Impact of Diet Composition in Adult Offspring is Dependent on Maternal Diet during Pregnancy and Lactation in Rats



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
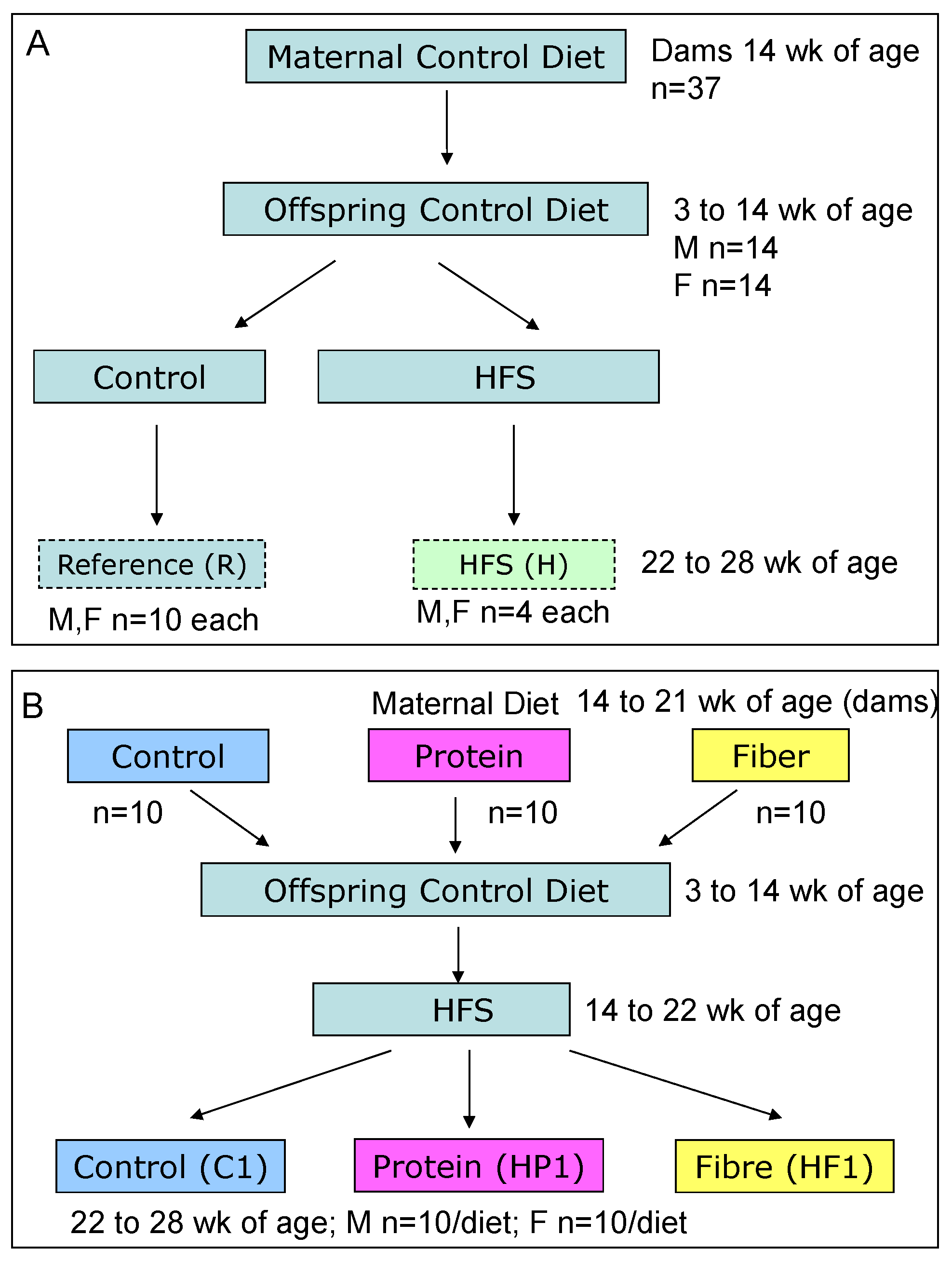
2.2. Animals and Diets


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g/kg) | Control (AIN-93M) | High Protein | High Fiber | HFS |
|---|---|---|---|---|
| Cornstarch | 465.7 | 205.7 | 325.5 | 49.5 |
| Casein | 140.0 | 400.0 | 120.4 | 140.0 |
| Dyetrose | 155.0 | 155.0 | 133.0 | - |
| Sucrose | 100.0 | 100.0 | 86.0 | 510.0 |
| Soybean Oil | 40.0 | 40.0 | 34.4 | 100.0 |
| Lard | - | - | - | 100.0 |
| Alphacel | 50.0 | 50.0 | 43.0 | 50.0 |
| AIN-93M Mineral Mix | 35.0 | 35.0 | 30.1 | 35.0 |
| AIN-93-VX Vitamin Mix | 10.0 | 10.0 | 8.6 | 10.0 |
| l-Cystine | 1.8 | 1.8 | 1.6 | 2.0 |
| dl-Methionine | - | - | - | 3.0 |
| Choline Bitartrate | 2.5 | 2.5 | 2.2 | 2.5 |
| Inulin | - | - | 107.6 | - |
| Oligofructose | - | - | 107.6 | - |
2.3. Oral Glucose Tolerance Test and Tissue Sampling
2.4. Plasma Analysis
2.5. Hepatic Triglyceride Analysis
2.6. RNA Extraction and Real-Time PCR
2.7. Statistical Analysis
3. Results
3.1. Offspring Re-Matched to Maternal HP, HF or C
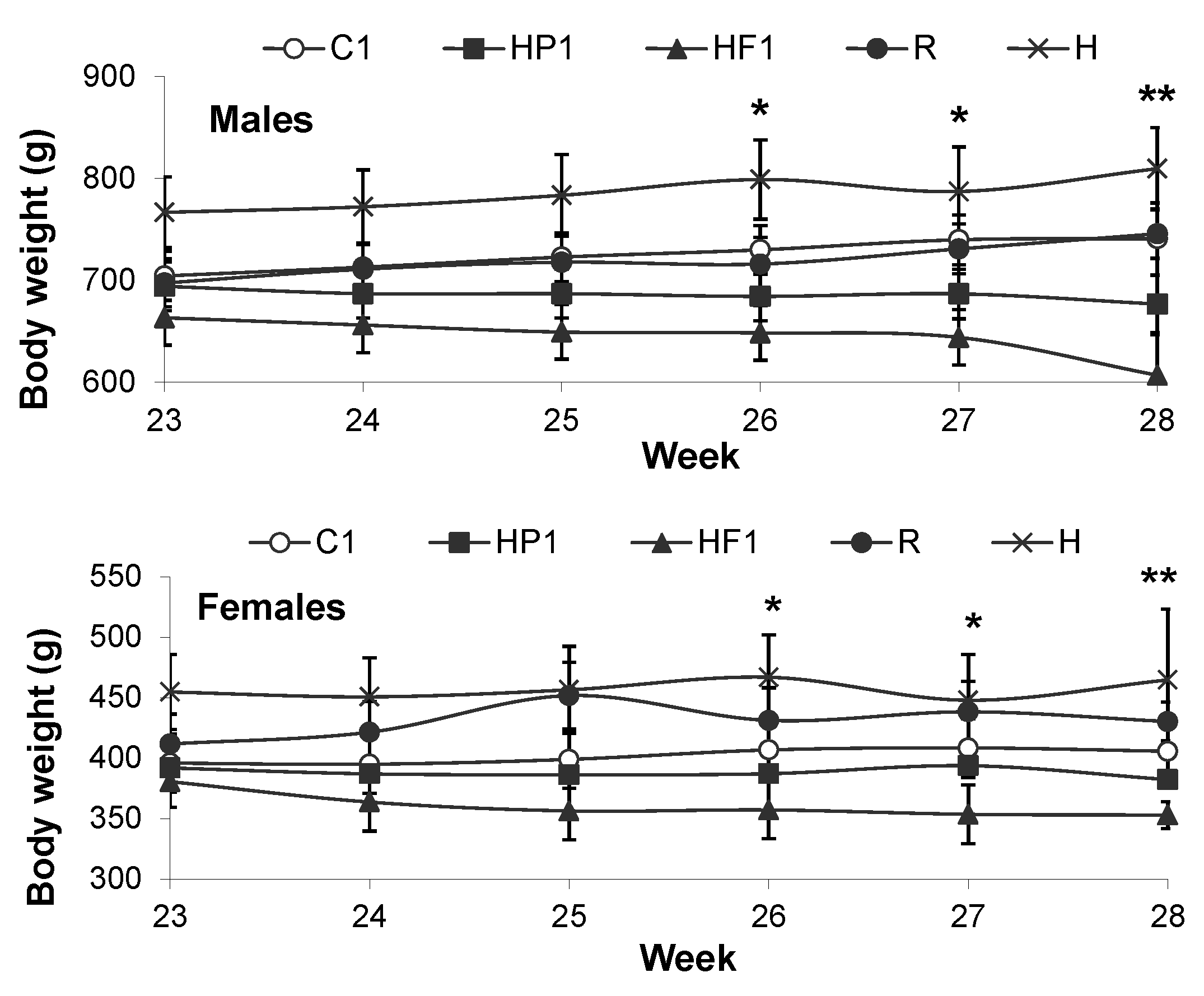
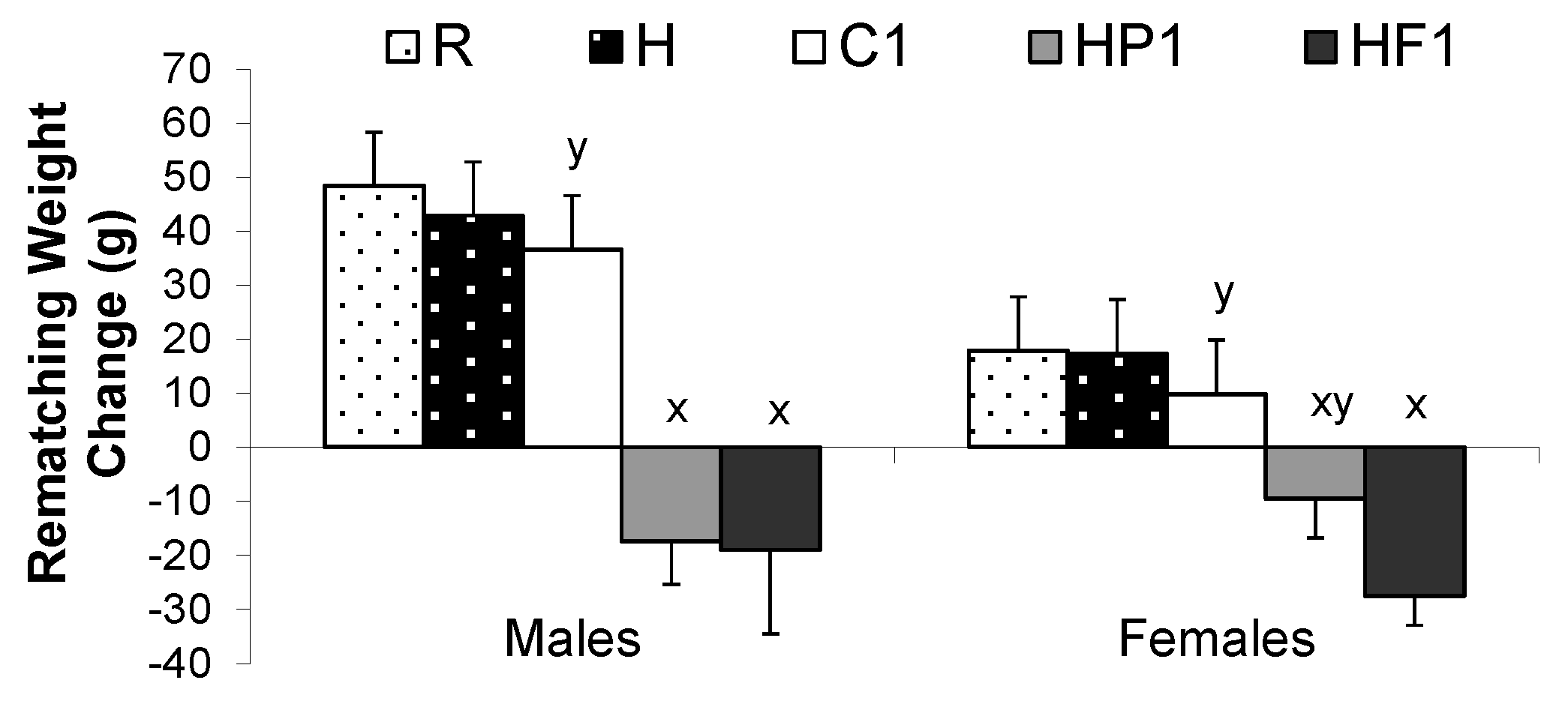
3.1.1. Growth and Energy Intake



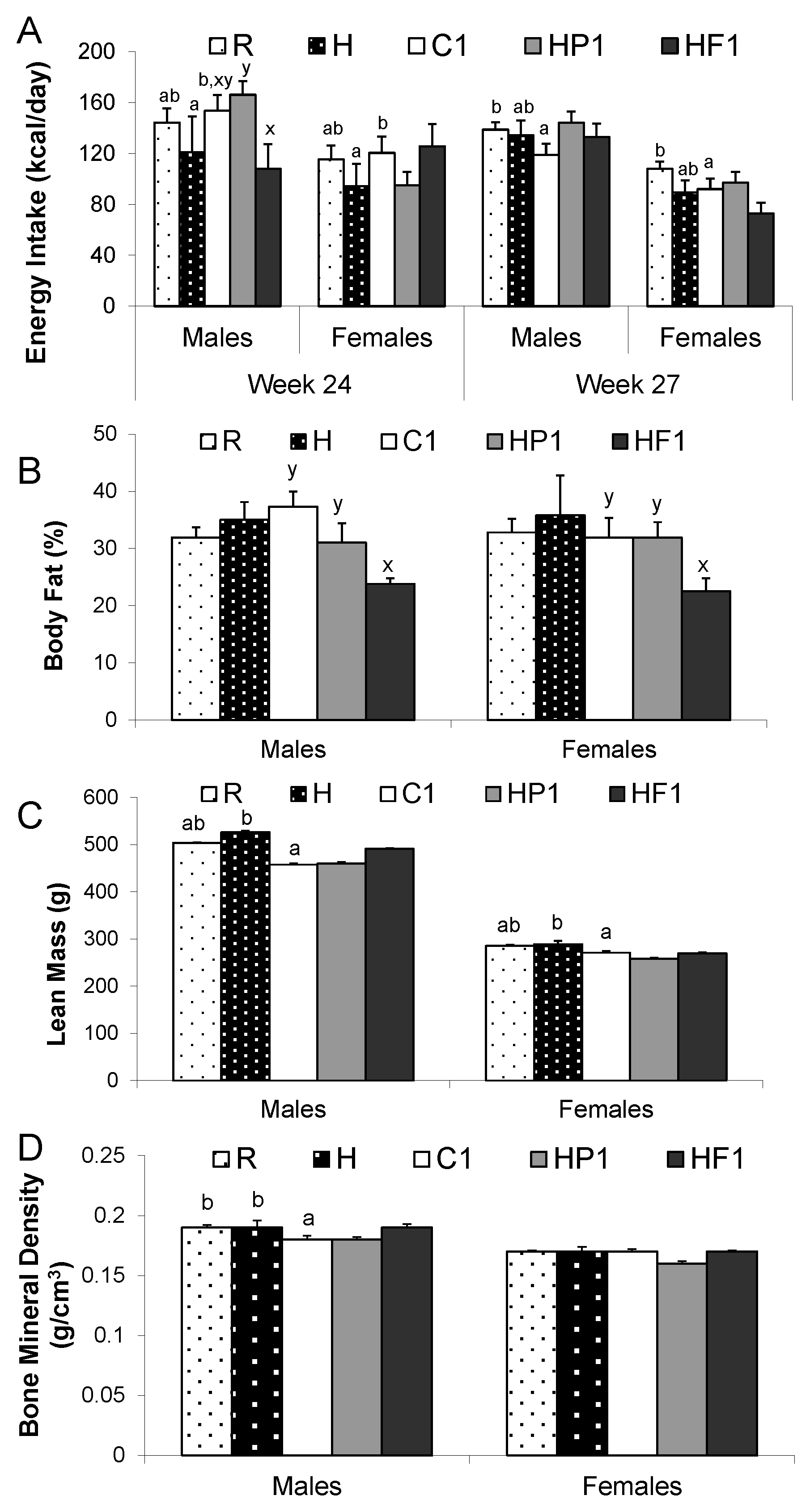
3.1.2. Body Composition
| Diet Group | ||||||
|---|---|---|---|---|---|---|
| Sex | HP1 | HF1 | C1 | R | H | |
| Liver (g) | M | 16.1 ± 0.5 | 15.5 ± 0.7 | 17.3 ± 0.7 | 18.8 ± 0.9 | 19.0 ± 1.0 |
| F | 9.5 ± 0.6 | 8.9 ± 0.4 | 9.8 ± 0.8 | 9.0 ± 1.2 | 10.2 ± 1.1 | |
| Stomach (g) | M | 2.7 ± 0.1 | 2.6 ± 0.1 | 2.8 ± 0.1 a | 3.3 ± 0.1 b | 3.1 ± 0.2 b |
| F | 2.0 ± 0.1 | 2.0 ± 0.1 | 1.9 ± 0.1 a | 1.9 ± 0.2 b | 2.4 ± 0.3 b | |
| Small Intestine Length (cm) | M | 130.5 ± 1.5 x | 143.8 ± 2.7 y | 125.7 ± 2.4 x,a | 134.8 ± 2.2 b | 133.8 ± 1.3 a,b |
| F | 115.0 ± 3.4 x | 122.8 ± 2.5 y | 116.8 ± 3.7 x,a | 104.6 ± 2.2 b | 122 ± 5.5 a,b | |
| Small Intestine Weight (g) | M | 8.1 ± 0.4 | 8.7 ± 0.4 | 7.9 ± 0.2 | 8.2 ± 0.24 | 8.6 ± 0.13 |
| F | 6.1 ± 0.4 | 6.1 ± 0.4 | 6.8 ± 0.3 | 5.4 ± 0.64 | 6.3 ± 0.41 | |
| Cecum (g) | M | 1.07 ± 0.1 x | 3.2 ± 0.3 y | 1.01 ± 0.1 x,a | 1.5 ± 0.04 b | 0.63 ± 0.36 a |
