2.2. Ethics
2.3. Lifestyle Factors and Dietary Intake Assessment and Analysis
2.4. Anthropometrical and Blood Pressure Assessment
2.5. Blood Sampling and Storage
2.6. Biochemical Analyses
2.7. Statistical Analysis
3. Results
3.1. Characteristics of Study Participants and Correlations between Measured Variables and CRP Concentrations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Participant’s Characteristics | Whole Group (n = 2010) | Correlation with CRP | |||
|---|---|---|---|---|---|
| r | p | ||||
| Age (year) | 48.0 (41.0–56.0) | 0.11 | <0.001 | ||
| Blood pressure (mmHg) | Diastolic | 87.0 (78.0–97.0) | 0.07 | <0.001 | |
| Systolic | 130 (116–147) | 0.05 | 0.03 | ||
| Tobacco use (%) | Current | 52.7 | 0.02 | 0.30 | |
| Former | 4.14 | ||||
| Never | 42.7 | ||||
| Physical activity | 7.33 (5.79–8.68) | −0.11 | <0.001 | ||
| Anthropo-metrical markers | BMI (kg/m2) | 22.9 (19.3–28.6) | 0.22 | <0.001 | |
| Waist circumference (cm) | 77.5 (70.2–87.7) | 0.27 | <0.001 | ||
| Hip circumference (cm) | 93.1 (84.8–106) | 0.20 | <0.001 | ||
| Waist-to-hip ratio | 0.83 (0.78–0.88) | 0.09 | <0.001 | ||
| Upper-arm circumference (cm) | 27.5 (24.3–31.3) | 0.20 | <0.001 | ||
| Biochemical markers | HIV status (n) (−/+/do not know) | 1668/326/14 | 0.03 | 0.14 | |
| TC (mmol/L) | 4.82 (4.01–5.87) | 0.03 | 0.21 | ||
| LDL-C (mmol/L) | 2.79 (2.08–3.65) | 0.07 | 0.004 | ||
| HDL-C (mmol/L) | 1.42 (1.06–1.87) | −0.14 | <0.001 | ||
| Triglycerides (mmol/L) | 1.08 (0.82–1.55) | 0.13 | <0.001 | ||
| Fasting glucose (mmol/L) | 4.80 (4.30–5.30) | 0.07 | 0.003 | ||
| HbA1c (%) | 5.50 (5.30–5.80) | 0.21 | <0.001 | ||
| CRP (mg/L) | 3.29 (0.96–9.34) | ||||
| Dietary intake | Energy (KJ) | 7175 (5268–10001) | 0.05 | 0.05 | |
| Protein (%TE) | 11.6 (10.4–12.9) | 0.01 | 0.60 | ||
| Carbohydrate (%TE) | 60.3 (54.2–67.5) | −0.01 | 0.57 | ||
| Added sugar (%TCHO) | 15.2 (9.51–21.8) | 0.05 | 0.05 | ||
| Total fat (%TE) | 22.5 (17.4 – 27.7) | 0.03 | 0.23 | ||
| SFA (%TE) | 5.29 (3.63–7.06) | 0.03 | 0.17 | ||
| MUFA (%TE) | 5.77 (3.75–7.74) | 0.05 | 0.05 | ||
| MUFA-to-SFA ratio (g:g) | 1.13 (0.97–1.27) | 0.03 | 0.24 | ||
| PUFA | PUFA (%TE) | 6.78 (5.06–8.60) | 0.03 | 0.2 | |
| Omega-6 (g) | 12 (6.98–18.5) | 0.003 | 0.91 | ||
