Intakes of Folate and Vitamin B12 and Biomarkers of Status in the Very Old: The Newcastle 85+ Study

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Newcastle 85+ Study
2.2. Dietary Assessment and Food Groups
2.3. Nutritional Biomarkers and Single Nucleotide Polymorphisms
2.4. Statistical Analysis
3. Results
3.1. Folate and Vitamin B12 Intake and Status “Inadequacies”
3.2. Folate, Vitamin B12 Status and Genotype
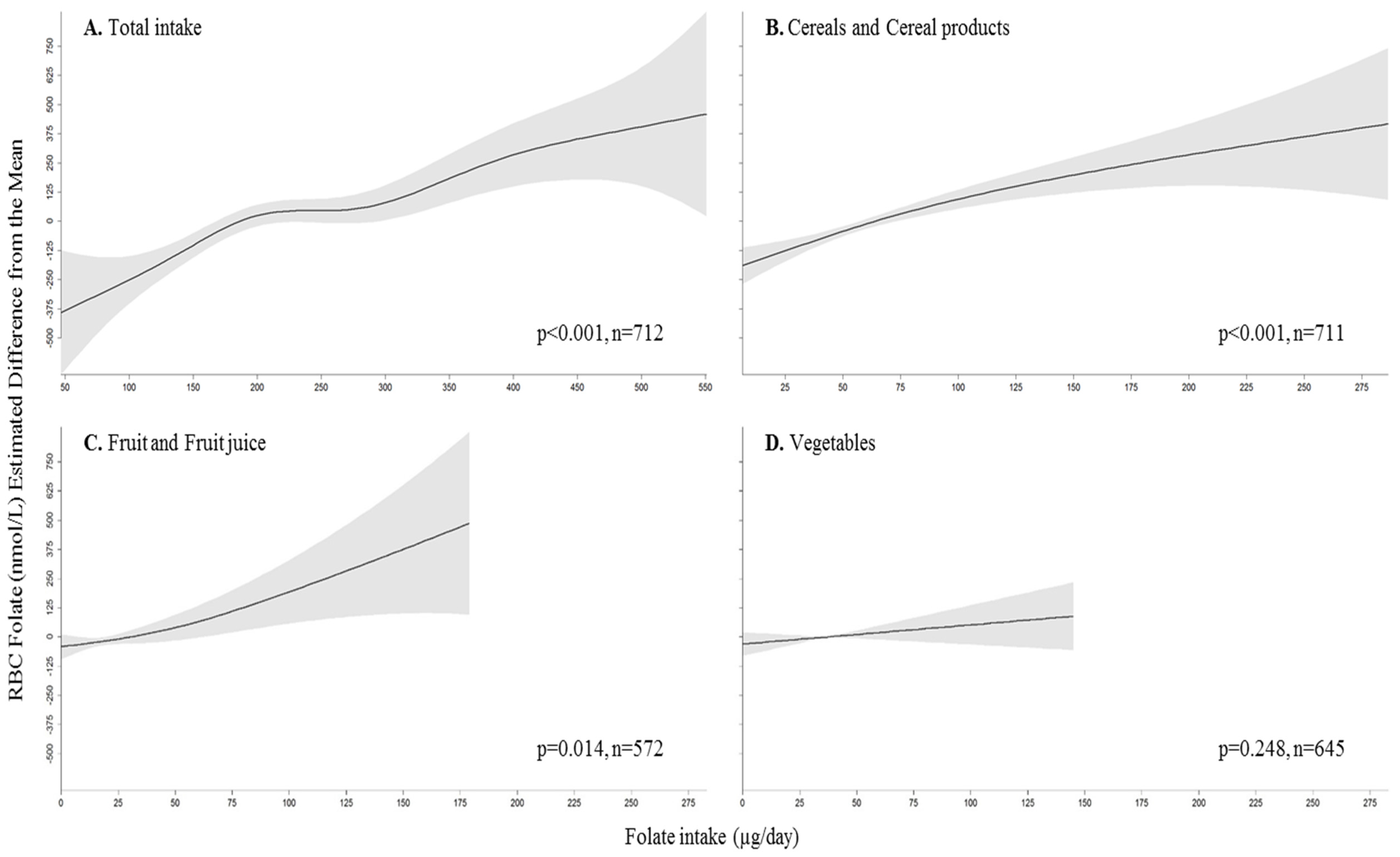
3.3. Association between Folate Intake and Status
3.4. Risk of Low Folate Status by Folate Intake
3.5. Association between Vitamin B12 Intake and Status
3.6. Risk of Deficient Vitamin B12 Status by Vitamin B12 Intake
4. Discussion
4.1. Folate and Vitamin B12 Intake and Status “Inadequacies”
4.2. Association between Folate Intake and Status
4.3. Association between Vitamin B12 Intake and Status
4.4. MTHFR and RBC Folate, and FUT2 and Plasma Vitamin B12
4.5. Strengths and Weaknesses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Locasale, J.W. Serine, glycine and the one-carbon cycle: Cancer metabolism in full circle. Nat. Rev. Cancer 2013, 13, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Durga, J.; van Boxtel, M.P.; Schouten, E.G.; Kok, F.J.; Jolles, J.; Katan, M.B.; Verhoef, P. Effect of 3-year folic acid supplementation on cognitive function in older adults in the facit trial: A randomised, double blind, controlled trial. Lancet 2007, 369, 208–216. [Google Scholar] [CrossRef]
- Nurk, E.; Refsum, H.; Tell, G.S.; Engedal, K.; Vollset, S.E.; Ueland, P.M.; Nygaard, H.A.; Smith, A.D. Plasma total homocysteine and memory in the elderly: The hordaland homocysteine study. Ann. Neurol. 