| F | 0.74 ± 0.06 x | 2.3 ± 0.3 y | 0.8 ± 0.1 x,a | 1.0 ± 0.19 b | 0.88 ± 0.29 a | |
| Colon Length (cm) | M | 23.5 ± 0.7 x | 26.1 ± 0.9 y | 21.6 ± 0.7 x | 21.1 ± 0.43 | 23.3 ± 0.85 |
| F | 19.6 ± 0.3 x | 24.8 ± 0.4 y | 20.0 ± 0.7 x | 16.4 ± 1.9 | 20.5 ± 1.3 | |
| Colon Weight (g) | M | 2.1 ± 0.1 y | 2.4 ± 0.2 z | 1.8 ± 0.1 x | 1.6 ± 0.05 | 1.9 ± 0.1 |
| F | 1.5 ± 0.1 y | 2.0 ± 0.1 z | 1.3 ± 0.05 x | 1.1 ± 0.13 | 1.3 ± 0.06 | |
| Kidneys (g) | M | 4.0 ± 0.1 y | 3.4 ± 0.1 x | 3.6 ± 0.1 x | 3.5 ± 0.1 | 3.8 ± 0.2 |
| F | 2.4 ± 0.1 y | 2.1 ± 0.1 x | 2.3 ± 0.1 x | 1.9 ± 0.2 | 2.4 ± 0.3 | |
| Nasoanal Length (cm) | M | 28.9 ± 0.16 | 28.6 ± 0.34 | 28.5 ± 0.19 | 28.4 ± 0.16 | 28.8 ± 0.32 |
| F | 24.0 ± 0.41 | 23.5 ± 0.19 | 24.0 ± 0.17 | 23.6 ± 0.27 | 23.8 ± 0.32 | |
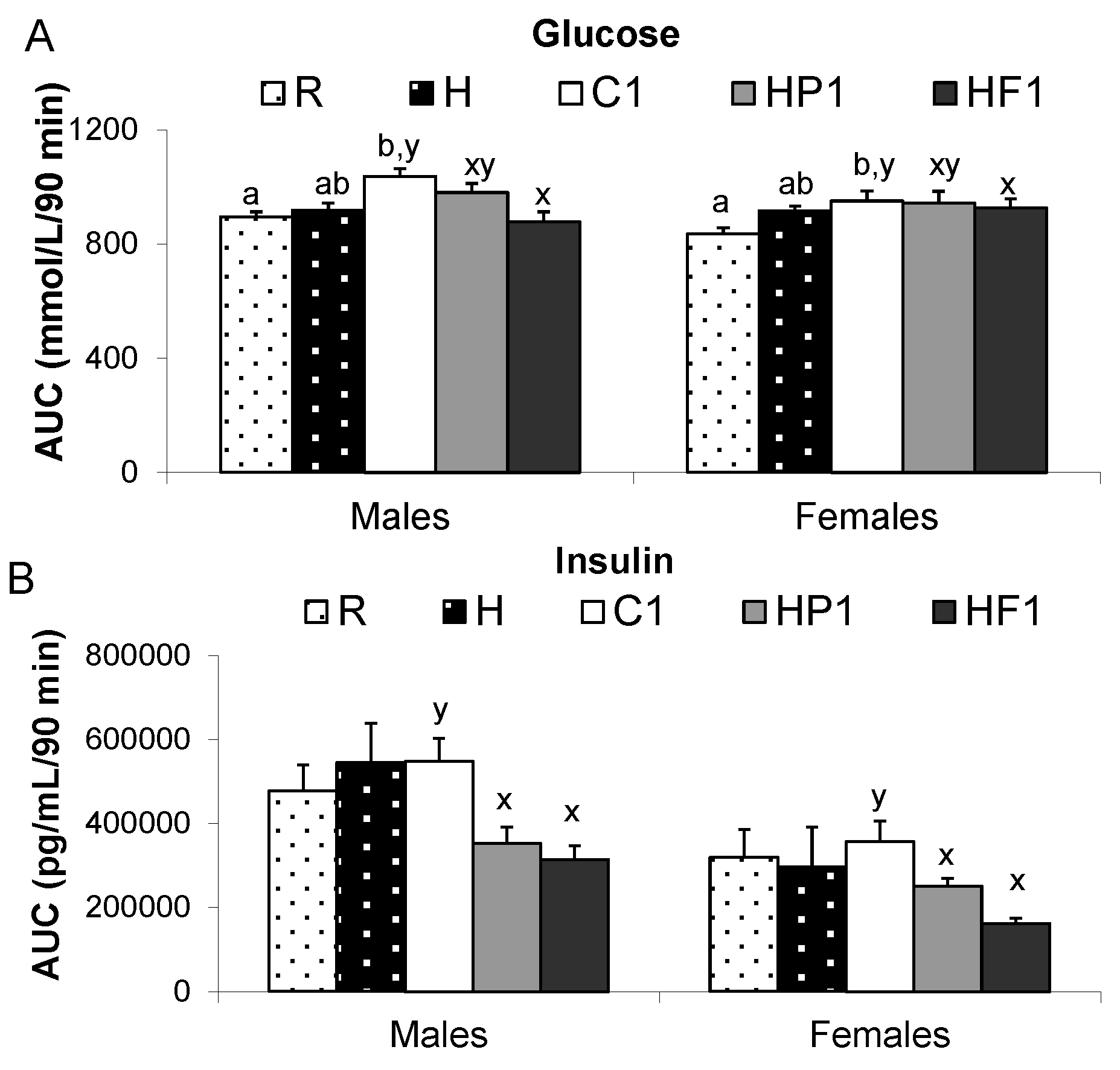
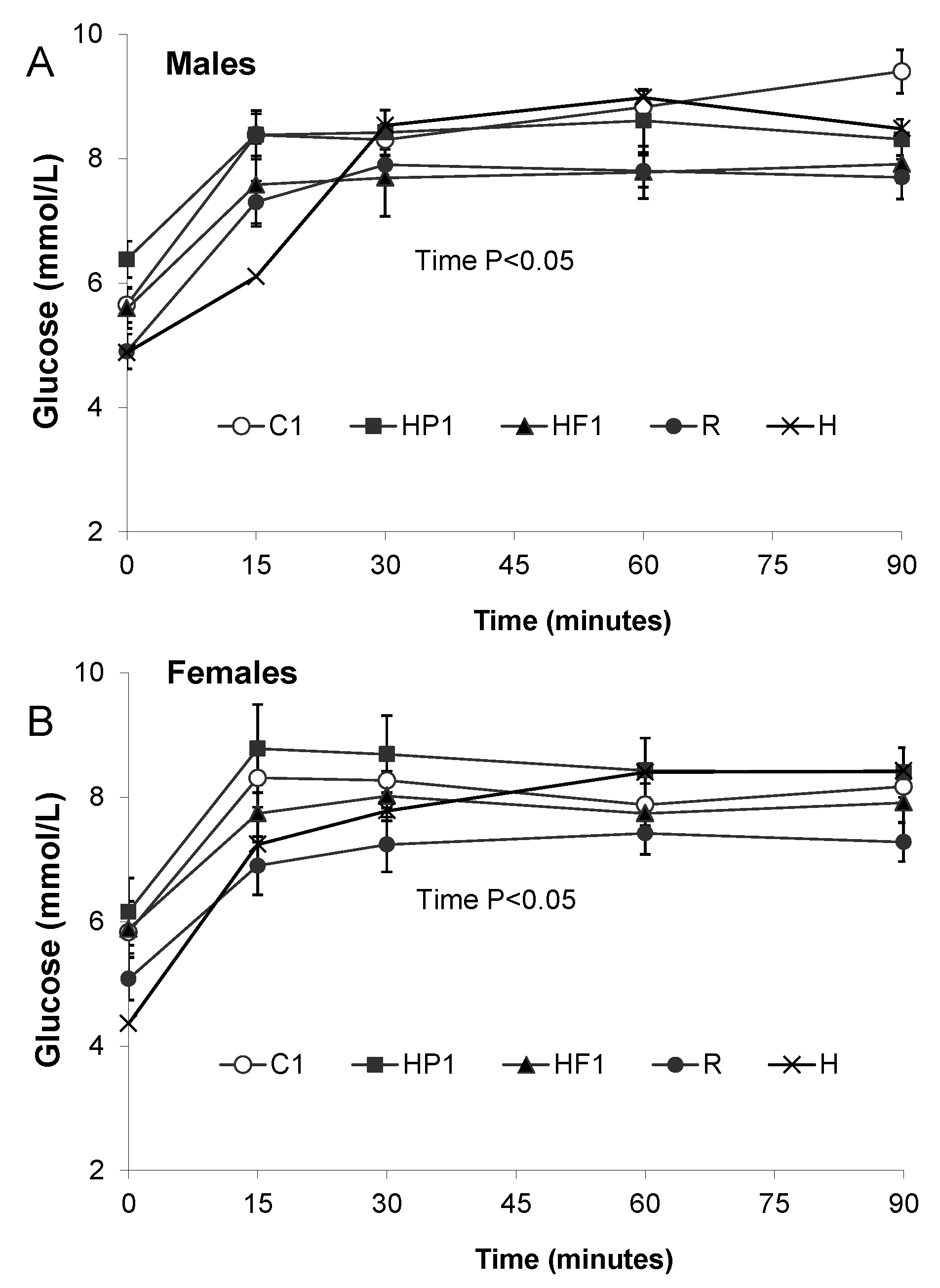
3.1.3. Plasma Satiety Hormones and Blood Glucose
| Diet Group | ||||||
|---|---|---|---|---|---|---|
| Sex | HP1 | HF1 | C1 | R | H | |
| Glucose (mmol/l) | M | 6.38 ± 0.29 | 5.59 ± 0.32 | 5.65 ± 0.28 b | 4.98 ± 0.08 a | 4.88 ± 0.45 a |
| F | 6.16 ± 0.54 | 5.88 ± 0.45 | 5.83 ± 0.34 b | 3.20 ± 0.67 a | 4.55 ± 0.17 a | |
| Insulin (pg/mL) | M | 2565 ± 443 x | 3312 ± 633 x | 5958 ± 731 y,b | 3022 ± 595 a | 1888 ± 150 a |
| F | 1880 ± 274 | 1321 ± 343 | 1692 ± 513 a | 1599 ± 394 a,b | 2375 ± 1133 a,b | |
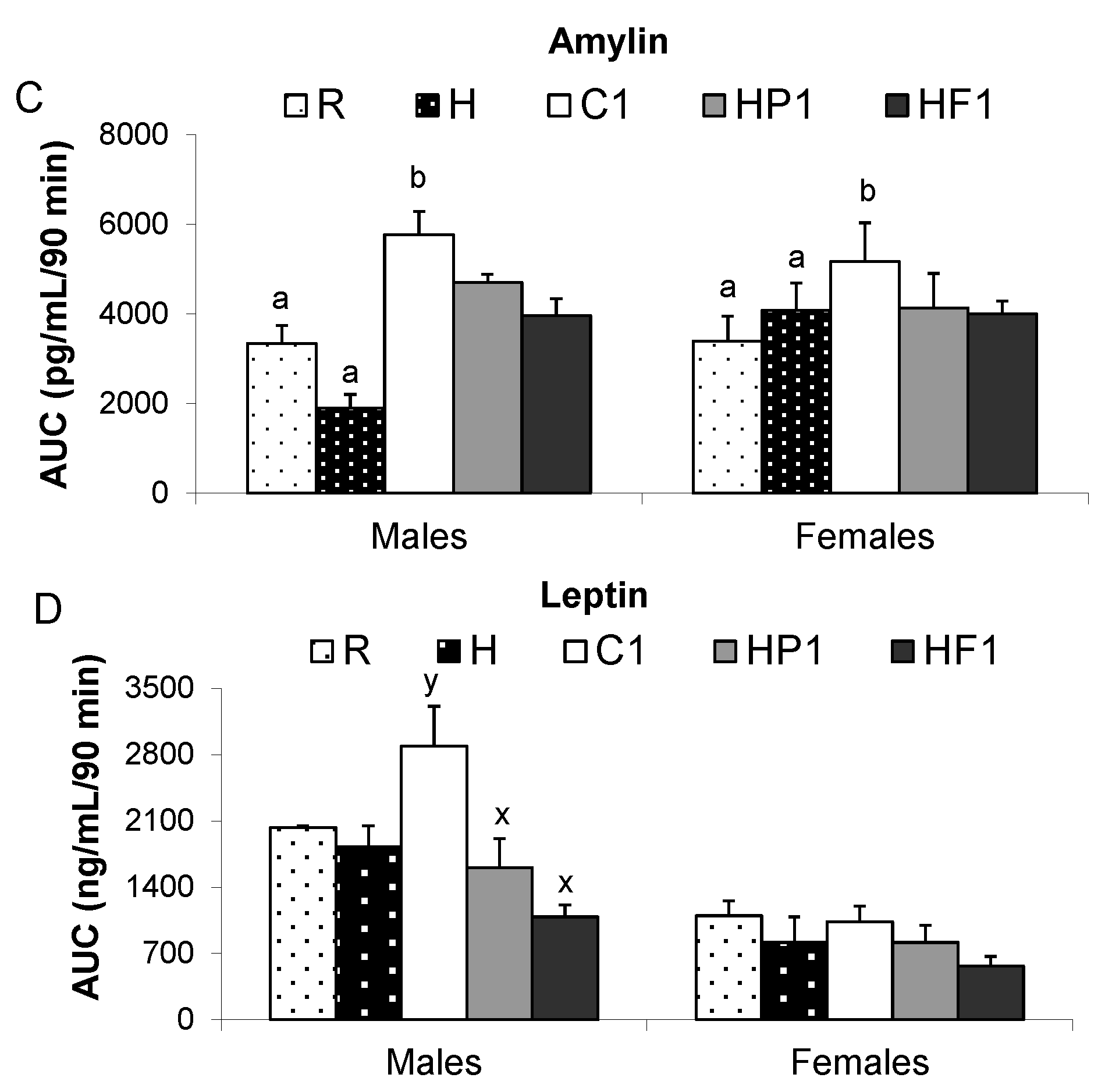
| Amylin (pg/mL) | M | 56.4 ± 10.5 | 63.0 ± 5.7 | 75.0 ± 11.1 a | 43.1 ± 6.8 a | 418.6 ± 145.2 b |
| F | 52.8 ± 9.4 | 54.3 ± 4.8 | 52.2 ± 8.1 a,b | 37.3 ± 6.4 a,b | 51.5 ± 9.7 a | |
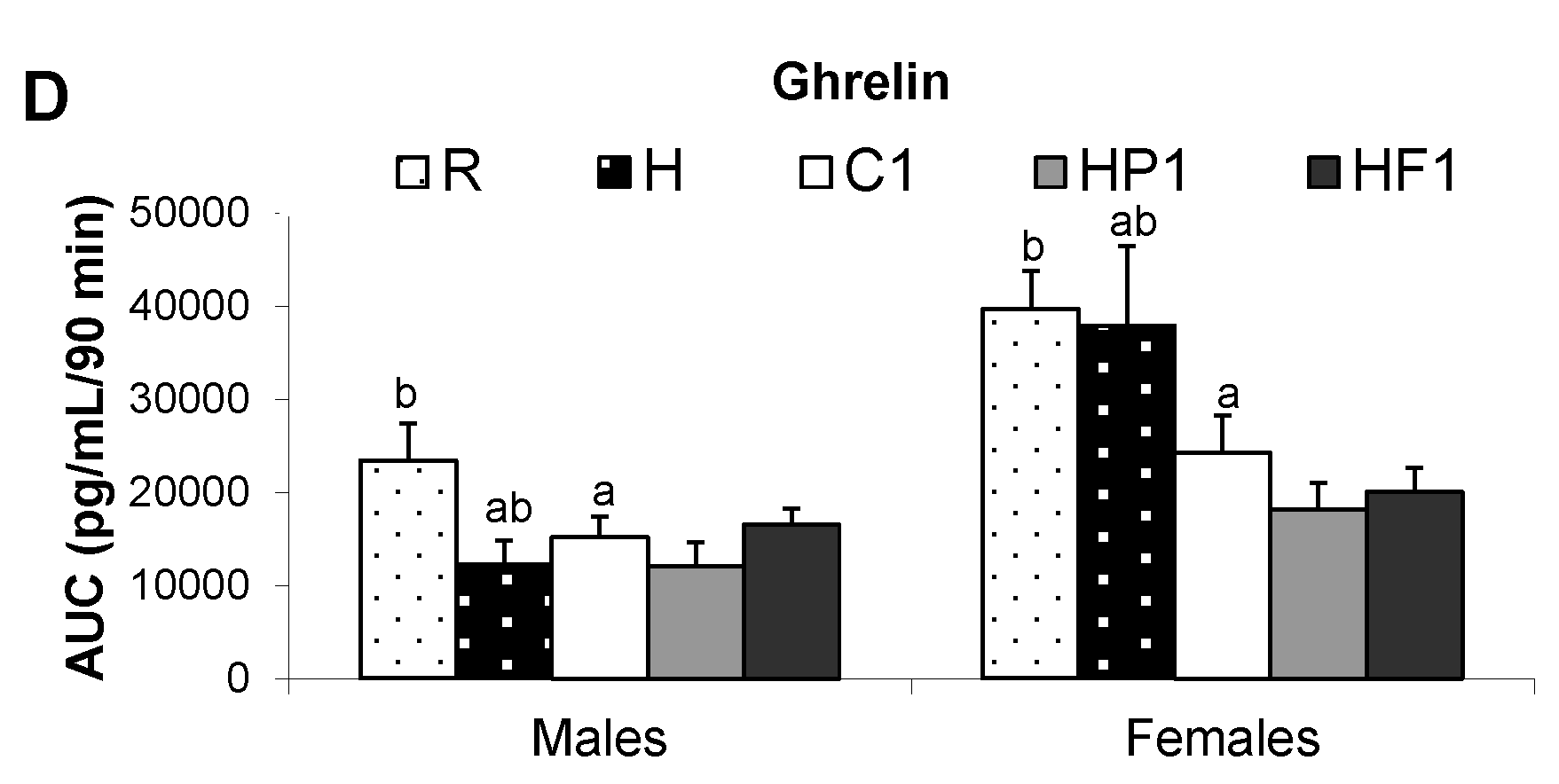
| Ghrelin (ng/mL) | M | 159.4 ± 36.8 | 235.1 ± 39.2 | 233.1 ± 39.7 | 266.8 ± 52.1 | 171.4 ± 32.1 |
| F | 246.1 ± 50.7 | 369.6 ± 47.7 | 344.7 ± 55.4 | 529.1 ± 74.4 | 402.7 ± 65.7 | |
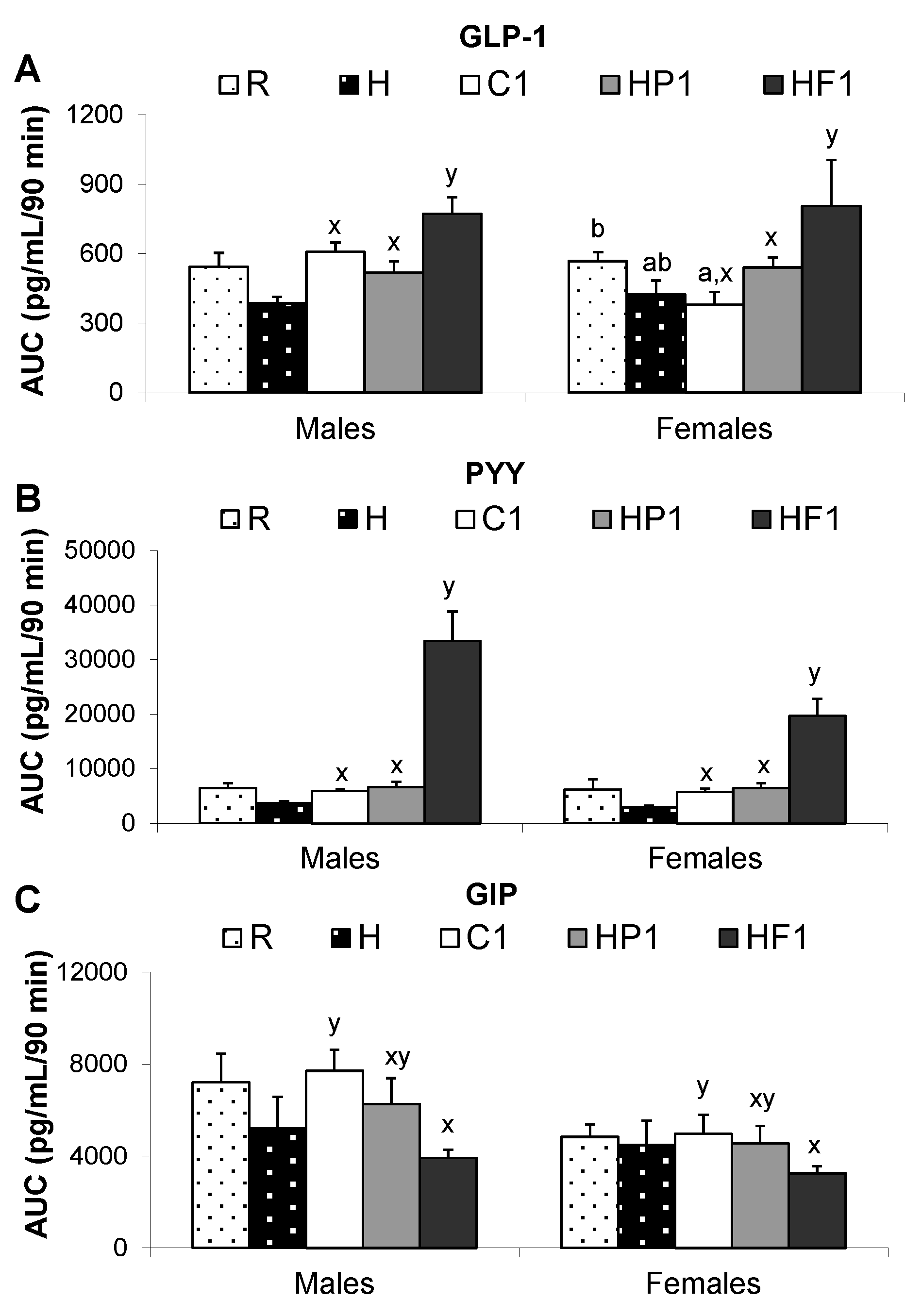
| GIP (ng/mL) | M | 46.5 ± 8.3 | 47.8 ± 6.6 | 49.9 ± 6.3 | 49.9 ± 6.3 | 87.3 ± 14.0 |
| F | 33.4 ± 9.2 | 42.1 ± 3.2 | 41.6 ± 8.4 | 49.7 ± 5.3 | 34.9 ± 6.1 | |
| PYY (pg/mL) | M | 62.7 ± 7.5 x | 262.5 ± 43.2 y | 64.1 ± 4.4 x | 72.4 ± 6.3 | 54.0 ± 10.9 |
| F | 63.5 ± 8.9 x | 201.3 ± 35.1 y | 55.8 ± 5.0 x | 62.2 ± 13.6 | 36.1 ± 4.9 | |
| GLP-1 (pg/mL) | M | 6.0 ± 0.58 x | 8.4 ± 0.79 y | 7.22 ± 0.48 x,a,b | 6.5 ± 0.42 a,b | 5.4 ± 0.19 a,b |