| Omega-3 (g) | 0.34 (0.20–0.52) | 0.004 | 0.88 | ||
| Omega-6 to -3 ratio (g:g) | 36.1 (26.1–50.3) | 0.005 | 0.83 | ||
| Cholesterol (mg) | 150 (80.0–259) | 0.07 | 0.002 | ||
| Alcohol intake (g/day) | 11.5 ± 22.7 | −0.01 | 0.58 | ||
| Abstainers:drinkers (n:n) | 824:659 | ||||
| Dietary fiber (g) | Total | 19.1 (13.7–27.0) | 0.02 | 0.29 | |
| Insoluble | 2.02 (1.15–3.31) | 0.08 | <0.001 | ||
| Soluble | 1.46 (0.84–2.44) | 0.08 | <0.001 | ||
| Vitamin C (mg) | 17.7 (10.4–38.1) | 0.07 | 0.003 | ||
| Vitamin E (mg) | 9.67 (5.92–14.2) | 0.03 | 0.26 | ||
| Mg (mg) | 285 (199–406) | 0.03 | 0.20 | ||
| Zinc (mg) | 8.90 (6.49–12.6) | 0.03 | 0.15 | ||
| Carotenoids (μg) | 1117 (303–2305) | 0.09 | <0.001 | ||
| Fruit and vegetables (g) | 86.6 (51.9–154) | 0.08 | <0.001 | ||
| Pulses, nuts and seeds (g) | 4.29 (0–19.3) | 0.02 | 0.49 | ||
| Adapted Thiele dietary quality score | 1490 (1275–1630) | 0.07 | 0.003 | ||
| Healthy diet indicator | 6.68 (6.11–7.21) | 0.03 | 0.14 | ||
| CRP Gene Polymorphisms | Genotype | MAF% | ΔCRP |
|---|---|---|---|
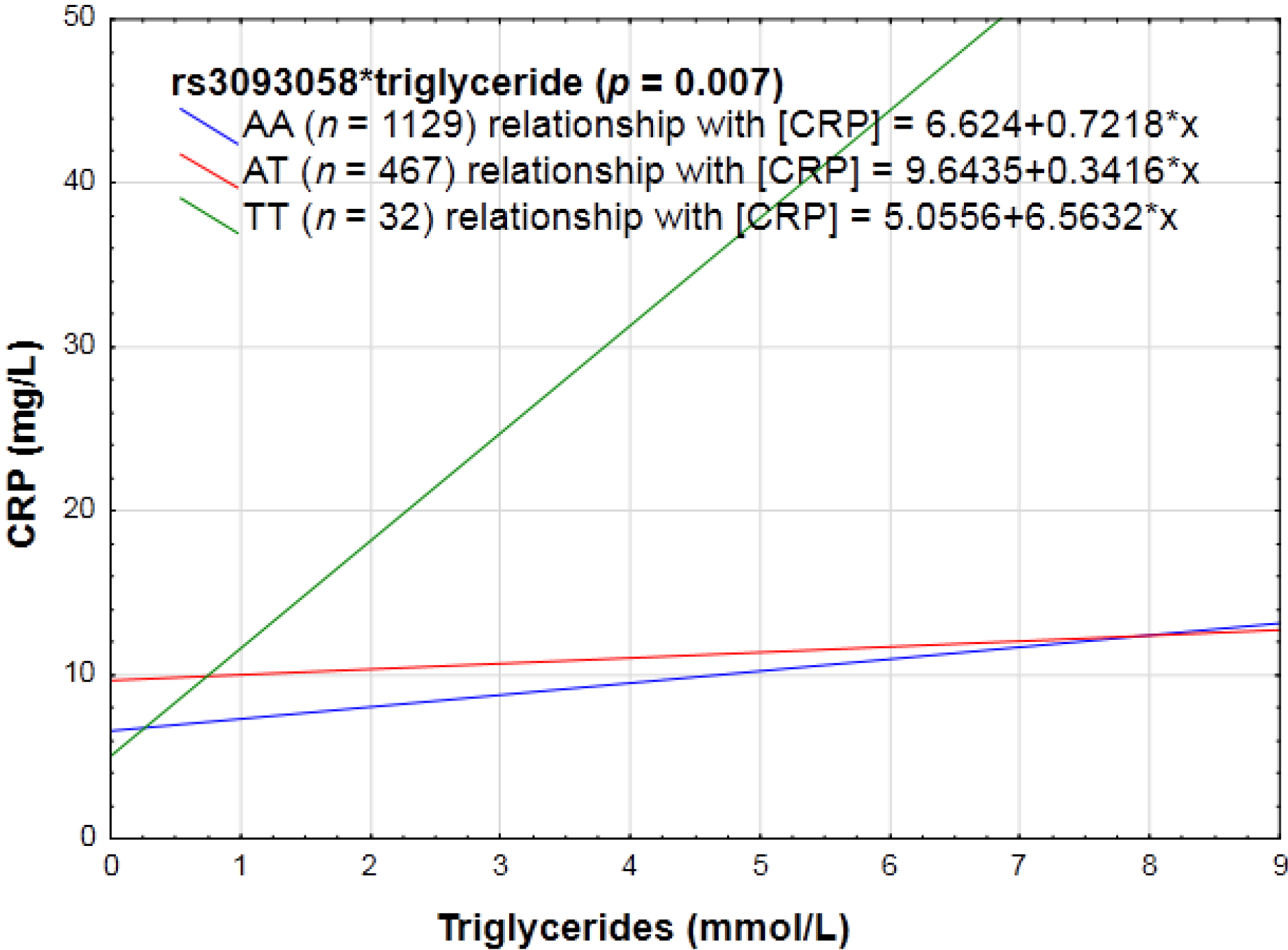
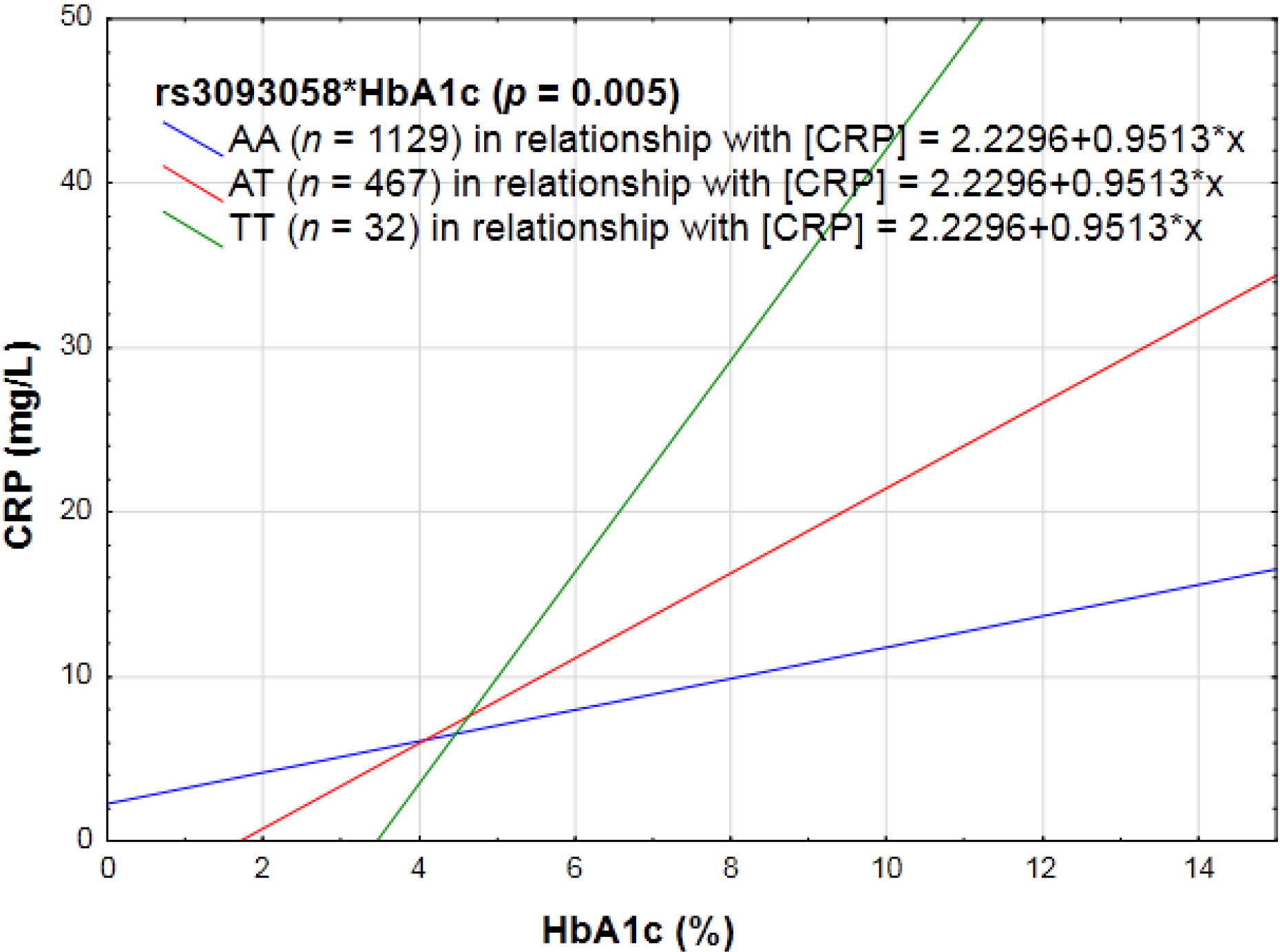
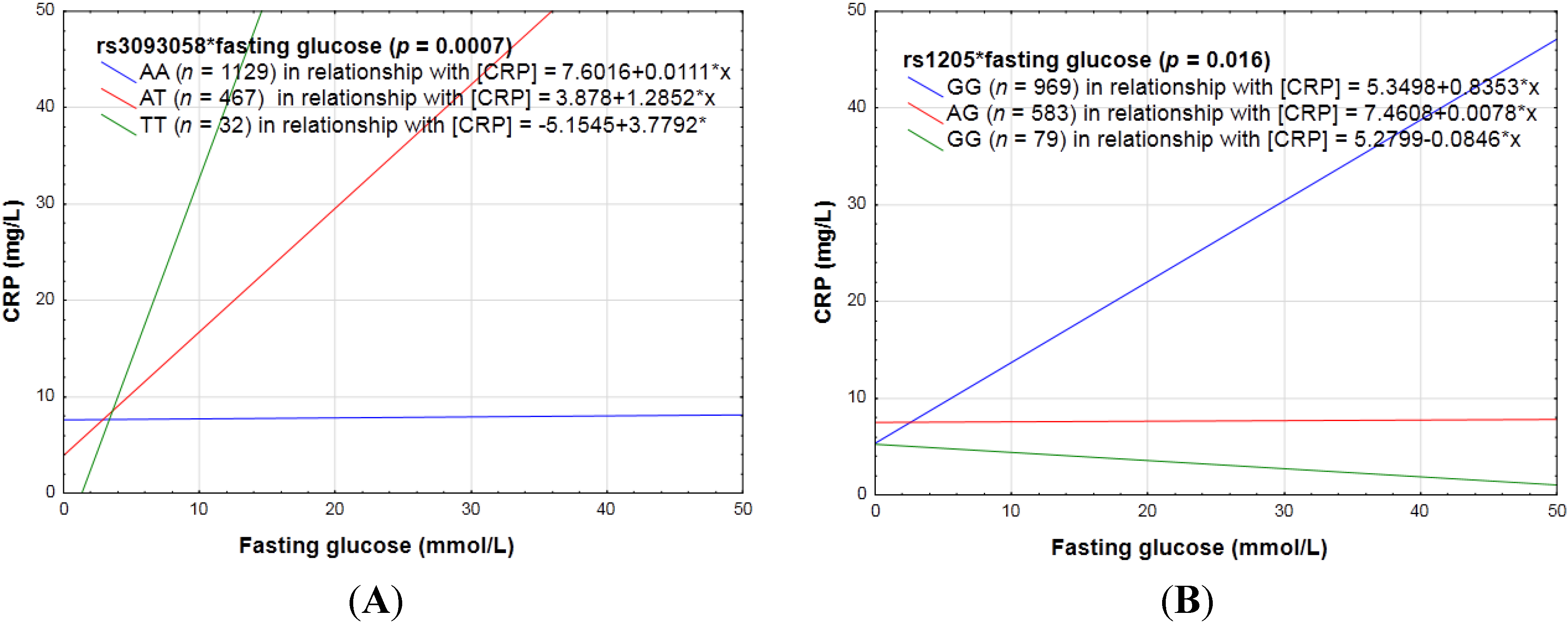
| rs3093058 (−790A > T) | AA | T = 19.4 | Increase |
| AT | |||
| TT | |||
| rs1205 (3′ UTR) (3u2131C > T or 3872A > G or +2147G > A) | GG | A = 22.8 | Decrease |
| GA | |||
| AA | |||
| rs7553007 | GG | A = 23.8 | Decrease |
| GA | |||
| AA | |||
| rs2794520 | GG | A = 23.6 | Decrease |
| GA | |||
| AA | |||
| rs2808630 (+5237A > G) | AA | G = 14.3 | No change |
| AG | |||
| GG | |||
| rs3093068 (+2911C > G) | CC | G = 37.5 | Increase |
| CG | |||
| GG | |||
| rs1417938 (intron) (i178T > A) | TT | A = 97.5 | No change |
| TA | |||
| rs3093062 (−409G > A) | GG | A = 16.2 | Increase |
| GA | |||
| AA | |||
| rs2027471 | TT | A = 23.5 | Decrease |
| TA | |||
| AA | |||
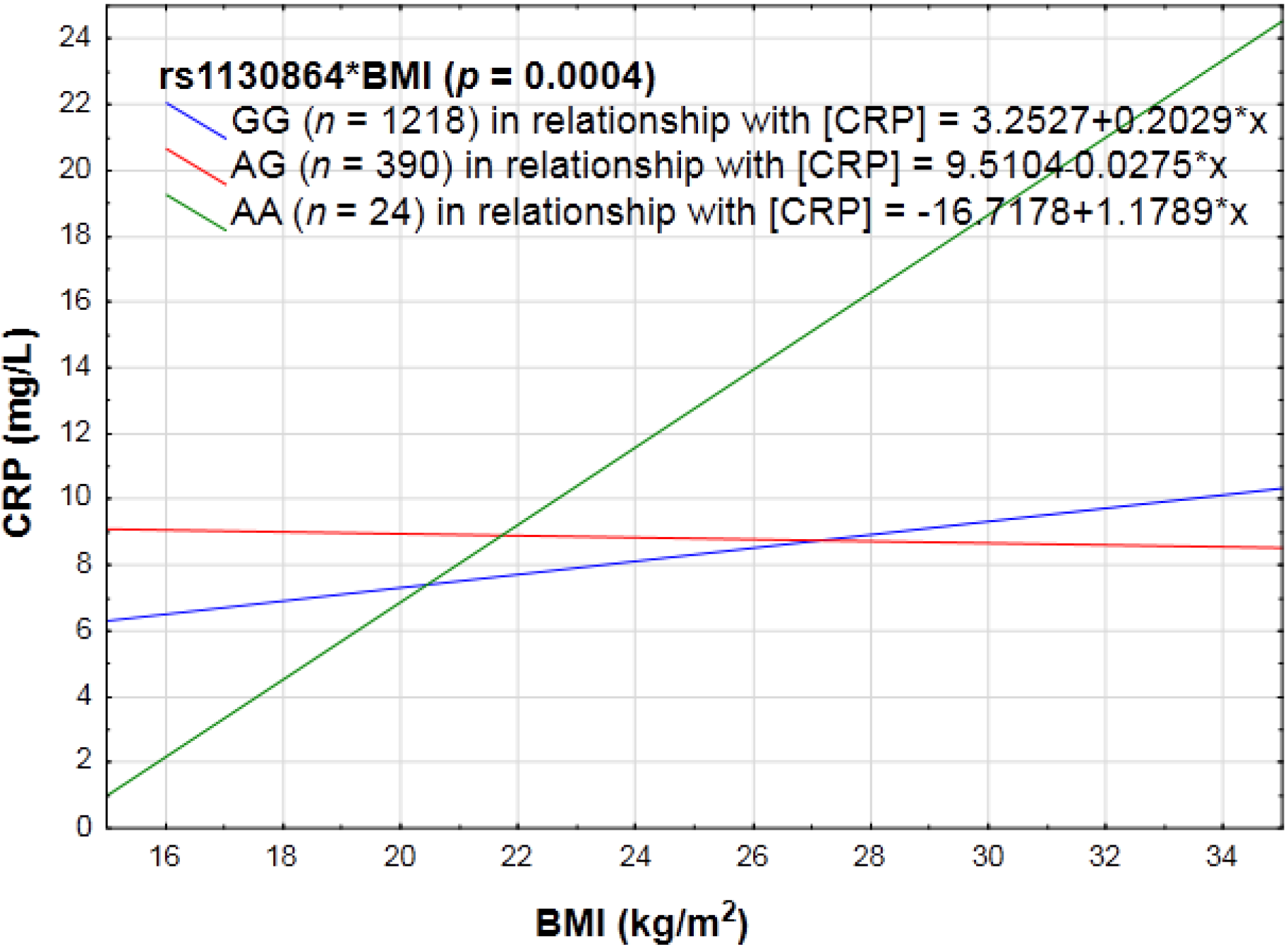
| rs1130864 (3′ UTR) * (3u1273C > T or +1444C > T) | GG | A = 13.4 | No change |
| GA | |||
| AA | |||
| rs1341665 * (−7180C > T) | GG | A = 23.5 | Decrease |
| GA | |||
| AA | |||
| rs1800947 (exon 2) (L184L or +1059C > G) | CC | C = 100 | No change |
| CG |
3.2. Interaction Effects of Various Markers of Nutritional Status with CRP SNPs in Relation to the CRP Phenotype