2005, 58, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Kado, D.M.; Karlamangla, A.S.; Huang, M.H.; Troen, A.; Rowe, J.W.; Selhub, J.; Seeman, T.E. Homocysteine versus the vitamins folate, B6, and B12 as predictors of cognitive function and decline in older high-functioning adults: Macarthur studies of successful aging. Am. J. Med. 2005, 118, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Saposnik, G.; Ray, J.G.; Sheridan, P.; McQueen, M.; Lonn, E. Homocysteine-lowering therapy and stroke risk, severity, and disability: Additional findings from the hope 2 trial. Stroke 2009, 40, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Spence, J.D. Metabolic vitamin B12 deficiency: A missed opportunity to prevent dementia and stroke. Nutr. Res. 2016, 36, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Fratoni, V.; Brandi, M.L. B vitamins, homocysteine and bone health. Nutrients 2015, 7, 2176–2192. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Hu, X.; Zhang, Q.; Cao, H.; Wang, J.; Liu, B. Homocysteine level and risk of fracture: A meta-analysis and systematic review. Bone 2012, 51, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Ericson, U.; Sonestedt, E.; Gullberg, B.; Olsson, H.; Wirfalt, E. High folate intake is associated with lower breast cancer incidence in postmenopausal women in the malmo diet and cancer cohort. Am. J. Clin. Nutr. 2007, 86, 434–443. [Google Scholar] [PubMed]
- Ter Borg, S.; Verlaan, S.; Hemsworth, J.; Mijnarends, D.M.; Schols, J.M.; Luiking, Y.C.; de Groot, L.C. Micronutrient intakes and potential inadequacies of community-dwelling older adults: A systematic review. Br. J. Nutr. 2015, 113, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Bates, B.; Lennox, A.; Prentice, A.; Bates, C.; Page, P.; Nicholson, S.; Swan, G. National Diet and Nutrition Survey Years 1–4 (Combined) (2008/2009-2010/2012)-Appendices and Tables. Available online: https://www.gov.uk/government/statistics/national-diet-and-nutrition-survey-results-from-years-1-to-4-combined-of-the-rolling-programme-for-2008-and-2009-to-2011-and-2012 (accessed on 15 August 2015).