| F | 6.36 ± 0.37 x | 8.54 ± 1.5 y | 4.45 ± 0.72 x,a | 6.4 ± 0.37 b | 4.82 ± 0.35 a,b | |
| Leptin (ng/mL) | M | 21.0 ± 4.2 y | 14.5 ± 2.2 x | 27.2 ± 2.5 y | 20.4 ± 1.9 | 18.4 ± 2.4 |
| F | 10.3 ± 1.6 y | 6.5 ± 1.0 x | 11.1 ± 2.1 y | 12.9 ± 1.4 | 11.3 ± 4.1 | |
| HOMA-IR | M | 20.2 ± 3.8 x | 19.0 ± 3.4 x | 35.9 ± 4.8 y,b | 15.3 ± 2.9 a | 9.8 ± 1.1 a |
| F | 10.4 ± 1.6 z | 5.8 ± 1.1 z | 7.1 ± 1.5 z,d | 8.8 ± 2.2 a,b,c,d | 35.5 ± 4.5 c | |
| Liver TG (mg/mg protein) | M | 36.6 ± 3.1 y | 30.8 ± 3.1 x | 43.1 ± 3.7 y | 33.1 ± 2.5 | 30.4 ± 4.0 |
| F | 26.8 ± 2.5 y | 23.2 ± 1.5 x | 32.2 ± 1.6 y | 36.8 ± 2.0 | 35.5 ± 4.5 | |





3.1.4. Liver Triglyceride
3.1.5. Hepatic Gene Expression
| Diet Group | p (2-Way ANOVA) | ||||||
|---|---|---|---|---|---|---|---|
| Sex | HP1 | HF1 | C1 | Diet | Sex | Diet × Sex | |
| SREBP-1c | M | 5.5 ± 0.73 | 2.6 ± 0.7 | 4.6 ± 0.67 | 0.002 | 0.002 | 0.143 |
| F | 4.0 ± 0.66 | 2.1 ± 0.47 | 1.7 ± 0.53 | ||||
| ACC | M | 13.3 ± 2.0 | 10.9 ± 2.0 | 15.0 ± 1.6 | 0.032 | 0.553 | 0.097 |
| F | 18.9 ± 3.1 | 10.6 ± 1.2 | 12.6 ± 1.3 | ||||
| FAS | M | 6.8 ± 2.0 | 4.7 ± 1.1 | 4.0 ± 0.74 | 0.002 | 0.001 | 0.124 |
| F | 18.2 ± 3.1 | 13.0 ± 1.9 | 8.2 ± 0.92 | ||||
| PGC1α | M | 88.9 ± 19.6 y | 45.9 ± 9.8 x | 24.1 ± 5.8 x | 0.001 | 0.064 | 0.972 |
| F | 110.9 ± 21.1 y | 62.5 ± 13.6 x | 45.7 ± 6.4 x | ||||
| Glucose-6-Phosphatase | M | 23.6 ± 5.5 x | 19.5 ± 3.8 x,y | 17.1 ± 2.7 x,y | 0.001 | 0.031 | 0.014 |
| F | 43.6 ± 4.4 y | 16.7 ± 2.9 x | 20.5 ± 2.3 x | ||||
| AMPKα1 | M | 16.5 ± 1.6 | 11.7 ± .9 | 10.7 ± 1.4 | 0.006 | 0.587 | 0.121 |
| F | 16.2 ± 2.8 | 9.7 ± 1.8 | 15.2 ± 1.0 | ||||
3.2. Effect of HFS Exposure and Reversibility
3.2.1. Growth and Energy Intake
3.2.2. Body Composition
3.2.3. Plasma Satiety Hormones and Blood Glucose
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| HFS | High fat/high sucrose |
| HF | High prebiotic fiber diet |
| HP | High protein diet |
| C | Control diet |
| R | Reference group that consumed control diet throughout |
| C1 | Rats switched back to control diet after high fat/sucrose diet |
| HF1 | Rats switched back to high fiber diet after high fat/sucrose diet |
| HP1 | Rats switched back to high protein diet after high fat/sucrose diet |
| AIN-93 | American Institute of Nutrition-93 |
| GLP-1 | Glucagon-like peptide 1 |
