Sake Protein Supplementation Affects Exercise Performance and Biochemical Profiles in Power-Exercise-Trained Mice



Abstract
:1. Introduction
2. Experimental Section
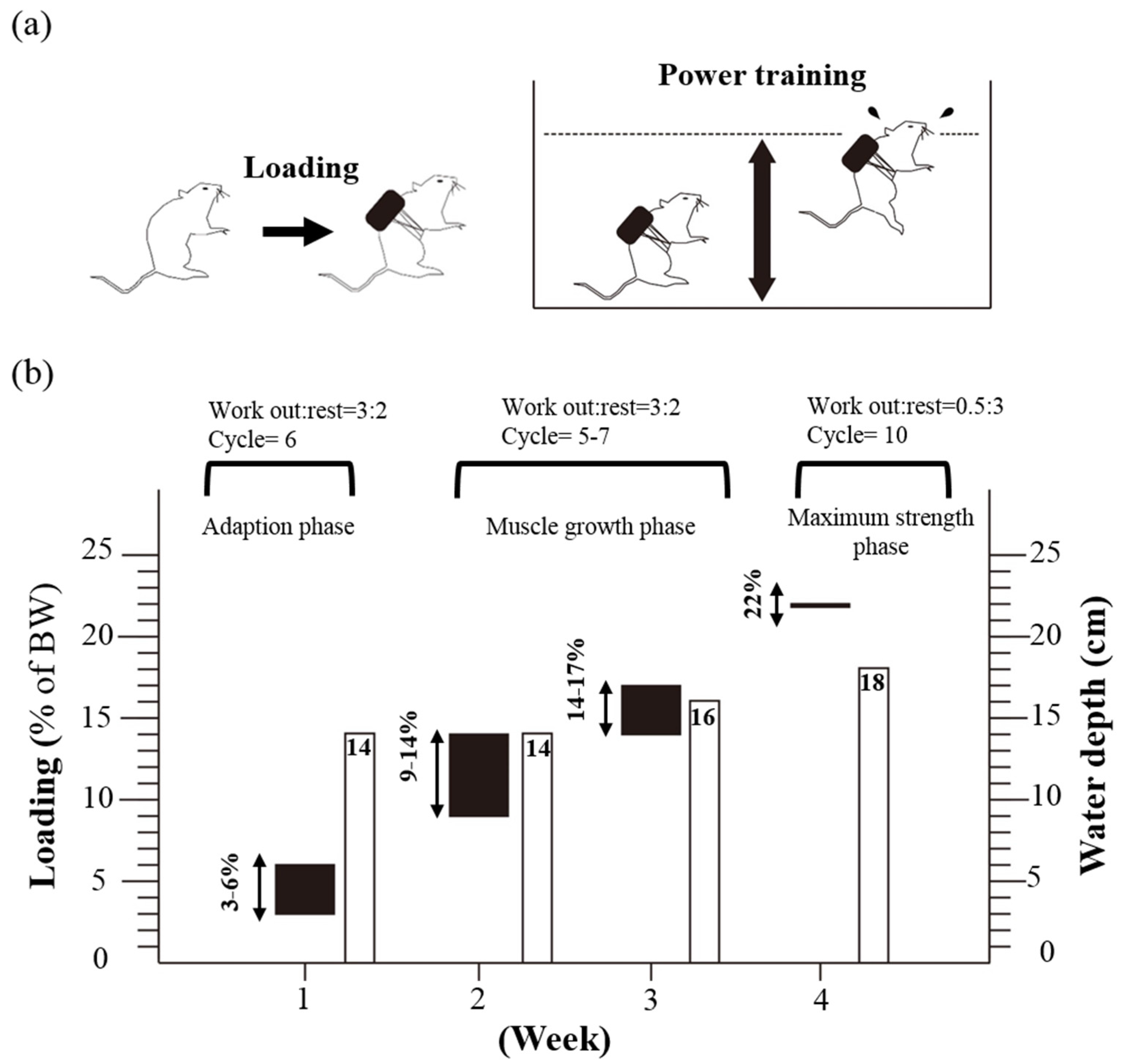
2.1. Materials, Animals, and Experiment Design
2.2. SP Supplementation
2.3. PET Protocol
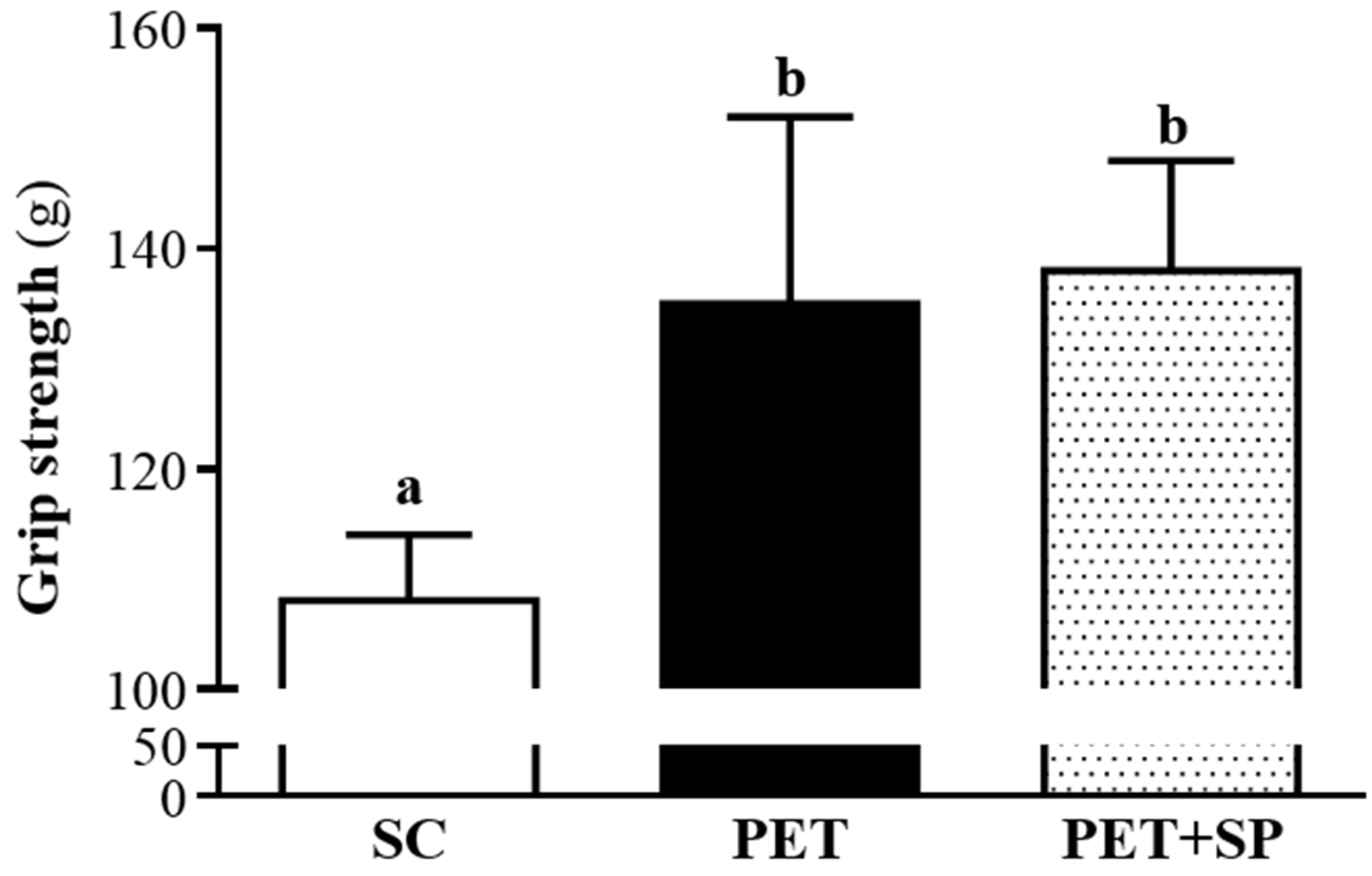