- Johnson, M.A. If high folic acid aggravates vitamin B12 deficiency what should be done about it? Nutr. Rev. 2007, 65, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.F.; Ward, M.; Hoey, L.; McNulty, H. Vitamin B12 and ageing: Current issues and interaction with folate. Ann. Clin. Biochem. 2013, 50, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Zinck, J.W.; de Groh, M.; MacFarlane, A.J. Genetic modifiers of folate, vitamin B-12, and homocysteine status in a cross-sectional study of the Canadian population. Am. J. Clin. Nutr. 2015, 101, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Kwan, L.L.; Bermudez, O.I.; Tucker, K.L. Low vitamin B-12 intake and status are more prevalent in Hispanic older adults of Caribbean origin than in neighborhood-matched non-hispanic whites. J. Nutr. 2002, 132, 2059–2064. [Google Scholar] [PubMed]
- Bor, M.V.; Lydeking-Olsen, E.; Moller, J.; Nexo, E. A daily intake of approximately 6 microg vitamin B-12 appears to saturate all the vitamin B-12-related variables in danish postmenopausal women. Am. J. Clin. Nutr. 2006, 83, 52–58. [Google Scholar] [PubMed]
- Powers, H.J.; Hill, M.H.; Welfare, M.; Spiers, A.; Bal, W.; Russell, J.; Duckworth, Y.; Gibney, E.; Williams, E.A.; Mathers, J.C. Responses of biomarkers of folate and riboflavin status to folate and riboflavin supplementation in healthy and colorectal polyp patients (the fab2 study). Cancer Epidemiol. Biomark. Prev. 2007, 16, 2128–2135. [Google Scholar] [CrossRef] [PubMed]
- Bates, C.J.; Schneede, J.; Mishra, G.; Prentice, A.; Mansoor, M.A. Relationship between methylmalonic acid, homocysteine, vitamin B12 intake and status and socio-economic indices, in a subset of participants in the british national diet and nutrition survey of people aged 65 y and over. Eur. J. Clin. Nutr. 2003, 57, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Brouwer-Brolsma, E.M.; Dhonukshe-Rutten, R.A.; van Wijngaarden, J.P.; Zwaluw, N.L.; Velde, N.; de Groot, L.C. Dietary sources of vitamin B-12 and their association with vitamin B-12 status markers in healthy older adults in the B-proof study. Nutrients 2015, 7, 7781–7797. [Google Scholar] [CrossRef] [PubMed]
- Howard, J.M.; Azen, C.; Jacobsen, D.W.; Green, R.; Carmel, R. Dietary intake of cobalamin in elderly people who have abnormal serum cobalamin, methylmalonic acid and homocysteine levels. Eur. J. Clin. Nutr. 1998, 52, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.K.; Wong, K.C.; Wu, M.Y.; Liao, S.L.; Kuo, C.S.; Huang, R.F. Correlations between folate, B12, homocysteine levels, and radiological markers of neuropathology in elderly post-stroke patients. J. Am. Coll. Nutr. 2007, 26, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Van Guelpen, B.; Hultdin, J.; Johansson, I.; Stegmayr, B.; Hallmans, G.; Nilsson, T.K.; Weinehall, L.; Witthoft, C.; Palmqvist, R.; Winkvist, A. Folate, vitamin B12, and risk of ischemic and hemorrhagic stroke: A prospective, nested case-referent study of plasma concentrations and dietary intake. Stroke 2005, 36, 1426–1431. [Google Scholar] [CrossRef] [PubMed]
- EFSA NDA Panel (EFSA Panel on Dietetic Products Nutrition and Allergies). Scientific opinion on dietary reference values for folate. EFSA J. 2015, 12, 3893. [Google Scholar]
- EFSA NDA Panel (EFSA Panel on Dietetic Products Nutrition and Allergies). Scientific opinion on dietary reference values for cobalamin (vitamin B12). EFSA J. 2015, 13, 4150. [Google Scholar]