| PYY | Peptide tyrosine tyrosine |
| GIP | Glucose-dependent insulinotropic polypeptide |
| OGTT | Oral glucose tolerance test |
| DXA | Dual-energy x-ray absorptiometry |
| HOMA-IR | Homeostasis model assessment of insulin resistance |
| ACC | Acetyl Co-A carboxylase |
| FAS | Fatty acid synthase |
| SREBP-1c | Sterol regulatory element binding protein-1c |
| AMPKα1 | Adenosine monophosphate-activated protein kinase alpha-1 |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| CT | Cycle threshold |
| AUC | Area under the curve |
References
- Lucas, A. Programming by Early Nutrition: An Experimental Approach. J. Nutr. 1998, 128, 410S–406S. [Google Scholar]
- Ross, M.G.; Desai, M. Developmental Programming of Offspring Obesity, Adipogenesis, and Appetite. Clin. Obstet. Gynecol. 2013, 56, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Dekou, V.; Hanson, M.; Poston, L.; Taylor, P. Predictive Adaptive Responses to Maternal High-Fat Diet Prevent Endothelial Dysfunction but Not Hypertension in Adult Rat Offspring. Circulation 2004, 110, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Lecourt, E.; Dewulf, E.M.; Sohet, F.M.; Pachikian, B.D.; Naslain, D.; de Backer, F.; Neyrinck, A.M.; Delzenne, N.M. Gut Microbiota Fermentation of Prebiotics Increases Satietogenic and Incretin Gut Peptide Production With Consequences for Appetite Sensation and Glucose Response After a Meal. Am. J. Clin. Nutr. 2009, 90, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Daubioul, C.; Rousseau, N.; Demeure, R.; Gallez, B.; Taper, H.S.; Declerck, B.; Delzenne, N.M. Dietary Fructans, but Not Cellulose, Decrease Triglyceride Accumulation in the Liver of Obese Zucker Fa/Fa Rats. J. Nutr. 2002, 132, 967–973. [Google Scholar] [PubMed]
- Parnell, J.A.; Reimer, R.A. Effect of Prebiotic Fiber Supplementation on Hepatic Gene Expression and Serum Lipids: A Dose-Response Study in JCR: LA-Cp Rats. Br. J. Nutr. 2010, 103, 1577–1584. [Google Scholar] [CrossRef] [PubMed]
- Reimer, R.A.; Maurer, A.D.; Eller, L.K.; Hallam, M.C.; Shaykhutdinov, R.; Vogel, H.J.; Weljie, A.M. Satiety Hormone and Metabolomic Response to an Intermittent High Energy Diet Differs in Rats Consuming Long-Term Diets High in Protein or Prebiotic Fiber. J. Proteome Res. 2012, 11, 4065–4074. [Google Scholar] [CrossRef] [PubMed]