3.2.1. PA, Anthropometry and CRP Genotype Interactions in Relation to Inflammation

3.2.2. Biochemical Markers of the Lipid Profile, as Well as Glycemic Control (HbA1c and Fasting Glucose) and CRP Genotype Interactions in Relation to Inflammation



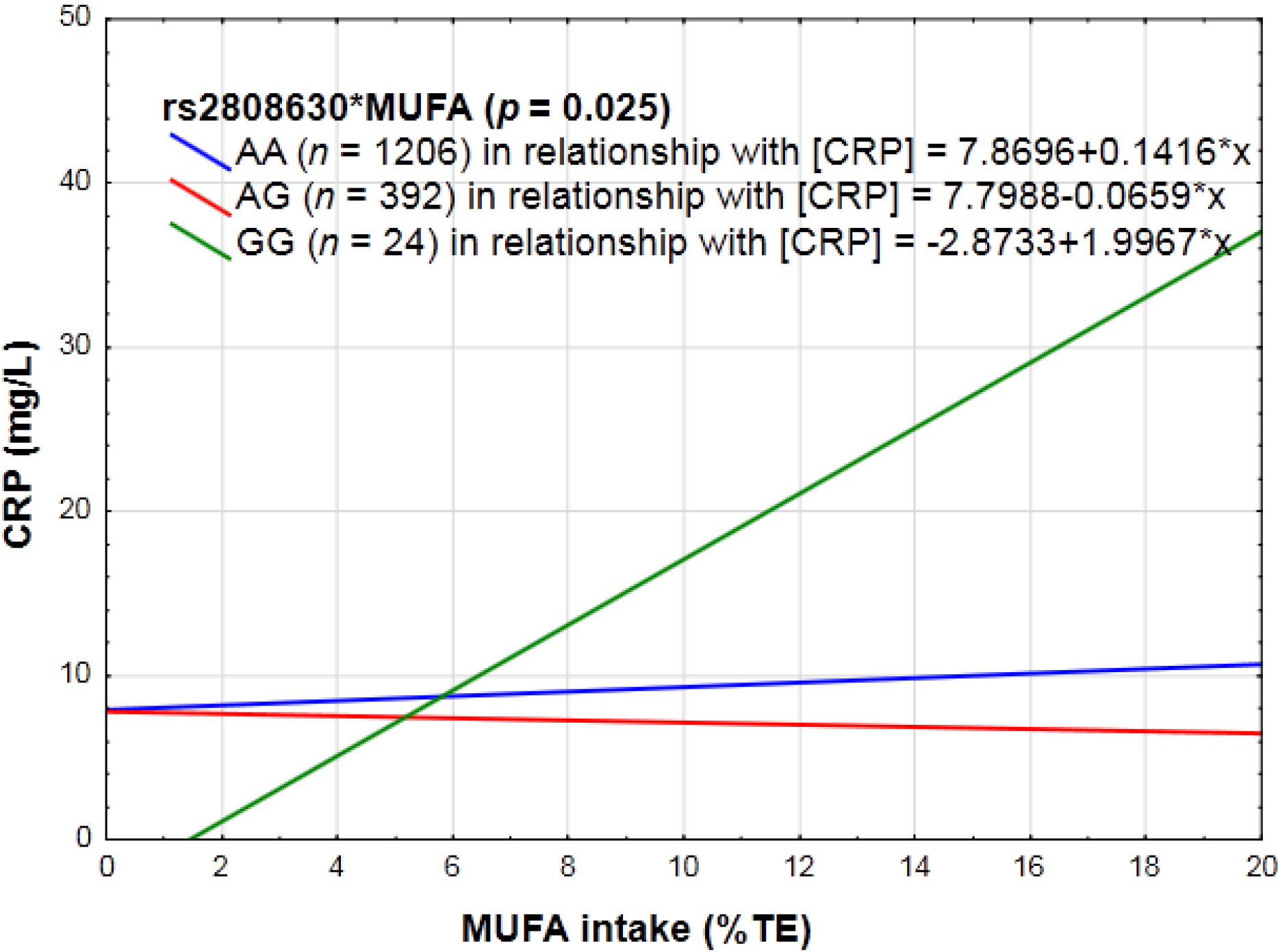
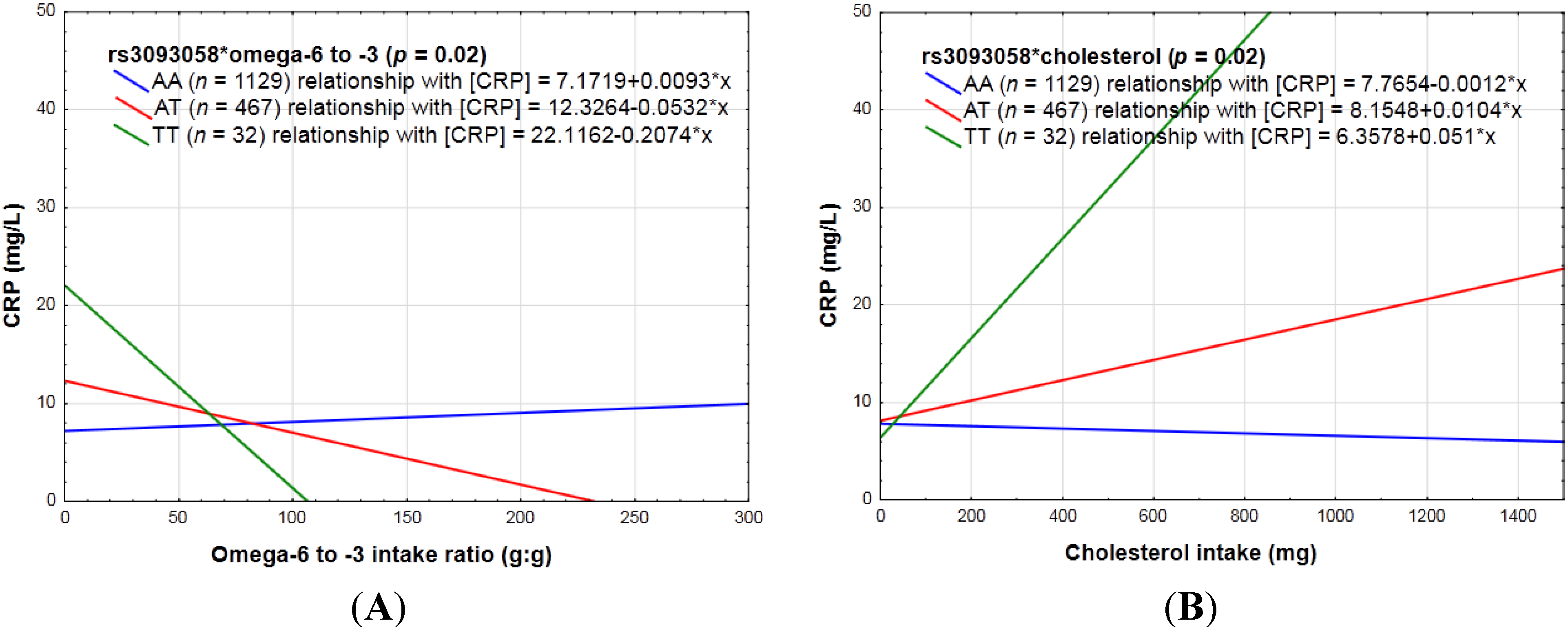
3.2.3. Dietary Sugar Intake, Fat Intake, Absolute Alcohol Intake, Dietary Fiber, Vitamin C and E, Magnesium, Zinc, Carotenoid, Fruit and Vegetables, Pulses, Nuts and Seeds, as Well as Diet Quality and CRP Genotype Interactions in Relation to Inflammation


4. Discussion
4.1. Anthropometry and PA with CRP Genotype Interactions in Relation to Inflammation
4.2. Biochemical Markers and CRP Genotype Interactions in Relation to Inflammation
4.3. Specific Dietary Components as Well as Diet Quality Scores and CRP Genotype Interactions in Relation to Inflammation
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Berk, M.; Williams, L.J.; Jacka, F.N.; O’Neil, A.; Pasco, J.A.; Moylan, S.; Allen, N.B.; Stuart, A.L.; Hayley, A.C.; Byrne, M.L.; et al. So depression is an inflammatory disease, but where does the inflammation come from? BMC Med. 2013, 11, 200. [Google Scholar] [CrossRef]
- Duncan, B.B.; Schmidt, M.I.; Pankow, J.S.; Ballantyne, C.M.; Couper, D.; Vigo, A.; Hoogeveen, R.; Folsom, A.R.; Heiss, G. Low-grade systemic inflammation and the development of type 2 diabetes the atherosclerosis risk in communities study. Diabetes 2003, 52, 1799–1805. [Google Scholar] [CrossRef] [PubMed]
- Schaumberg, D.A.; Christen, W.G.; Buring, J.E.; Glynn, R.J.; Rifai, N.; Ridker, P.M. High-sensitivity C-reactive protein, other markers of inflammation, and the incidence of macular degeneration in women. Arch. Ophthalmol. 2007, 125, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Tsilidis, K.K.; Branchini, C.; Guallar, E.; Helzlsouer, K.J.; Erlinger, T.P.; Platz, E.A. C‐reactive protein and colorectal cancer risk: A systematic review of prospective studies. Int. J. Cancer 2008, 123, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Buckley, D.I.; Fu, R.; Freeman, M.; Rogers, K.; Helfand, M. C-reactive protein as a risk factor for coronary heart disease: A systematic review and meta-analyses for the US preventive services task