- Collerton, J.; Davies, K.; Jagger, C.; Kingston, A.; Bond, J.; Eccles, M.P.; Robinson, L.A.; Martin-Ruiz, C.; von Zglinicki, T.; James, O.F.; et al. Health and disease in 85 year olds: Baseline findings from the newcastle 85+ cohort study. BMJ 2009, 339, b4904. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.R.; Mendonça, N.; Granic, A.; Siervo, M.; Jagger, C.; Seal, C.J.; Kerse, N.; Wham, C.; Adamson, A.J.; Mathers, J.C. What do we know about the nutritional status of the very old? Insights from three cohorts of advanced age from the UK and New Zealand. Proc. Nutr. Soc. 2016, 75, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.; Kingston, A.; Robinson, L.; Hughes, J.; Hunt, J.M.; Barker, S.A.H.; Edwards, J.; Collerton, J.; Jagger, C.; Kirkwood, T.B.L. Improving retention of very old participants in longitudinal research: Experiences from the newcastle 85+ study. PLoS ONE 2014, 9, e108370. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, N.; Hill, T.R.; Granic, A.; Mathers, J.C.; Wrieden, W.; Siervo, M.; Seal, C.; Jagger, C.; Adamson, A.J. Macronutrient intake and food sources in the very old: Analysis of the Newcastle 85+ study. Br. J. Nutr. 2016, 115, 2170–2180. [Google Scholar] [CrossRef] [PubMed]
- Food Standards Agency. Mccance and Widdowson's the Composition of Foods, 6th ed.Royal Society of Chemistry: Cambridge, UK, 2002.
- Mendonça, N.; Hill, T.R.; Granic, A.; Mathers, J.C.; Wrieden, W.; Siervo, M.; Seal, C.; Jagger, C.; Adamson, A.J. Micronutrient intake and food sources in the very old: Analysis of the Newcastle 85+ study. Br. J. Nutr. 2016, 116, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Martin-Ruiz, C.; Jagger, C.; Kingston, A.; Collerton, J.; Catt, M.; Davies, K.; Dunn, M.; Hilkens, C.; Keavney, B.; Pearce, S.H.S.; et al. Assessment of a large panel of candidate biomarkers of ageing in the newcastle 85+ study. Mech. Ageing Dev. 2011, 132, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Deelen, J.; Beekman, M.; Uh, H.W.; Broer, L.; Ayers, K.L.; Tan, Q.; Kamatani, Y.; Bennet, A.M.; Tamm, R.; Trompet, S.; et al. Genome-wide association meta-analysis of human longevity identifies a novel locus conferring survival beyond 90 years of age. Hum. Mol. Genet. 2014, 23, 4420–4432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Department of Health: Committee on Medical Aspects of Food Policy (COMA). Report on Health and Social Subjects 41: Dietary Reference Values for Food Energy and Nutrients for the United Kingdom; Department of Health: Committee on Medical Aspects of Food Policy: London, UK, 1991.
- Vogiatzoglou, A.; Oulhaj, A.; Smith, A.D.; Nurk, E.; Drevon, C.A.; Ueland, P.M.; Vollset, S.E.; Tell, G.S.; Refsum, H. Determinants of plasma methylmalonic acid in a large population: Implications for assessment of vitamin B12 status. Clin. Chem. 2009, 55, 2198–2206. [Google Scholar] [CrossRef] [PubMed]
- Bang, H.; Mazumdar, M.; Spence, J.D. Tutorial in biostatistics: Analyzing associations between total plasma homocysteine and B vitamins using optimal categorization and segmented regression. Neuroepidemiology 2006, 27, 188–200. [Google Scholar] [CrossRef] [PubMed]
- The 1000 Genomes Project Consortium. 1000 Genomes Project Phase 3. Available online: http://browser.1000genomes.org/Homo_sapiens/Variation/Population?r=1:11855878-11856878;v=rs1801133;vdb=variation;vf=1230309 (accessed on 10 December 2015).