- Hallam, M.C.; Reimer, R.A. A Maternal High Protein Diet Predisposes Female Offspring to Increased Fat Mass in Adulthood Whereas a Prebiotic Fibre Diet Decreases Fat Mass in Rats. Br. J. Nutr. 2013, 110, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Maurer, A.D.; Eller, L.K.; Hallam, M.C.; Taylor, K.; Reimer, R.A. Consumption of Diets High in Prebiotic Fiber or Protein During Growth Influences the Response to a High Fat and Sucrose Diet in Adulthood in Rats. Nutr. Metab. 2010, 7, 77. [Google Scholar] [CrossRef] [PubMed]
- Moran, T.H.; Dailey, M.J. Intestinal Feedback Signaling and Satiety. Physiol. Behav. 2011, 105, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Hoffmann, G. Long-Term Effects of Low-Fat Diets Either Low or High in Protein on Cardiovascular and Metabolic Risk Factors: A Systematic Review and Meta-Analysis. Nutr. J. 2013, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Kellow, N.J.; Coughlan, M.T.; Reid, C.M. Metabolic Benefits of Dietary Prebiotics in Human Subjects: A Systematic Review of Randomised Controlled Trials. Br. J. Nutr. 2013, 111, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Janevski, M.; Ratnayake, S.; Siljanovski, S.; McGlynn, M.A.; Cameron-Smith, D.; Lewandowski, P. Fructose Containing Sugars Modulate MRNA of Lipogenic Genes ACC and FAS and Protein Levels of Transcription Factors ChREBP and SREBP1c with No Effect on Body Weight or Liver Fat. Food Funct. 2012, 3, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Sadi, G.; Ergin, V.; Yilmaz, G.; Pektas, M.B.; Yildirim, O.G.; Menevse, A.; Akar, F. High-Fructose Corn Syrup-Induced Hepatic Dysfunction in Rats: Improving Effect of Resveratrol. Eur. J. Nutr. 2014, 54, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Haas, J.T.; Miao, J.; Chanda, D.; Wang, Y.; Zhao, E.; Haas, M.E.; Hirschey, M.; Vaitheesvaran, B.; Farese, R.V., Jr.; Kurland, I.J.; et al. Hepatic Insulin Signaling Is Required for Obesity-Dependent Expression of SREBP-1c MRNA but Not for Feeding-Dependent Expression. Cell Metab. 2012, 15, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.L.; Yang, H.T.; Lee, Y.J.; Lin, C.C.; Chang, M.H.; Yin, M.C. Asiatic Acid Ameliorates Hepatic Lipid Accumulation and Insulin Resistance in Mice Consuming a High-Fat Diet. J. Agric. Food Chem. 2014, 62, 4625–4631. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.B.; Wang, Z.S.; Peng, Q.H.; Tan, C.; Zou, H.W. Effects of Different Levels of Protein Supplementary Diet on Gene Expressions Related to Intramuscular Deposition in Early-Weaned Yaks. Anim. Sci. J. 2014, 85, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zeng, T.; Wang, S.E.; Wang, W.; Wang, Q.; Yu, H.X. Fructo-Oligosaccharides Enhance the Mineral Absorption and Counteract the Adverse Effects of Phytic Acid in Mice. Nutrition 2010, 26, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Belobrajdic, D.P.; Jenkins, C.L.; Bushell, R.; Morell, M.K.; Bird, A.R. Fructan Extracts From Wheat Stem and Barley Grain Stimulate Large Bowel Fermentation in Rats. Nutr. Res. 2012, 32, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in Gut Microbiota Control Metabolic Endotoxemia-Induced Inflammation in High-Fat Diet-Induced Obesity and Diabetes in Mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Neyrinck, A.M.; Fava, F.; Knauf, C.; Burcelin, R.G.; Tuohy, K.M.; Gibson, G.R.; Delzenne, N.M. Selective Increases of Bifidobacteria in Gut Microflora Improve High-Fat-Diet-Induced Diabetes in Mice Through a Mechanism Associated With Endotoxaemia. Diabetologia 2007, 50, 2374–2383. [Google Scholar] [CrossRef] [PubMed]
- Hallam, M.C.; Barile, D.; Meyrand, M.; German, J.B.; Reimer, R.A. Maternal High-Protein or High-Prebiotic-Fiber Diets Affect Maternal Milk Composition and Gut Microbiota in Rat Dams and Their Offspring. Obesity 2014, 22, 2344–2351. [Google Scholar] [CrossRef] [PubMed]
- Bomhof, M.R.; Saha, D.C.; Reid, D.T.; Paul, H.A.; Reimer, R.A. Combined Effects of Oligofructose and Bifidobacterium Animalis on Gut Microbiota and Glycemia in Obese Rats. Obesity 2014, 22, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Buettner, R.; Scholmerich, J.; Bollheimer, L.C. High-Fat Diets: Modeling the Metabolic Disorders of Human Obesity in Rodents. Obesity 2007, 15, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Sugatani, J.; Osabe, M.; Wada, T.; Yamakawa, K.; Yamazaki, Y.; Takahashi, T.; Ikari, A.; Miwa, M. Comparison of Enzymatically Synthesized Inulin, Resistant Maltodextrin and Clofibrate Effects on Biomarkers of Metabolic Disease in Rats Fed a High-Fat and High-Sucrose (Cafeteria) Diet. Eur. J. Nutr. 2008, 47, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Baggio, L.L.; Drucker, D.J. Biology of Incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef] [PubMed]