force. Ann. Intern. Med. 2009, 151, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Danesh, J.; Wheeler, J.G.; Hirschfield, G.M.; Eda, S.; Eiriksdottir, G.; Rumley, A.; Lowe, G.D.; Pepys, M.B.; Gudnason, V. C-reactive protein and other circulating markers of inflammation in the prediction of coronary heart disease. N. Engl. J. Med. 2004, 350, 1387–1397. [Google Scholar] [CrossRef] [PubMed]
- Gaesser, G.A.; Angadi, S.S.; Ryan, D.M.; Johnston, C.S. Lifestyle measures to reduce inflammation. Am. J. Lifestyle Med. 2012, 6, 4–13. [Google Scholar] [CrossRef]
- Kadoglou, N.P.; Iliadis, F.; Angelopoulou, N.; Perrea, D.; Ampatzidis, G.; Liapis, C.D.; Alevizos, M. The anti-inflammatory effects of exercise training in patients with type 2 diabetes mellitus. Eur. J. Cardiovasc. Prev. Rehabil. 2007, 14, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Nicklas, B.J.; You, T.; Pahor, M. Behavioural treatments for chronic systemic inflammation: Effects of dietary weight loss and exercise training. Can. Med. Assoc. J. 2005, 172, 1199–1209. [Google Scholar] [CrossRef]
- Thompson, D.; Markovitch, D.; Betts, J.A.; Mazzatti, D.; Turner, J.; Tyrrell, R.M. Time course of changes in inflammatory markers during a 6-mo exercise intervention in sedentary middle-aged men: A randomized-controlled trial. J. Appl. Physiol. 2010, 108, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Devaraj, S.; Jialal, I. Dietary factors that promote or retard inflammation. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Devaraj, S.; Jialal, I. Alpha tocopherol supplementation decreases serum C-reactive protein and monocyte interleukin-6 levels in normal volunteers and type 2 diabetic patients. Free Radic. Biol. Med. 2000, 29, 790–792. [Google Scholar] [CrossRef] [PubMed]
- Galland, L. Diet and inflammation. Nutr. Clin. Pract. 2010, 25, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Esposito, K.; Marfella, R.; Ciotola, M.; di Palo, C.; Giugliano, F.; Giugliano, G.; D’Armiento, M.; D’Andrea, F.; Giugliano, D. Effect of a Mediterranean-style diet on endothelial dysfunction and markers of vascular inflammation in the metabolic syndrome: A randomized trial. JAMA 2004, 292, 1440–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, C.S.; Aldred, S.F.; Lee, P.K.; Tracy, R.P.; Schwartz, S.M.; Rieder, M.; Liu, K.; Williams, O.D.; Iribarren, C.; Lewis, E.C. Polymorphisms within the C-reactive protein (CRP) promoter region are associated with plasma CRP levels. Am. J. Hum. Genet. 