- Pfeiffer, C.M.; Hughes, J.P.; Lacher, D.A.; Bailey, R.L.; Berry, R.J.; Zhang, M.; Yetley, E.A.; Rader, J.I.; Sempos, C.T.; Johnson, C.L. Estimation of trends in serum and RBC folate in the U.S. Population from pre- to postfortification using assay-adjusted data from the NHANES 1988–2010. J. Nutr. 2012, 142, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine (US) Food and Nutrition Board. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic acid, Biotin, and Choline; National Academy Press: Washington, DC, USA, 1998. [Google Scholar]
- Bailey, R.L.; Fulgoni, V.L.; Keast, D.R.; Dwyer, J.T. Examination of vitamin intakes among us adults by dietary supplement use. J. Acad. Nutr. Diet. 2012, 112, 657–663.e4. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Cogswell, M.E.; Hamner, H.C.; Carriquiry, A.; Bailey, L.B.; Pfeiffer, C.M.; Berry, R.J. Folic acid source, usual intake, and folate and vitamin B-12 status in us adults: National health and nutrition examination survey (NHANES) 2003–2006. Am. J. Clin. Nutr. 2010, 91, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Caudill, M.A. Folate bioavailability: Implications for establishing dietary recommendations and optimizing status. Am. J. Clin. Nutr. 2010, 91, 1455S–1460S. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, I.A.; van Dusseldorp, M.; West, C.E.; Steegers-Theunissen, R.P. Bioavailability and bioefficacy of folate and folic acid in man. Nutr. Res. Rev. 2001, 14, 267–294. [Google Scholar] [CrossRef] [PubMed]
- Hoey, L.; McNulty, H.; Askin, N.; Dunne, A.; Ward, M.; Pentieva, K.; Strain, J.; Molloy, A.M.; Flynn, C.A.; Scott, J.M. Effect of a voluntary food fortification policy on folate, related B vitamin status, and homocysteine in healthy adults. Am. J. Clin. Nutr. 2007, 86, 1405–1413. [Google Scholar] [PubMed]
- Doscherholmen, A.; McMahon, J.; Ripley, D. Vitamin B12 assimilation from chicken meat. Am. J. Clin. Nutr. 1978, 31, 825–830. [Google Scholar] [PubMed]
- Heyssel, R.M.; Bozian, R.C.; Darby, W.J.; Bell, M.C. Vitamin B12 turnover in man. The assimilation of vitamin B12 from natural foodstuff by man and estimates of minimal daily dietary requirements. Am. J. Clin. Nutr. 1966, 18, 176–184. [Google Scholar] [PubMed]
- Vogiatzoglou, A.; Smith, A.D.; Nurk, E.; Berstad, P.; Drevon, C.A.; Ueland, P.M.; Vollset, S.E.; Tell, G.S.; Refsum, H. Dietary sources of vitamin B-12 and their association with plasma vitamin B-12 concentrations in the general population: The hordaland homocysteine study. Am. J. Clin. Nutr. 2009, 89, 1078–1087. [Google Scholar] [CrossRef] [PubMed]
- Suter, P.M.; Golner, B.B.; Goldin, B.R.; Morrow, F.D.; Russell, R.M. Reversal of protein-bound vitamin B12 malabsorption with antibiotics in atrophic gastritis. Gastroenterology 1991, 101, 1039–1045. [Google Scholar] [CrossRef]