- Duca, F.A.; Sakar, Y.; Covasa, M. Combination of Obesity and High-Fat Feeding Diminishes Sensitivity to GLP-1R Agonist Exendin-4. Diabetes 2013, 62, 2410–2415. [Google Scholar] [CrossRef] [PubMed]
- South, T.; Westbrook, F.; Morris, M.J. Neurological and Stress Related Effects of Shifting Obese Rats from a Palatable Diet to Chow and Lean Rats from Chow to a Palatable Diet. Physiol. Behav. 2012, 105, 1052–1057. [Google Scholar] [CrossRef] [PubMed]
- Malta, A.; de Oliveira, J.C.; Ribeiro, T.A.; Tofolo, L.P.; Barella, L.F.; Prates, K.V.; Miranda, R.A.; Elmhiri, G.; Franco, C.C.; Agostinho, A.R.; et al. Low-Protein Diet in Adult Male Rats Has Long-Term Effects on Metabolism. J. Endocrinol. 2014, 221, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Rolls, B.J.; Rowe, E.A.; Turner, R.C. Persistent Obesity in Rats Following a Period of Consumption of a Mixed, High Energy Diet. J. Physiol. 1980, 298, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Jou, W.; Gavrilova, O.; Hall, K.D. Persistent Diet-Induced Obesity in Male C57BL/6 Mice Resulting From Temporary Obesigenic Diets. PLoS ONE 2009, 4, e5370. [Google Scholar] [CrossRef] [PubMed]
- Parekh, P.I.; Petro, A.E.; Tiller, J.M.; Feinglos, M.N.; Surwit, R.S. Reversal of Diet-Induced Obesity and Diabetes in C57BL/6J Mice. Metabolism 1998, 47, 1089–1096. [Google Scholar] [CrossRef]
- Bartness, T.J.; Polk, D.R.; McGriff, W.R.; Youngstrom, T.G.; DiGirolamo, M. Reversal of High-Fat Diet-Induced Obesity in Female Rats. Am. J. Physiol. 1992, 263, R790–R797. [Google Scholar] [PubMed]
- Sako, N.; Okamoto, K.; Mori, T.; Yamamoto, T. The Hardness of Food Plays an Important Role in Food Selection Behavior in Rats. Behav. Brain Res. 2002, 133, 377–382. [Google Scholar] [CrossRef]
- Levin, B.E.; Dunn-Meynell, A.A. Defense of Body Weight Depends on Dietary Composition and Palatability in Rats With Diet-Induced Obesity. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 282, R46–R54. [Google Scholar] [PubMed]