2005, 77, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Kivimäki, M.; Lawlor, D.A.; Smith, G.D.; Eklund, C.; Hurme, M.; Lehtimäki, T.; Viikari, J.S.; Raitakari, O.T. Variants in the CRP gene as a measure of lifelong differences in average C-reactive protein levels the cardiovascular risk in young Finns study, 1980–2001. Am. J. Epidemiol. 2007, 166, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Teo, K.; Chow, C.K.; Vaz, M.; Rangarajan, S.; Yusuf, S. The Prospective Urban Rural Epidemiology (PURE) study: Examining the impact of societal influences on chronic noncommunicable diseases in low-, middle-, and high-income countries. Am. Heart J. 2009, 158, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Vaz, M.; Karaolis, N.; Draper, A.; Shetty, P. A compilation of energy costs of physical activities. Public Health Nutr. 2005, 8, 1153–1183. [Google Scholar] [CrossRef] [PubMed]
- MacIntyre, U.; Venter, C.; Vorster, H. A culture-sensitive quantitative food frequency questionnaire used in an African population: 1. Development and reproducibility. Public Health Nutr. 2001, 4, 53–62. [Google Scholar] [PubMed]
- MacIntyre, U.; Venter, C.; Vorster, H. A culture-sensitive quantitative food frequency questionnaire used in an African population: 2. Relative validation by 7-day weighed records and biomarkers. Public Health Nutr. 2001, 4, 63–71. [Google Scholar] [PubMed]
- Venter, C.; MacIntyre, U.; Vorster, H. The development and testing of a food portion photograph book for use in an African population. J. Hum. Nutr. Diet. 2000, 13, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Dolman, R.C.; Wentzel-Viljoen, E.; Jerling, J.C.; Feskens, E.J.; Kruger, A.; Pieters, M. The use of predefined diet quality scores in the context of CVD risk during urbanization in the South African Prospective Urban and Rural Epidemiological (PURE) study. Public Health Nutr. 2014, 17, 1706–1716. [Google Scholar] [CrossRef] [PubMed]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [PubMed]