- Russell, R.M.; Baik, H.; Kehayias, J.J. Older men and women efficiently absorb vitamin B-12 from milk and fortified bread. J. Nutr. 2001, 131, 291–293. [Google Scholar] [PubMed]
- Tanaka, T.; Scheet, P.; Giusti, B.; Bandinelli, S.; Piras, M.G.; Usala, G.; Lai, S.; Mulas, A.; Corsi, A.M.; Vestrini, A.; et al. Genome-wide association study of vitamin B6, vitamin B12, folate, and homocysteine blood concentrations. Am. J. Hum. Genet. 2009, 84, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Hazra, A.; Kraft, P.; Selhub, J.; Giovannucci, E.L.; Thomas, G.; Hoover, R.N.; Chanock, S.J.; Hunter, D.J. Common variants of FUT2 are associated with plasma vitamin B12 levels. Nat. Genet. 2008, 40, 1160–1162. [Google Scholar] [CrossRef] [PubMed]
- Bates, C.J.; Prentice, A.; van der Pols, J.C.; Walmsley, C.; Pentieva, K.D.; Finch, S.; Smithers, G.; Clarke, P.C. Estimation of the use of dietary supplements in the national diet and nutrition survey: People aged 65 years and over. An observed paradox and a recommendation. Eur. J. Clin. Nutr. 1998, 52, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Hasuike, Y.; Nakanishi, T.; Moriguchi, R.; Otaki, Y.; Nanami, M.; Hama, Y.; Naka, M.; Miyagawa, K.; Izumi, M.; Takamitsu, Y. Accumulation of cyanide and thiocyanate in haemodialysis patients. Nephrol. Dial. Transplant. 2004, 19, 1474–1479. [Google Scholar] [CrossRef] [PubMed]
- Castro, R.; Barroso, M.; Rocha, M.; Esse, R.; Ramos, R.; Ravasco, P.; Rivera, I.; de Almeida, I.T. The TCN2 776CNG polymorphism correlates with vitamin B (12) cellular delivery in healthy adult populations. Clin. Biochem. 2010, 43, 645–649. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| All | Men | Women | p-Value 1 | |
|---|---|---|---|---|
| Sex (%) (n) | 732 | 39 (287) | 61 (445) | - |
| BMI (kg/m2) (mean ± SD) | 24.4 ± 4.3 | 24.7 ± 3.9 | 24.3 ± 4.6 | 0.244 2 |
| Smokers (%) (n) | 5.6 (41) | 4.2 (12) | 6.5 (29) | 0.183 |
| Alcohol Drinkers (%) (n) | 72 (364) | 84 (192) | 62 (172) | <0.001 |
| Total Energy Intakes (MJ/day) | 6.78 (5.62–8.31) | 8.01 (6.65–9.59) | 6.26 (5.17–7.38) | <0.001 |
| Folate and vitamin B12 supplement use (%) (n) | 4.8 (35) | 3.8 (11) | 5.4 (24) | 0.334 |
| H2 antagonists, PPI and biguanides use (%) (n) | 26.8 (196) | 27.2 (78) | 26.5 (118) | 0.844 |
| Total Homocysteine (µmol/L) | 16.7 (13.5–21.4) | 18.0 (14.5–21.9) | 16.1 (13.1–21.0) | 0.001 |
| >15 µmol/L (%) (n) | 63.1 (471) | 70.3 (206) | 58.5 (265) | 0.001 |
| Folate | ||||
| Intake (µg/day) | 209 (157–265) | 246 (185–296) | 189 (144–242) | <0.001 |
| <100 µg/day (%) (n) | 3.1 (23) | 0.7 (2) | 4.7 (21) | 0.002 |
| Top food group contributors | Cereals (32%), Vegetables (16%), Fruit (9%) | Cereals (32%), Vegetables (15%), Fruit (8%) | Cereals (31%), Vegetables (17%), Fruit (10%) | - |