- Wang, W.C.; Worsley, A.; Hodgson, V. Classification of Main Meal Patterns—A Latent Class Approach. Br. J. Nutr. 2013, 109, 2285–2296. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.W.; Zaghloul, S.; Ali, H.; Harrison, G.; Yeatts, K.; el Sadiq, M.; Popkin, B.M. Nutrition Transition in the United Arab Emirates. Eur. J. Clin. Nutr. 2011, 65, 1326–1337. [Google Scholar] [CrossRef] [PubMed]
- Neustaedter, M.; Reimer, R.A.; Faculty of Kinesiology, University of Calgary, Calgary, AB, Canada. Unpublished work; 2015.
- Gluckman, P.D.; Hanson, M.A. The Developmental Origins of the Metabolic Syndrome. Trends Endocrinol. Metab. 2004, 15, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Maurer, A.D.; Chen, Q.; McPherson, C.; Reimer, R.A. Changes in Satiety Hormones and Expression of Genes Involved in Glucose and Lipid Metabolism in Rats Weaned Onto Diets High in Fiber or Protein Reflect Susceptibility to Increased Fat Mass in Adulthood. J. Physiol. 2009, 587, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [PubMed]
- Pyra, K.A.; Saha, D.C.; Reimer, R.A. Prebiotic Fiber Increases Hepatic Acetyl CoA Carboxylase Phosphorylation and Suppresses Glucose-Dependent Insulinotropic Polypeptide Secretion More Effectively When Used With Metformin in Obese Rats. J. Nutr. 2012, 142, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of Housekeeping Genes for Gene Expression Studies in Human Reticulocytes Using Real-Time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hallam, M.C.; Reimer, R.A. Impact of Diet Composition in Adult Offspring is Dependent on Maternal Diet during Pregnancy and Lactation in Rats. Nutrients 2016, 8, 46. https://doi.org/10.3390/nu8010046
Hallam MC, Reimer RA. Impact of Diet Composition in Adult Offspring is Dependent on Maternal Diet during Pregnancy and Lactation in Rats. Nutrients. 2016; 8(1):46. https://doi.org/10.3390/nu8010046
Chicago/Turabian StyleHallam, Megan C., and Raylene A. Reimer. 2016. "Impact of Diet Composition in Adult Offspring is Dependent on Maternal Diet during Pregnancy and Lactation in Rats" Nutrients 8, no. 1: 46. https://doi.org/10.3390/nu8010046
APA StyleHallam, M. C., & Reimer, R. A. (2016). Impact of Diet Composition in Adult Offspring is Dependent on Maternal Diet during Pregnancy and Lactation in Rats. Nutrients, 8(1), 46. https://doi.org/10.3390/nu8010046