- National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov/projects/SNP/ (accessed on 30 May 2014).
- Hardy, G.H. Mendelian proportions in a mixed population. Science 1908, 28, 49–50. [Google Scholar] [CrossRef] [PubMed]
- Haploview. Available online: http://www.broad.mit.edu/mpg/haploview (accessed on 30 May 2014).
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, Y.; Benjamini, Y. More powerful procedures for multiple significance testing. Stat. Med. 1990, 9, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Hair Jr, J.; Anderson, R.; Tatham, R.; Black, W. Multivariate Data Analysis. Prentice-Hall Inc.: New York City, NY, USA, 1998. [Google Scholar]
- Swanepoel, B. The Relevance of Specific C-reactive Protein Genetic Variants towards Cardiovascular Disease Risk in a Black South African Population Undergoing Epidemiological Transition. Master’s Thesis, North-West University, Potchefstroom, South Africa, September 2013. [Google Scholar]
- Roche, H.M.; Mensink, R.; Gibney, M.; Macdonald, I. Molecular Aspects of Nutrition; Blackwell Science: Oxford, UK, 2003. [Google Scholar]
- Aronson, D.; Bartha, P.; Zinder, O.; Kerner, A.; Markiewicz, W.; Avizohar, O.; Brook, G.; Levy, Y. Obesity is the major determinant of elevated C-reactive protein in subjects with the metabolic syndrome. Int. J. Obes. 2004, 28, 674–679. [Google Scholar] [CrossRef]
- Lemieux, I.; Pascot, A.; Prud’homme, D.; Alméras, N.; Bogaty, P.; Nadeau, A.; Bergeron, J.; Després, J.-P. Elevated C-reactive protein another component of the atherothrombotic profile of abdominal obesity. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Visser, M.; Bouter, L.M.; McQuillan, G.M.; Wener, M.H.; Harris, T.B. Elevated C-reactive protein levels in overweight and obese adults. J. Am. Med. Assoc. 1999, 282, 2131–2135. [Google Scholar] [CrossRef]
- Dod, H.S.; Bhardwaj, R.; Sajja, V.; Weidner, G.; Hobbs, G.R.; Konat, G.W.; Manivannan, S.; Gharib, W.; Warden, B.E.; Nanda, N.C. Effect of intensive lifestyle changes on endothelial function and on inflammatory markers of atherosclerosis. Am. J. Cardiol. 2010, 105, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Jialal, I.; Devaraj, S.; Venugopal, S.K. C-reactive protein: Risk marker or mediator in atherothrombosis? Hypertension 2004, 44, 6–11. [Google Scholar] [CrossRef]