| Red Blood Cell Folate (nmol/L) | 863 (451–1287) | 868 (596–1282) | 854 (614–1287) | 0.728 |
| <340 nmol/L (%) (n) | 3.6 (26) | 2.1 (6) | 4.5 (20) | 0.103 |
| Vitamin B12 | ||||
| Intake (µg/day) | 2.9 (1.9–4.4) | 3.5 (2.2–5.2) | 2.5 (1.6–3.9) | <0.001 |
| <1.0 µg/day (%) (n) | 9.2 (67) | 4.5 (13) | 12.1 (54) | <0.001 |
| Top food group contributors | Meat (53%), Fish (17%), Milk (13%) | Meat (59%), Fish (16%), Milk (10%) | Meat (48%), Fish (19%), Milk (15%) | - |
| Plasma Vitamin B12 (pmol/L) | 232 (170–324) | 228 (166–309) | 238 (174–337) | 0.238 |
| <148 pmol/L (%) (n) | 17.1 (125) | 17.4 (50) | 16.9 (75) | 0.841 |
| RBC Folate (nmol/L) | p-Value 1 | Plasma Vitamin B12 (pmol/L) | p-Value 1 | |
|---|---|---|---|---|
| FUT2 (rs492602) | 0.531 | <0.001 | ||
| AA (n = 128) | 894 (629–1349 | 216 (146–281) | Ref. | |
| A/G (n = 308) | 917 (603–1322) | 221 (163–309) | 0.413 | |
| GG (n = 187) | 835 (595–1206) | 277 (209–381) | <0.001 | |
| MTHFR (rs1801133) | 0.028 | 0.244 | ||
| GG (n = 276) | 871 (614–1275) | Ref. | 234 (168–331) | |
| A/G (n = 279) | 845 (584–1263) | 1.000 | 230 (164–312) | |
| AA (n = 67) | 1010 (693–1626) | 0.060 | 249 (193–339) | |
| MTR (rs1805087) | 0.547 | 0.277 | ||
| AA (n = 419) | 881 (613–1278) | 240 (173–337) | ||
| A/G (n = 178) | 845 (596–1332) | 226 (162–297) | ||
| GG (n = 26) | 1053 (580–1593) | 247 (162–310) | ||
| TCN1 (rs526934) | 0.065 | 0.298 | ||
| AA (n = 331) | 877 (606–1317) | 237 (178–336) | ||
| A/G (n = 247) | 845 (595–1223) | 231 (160–325) | ||
| GG (n = 45) | 1074 (630–1439) | 222 (182–273) | ||
| Folate Intake | Model 1 (Unadjusted) | Model 2 (Adjusted) | ||
|---|---|---|---|---|
| Total (µg/day) | <600 nmol/L (n = 170) | p | <600 nmol/L (n = 170) | p |
| <157 | 1.00 (ref.) | - | 1.00 (ref.) | - |
| 157–208 | 0.64 (0.40, 1.04) | 0.071 | 0.65 (0.38, 1.09) | 0.103 |
| 209–264 | 0.72 (0.45, 1.15) | 0.173 | 0.58 (0.34, 1.02) | 0.057 |
| >264 | 0.58 (0.36, 0.94) | 0.028 | 0.43 (0.23, 0.82) | 0.010 |
| Cereals and Cereal products (µg/day) | <600 nmol/L (n = 170) | p | <600 nmol/L (n = 170) | p |
| <36 | 1.00 (ref.) | - | 1.00 (ref.) | - |
| 36–59 | 0.96 (0.61, 1.49) | 0.840 | 0.84 (0.51, 1.38) | 0.493 |
| 59–92 | 0.40 (0.24, 0.66) | <0.001 | 0.32 (0.18, 0.57) | <0.001 |
| >92 | 0.41 (0.25, 0.68) | 0.001 | 0.33 (0.18, 0.61) | <0.001 |
| Vegetables (µg/day) | <600 nmol/L (n = 154) | p | <600 nmol/L (n = 154) | p |
| <15 | 1.00 (ref.) | - | 1.00 (ref.) | - |
| 15–30 | 0.72 (0.43, 1.21) | 0.212 | 0.49 (0.25, 0.95) | 0.035 |
| 30–51 | 0.86 (0.52, 1.41) | 0.550 | 0.59 (0.32, 1.08) | 0.089 |
| >51 | 0.79 (0.48, 1.30) | 0.357 | 0.52 (0.28, 0.99) | 0.045 |
| Fruit and Fruit Juice (µg/day) | <600 nmol/L (n = 127) | p | <600 nmol/L (n = 127) | p |
| <7.3 | 1.00 (ref.) | - | 1.00 (ref.) | - |
| 7.3–16 | 0.90 (0.53, 1.52) | 0.682 | 1.01 (0.56, 1.83) | 0.979 |
| 16–34 | 0.61 (0.35, 1.07) | 0.086 | 0.67 (0.36, 1.25) | 0.213 |
| >34 | 0.76 (0.44, 1.31) | 0.329 | 0.79 (0.43, 1.44) | 0.437 |
| Vitamin B12 Intake | Model 1 (Unadjusted) | Model 2 (Adjusted) | ||
|---|---|---|---|---|
| Total (µg/day) | <148 pmol/L (n = 125) | p | <148 pmol/L (n = 125) | p |
| <1.87 | 1.00 (ref.) | - | 1.00 (ref.) | - |
| 1.87–2.88 | 0.70 (0.42, 1.18) | 0.180 | 0.57 (0.32, 1.01) | 0.056 |
| 2.88–4.40 | 0.60 (0.35, 1.02) | 0.057 | 0.50 (0.28, 0.92) | 0.026 |
| >4.40 | 0.53 (0.31, 0.92) | 0.024 | 0.40 (0.21, 0.76) | 0.005 |
| Meat and Meat products (µg/day) | <148 pmol/L (n = 118) | p | <148 pmol/L (n = 118) | p |
| <0.35 | 1.00 (ref.) | - | 1.00 (ref.) | - |
| 0.35–1.03 | 0.72 (0.42, 1.24) | 0.236 | 0.69 (0.38, 1.25) | 0.220 |
| 1.03–2.10 | 0.84 (0.50, 1.44) | 0.533 | 0.78 (0.43, 1.42) | 0.422 |
| >2.10 | 0.55 (0.31, 0.98) | 0.043 | 0.41 (0.20, 0.81) | 0.010 |
| Fish and Fish products (µg/day) | <148 pmol/L (n = 43) | p | <148 pmol/L (n = 43) | p |
| <0.46 | 1.00 (ref.) | - | 1.00 (ref.) | - |
| 0.46–1.06 | 0.61 (0.23, 1.65) | 0.331 | 0.66 (0.23, 1.91) | 0.444 |
| 1.06–2.45 | 0.86 (0.34, 2.15) | 0.743 | 0.66 (0.23, 1.86) | 0.427 |
| >2.45 | 1.00 (0.41, 2.42) | 0.992 | 0.70 (0.25, 1.97) | 0.503 |
| Milk and Milk products (µg/day) | <148 pmol/L (n = 102) | p | <148 pmol/L (n = 102) | p |
| <0.27 | 1.00 (ref.) | - | 1.00 (ref.) | - |
| 0.27–0.53 | 0.84 (0.47, 1.52) | 0.562 | 0.88 (0.46, 1.71) | 0.711 |
| 0.53–0.88 | 1.12 (0.64, 1.96) | 0.698 | 1.28 (0.70, 2.37) | 0.425 |
| >0.88 | 0.58 (0.31, 1.08) | 0.086 | 0.49 (0.24, 1.01) | 0.054 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendonça, N.; Mathers, J.C.; Adamson, A.J.; Martin-Ruiz, C.; Seal, C.J.; Jagger, C.; Hill, T.R. Intakes of Folate and Vitamin B12 and Biomarkers of Status in the Very Old: The Newcastle 85+ Study. Nutrients 2016, 8, 604. https://doi.org/10.3390/nu8100604
Mendonça N, Mathers JC, Adamson AJ, Martin-Ruiz C, Seal CJ, Jagger C, Hill TR. Intakes of Folate and Vitamin B12 and Biomarkers of Status in the Very Old: The Newcastle 85+ Study. Nutrients. 2016; 8(10):604. https://doi.org/10.3390/nu8100604
Chicago/Turabian StyleMendonça, Nuno, John C. Mathers, Ashley J. Adamson, Carmen Martin-Ruiz, Chris J. Seal, Carol Jagger, and Tom R. Hill. 2016. "Intakes of Folate and Vitamin B12 and Biomarkers of Status in the Very Old: The Newcastle 85+ Study" Nutrients 8, no. 10: 604. https://doi.org/10.3390/nu8100604
APA StyleMendonça, N., Mathers, J. C., Adamson, A. J., Martin-Ruiz, C., Seal, C. J., Jagger, C., & Hill, T. R. (2016). Intakes of Folate and Vitamin B12 and Biomarkers of Status in the Very Old: The Newcastle 85+ Study. Nutrients, 8(10), 604. https://doi.org/10.3390/nu8100604