- Ladeia, A.M.; Stefanelli, E.; Ladeia-Frota, C.; Moreira, A.; Hiltner, A.; Adan, L. Association between elevated serum C-reactive protein and triglyceride levels in young subjects with type 1 diabetes. Diabetes Care 2006, 29, 424–426. [Google Scholar] [CrossRef] [PubMed]
- Guha, M.; Bai, W.; Nadler, J.L.; Natarajan, R. Molecular mechanisms of tumor necrosis factor α gene expression in monocytic cells via hyperglycemia-induced oxidant stress-dependent and-independent pathways. J. Biol. Chem. 2000, 275, 17728–17739. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Rajala, M.W.; Berger, J.P.; Moller, D.E.; Barzilai, N.; Scherer, P.E. Hyperglycemia-induced production of acute phase reactants in adipose tissue. J. Biol. Chem. 2001, 276, 42077–42083. [Google Scholar] [CrossRef] [PubMed]
- Aronson, D.; Bartha, P.; Zinder, O.; Kerner, A.; Shitman, E.; Markiewicz, W.; Brook, G.; Levy, Y. Association between fasting glucose and C‐reactive protein in middle‐aged subjects. Diabet. Med. 2004, 21, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Mendall, M.; Patel, P.; Ballam, L.; Strachan, D.; Northfield, T. C-reactive protein and its relation to cardiovascular risk factors: A population based cross sectional study. Br. Med. J. 1996, 312, 1061–1065. [Google Scholar] [CrossRef]
- Yudkin, J.S.; Kumari, M.; Humphries, S.E.; Mohamed-Ali, V. Inflammation, obesity, stress and coronary heart disease: Is interleukin-6 the link? Atherosclerosis 2000, 148, 209–214. [Google Scholar] [CrossRef]
- Ferrucci, L.; Cherubini, A.; Bandinelli, S.; Bartali, B.; Corsi, A.; Lauretani, F.; Martin, A.; Andres-Lacueva, C.; Senin, U.; Guralnik, J.M. Relationship of plasma polyunsaturated fatty acids to circulating inflammatory markers. J. Clin. Endocrinol. Metab. 2006, 91, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Rallidis, L.S.; Paschos, G.; Liakos, G.K.; Velissaridou, A.H.; Anastasiadis, G.; Zampelas, A. Dietary α-linolenic acid decreases C-reactive protein, serum amyloid A and interleukin-6 in dyslipidaemic patients. Atherosclerosis 2003, 167, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Tannock, L.R.; O’Brien, K.D.; Knopp, R.H.; Retzlaff, B.; Fish, B.; Wener, M.H.; Kahn, S.E.; Chait, A. Cholesterol feeding increases C-reactive protein and serum amyloid A levels in lean insulin-sensitive subjects. Circulation 2005, 111, 3058–3062. [Google Scholar] [CrossRef] [PubMed]
- Pirro, M.; Schillaci, G.; Savarese, G.; Gemelli, F.; Mannarino, M.R.; Siepi, D.; Bagaglia, F.; Mannarino, E. Attenuation of inflammation with short-term dietary intervention is associated with a reduction of arterial stiffness in subjects with hypercholesterolaemia. Eur. J. Cardiovasc. Prev. Rehabil. 2004, 11, 497–502. [Google Scholar] [PubMed]
- Ridker, P.M.; Cannon, C.P.; Morrow, D.; Rifai, N.; Rose, L.M.; McCabe, C.H.; Pfeffer, M.A.; Braunwald, E. C-reactive protein levels and outcomes after statin therapy. N. Engl. J. Med. 2005, 352, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Momiyama, Y.; Ohmori, R.; Fayad, Z.A.; Kihara, T.; Tanaka, N.; Kato, R.; Taniguchi, H.; Nagata, M.; Nakamura, H.; Ohsuzu, F. Associations between plasma C-reactive protein levels and the severities of coronary and aortic atherosclerosis. J. Atheroscler. Thromb. 2010, 17, 460–467. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nienaber-Rousseau, C.; Swanepoel, B.; Dolman, R.C.; Pieters, M.; Conradie, K.R.; Towers, G.W. Interactions between C-Reactive Protein Genotypes with Markers of Nutritional Status in Relation to Inflammation. Nutrients 2014, 6, 5034-5050. https://doi.org/10.3390/nu6115034
Nienaber-Rousseau C, Swanepoel B, Dolman RC, Pieters M, Conradie KR, Towers GW. Interactions between C-Reactive Protein Genotypes with Markers of Nutritional Status in Relation to Inflammation. Nutrients. 2014; 6(11):5034-5050. https://doi.org/10.3390/nu6115034
Chicago/Turabian StyleNienaber-Rousseau, Cornelie, Bianca Swanepoel, Robin C. Dolman, Marlien Pieters, Karin R. Conradie, and G. Wayne Towers. 2014. "Interactions between C-Reactive Protein Genotypes with Markers of Nutritional Status in Relation to Inflammation" Nutrients 6, no. 11: 5034-5050. https://doi.org/10.3390/nu6115034
APA StyleNienaber-Rousseau, C., Swanepoel, B., Dolman, R. C., Pieters, M., Conradie, K. R., & Towers, G. W. (2014). Interactions between C-Reactive Protein Genotypes with Markers of Nutritional Status in Relation to Inflammation. Nutrients, 6(11), 5034-5050. https://doi.org/10.3390/nu6115034