2.4. Forelimb Grip Strength
2.5. Swimming Exercise Performance Test
2.6. Determination of Blood Biochemical Variables
2.7. Tissue Glycogen Determination
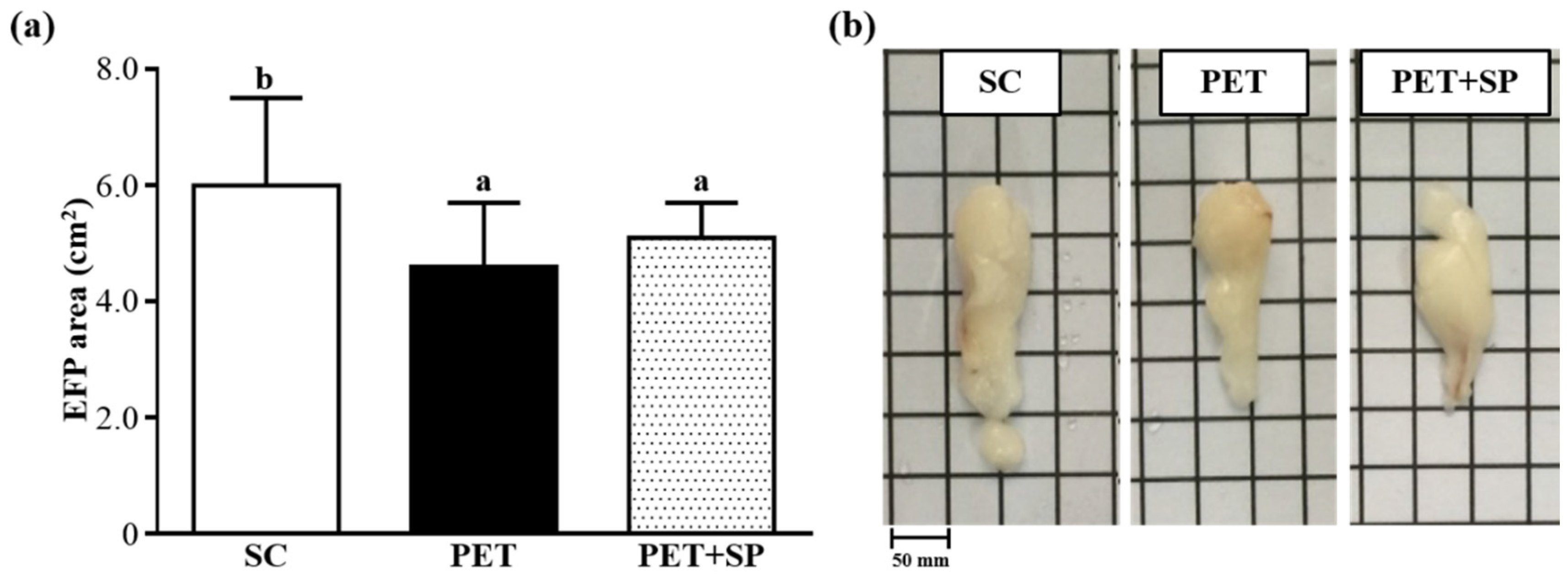
2.8. Histological Staining of Tissues and Calculated Epididymal Fat Pad Area
2.9. Statistical Analysis
3. Results and Discussion
3.1. Nutrition Facts and Total BCAAs of SP
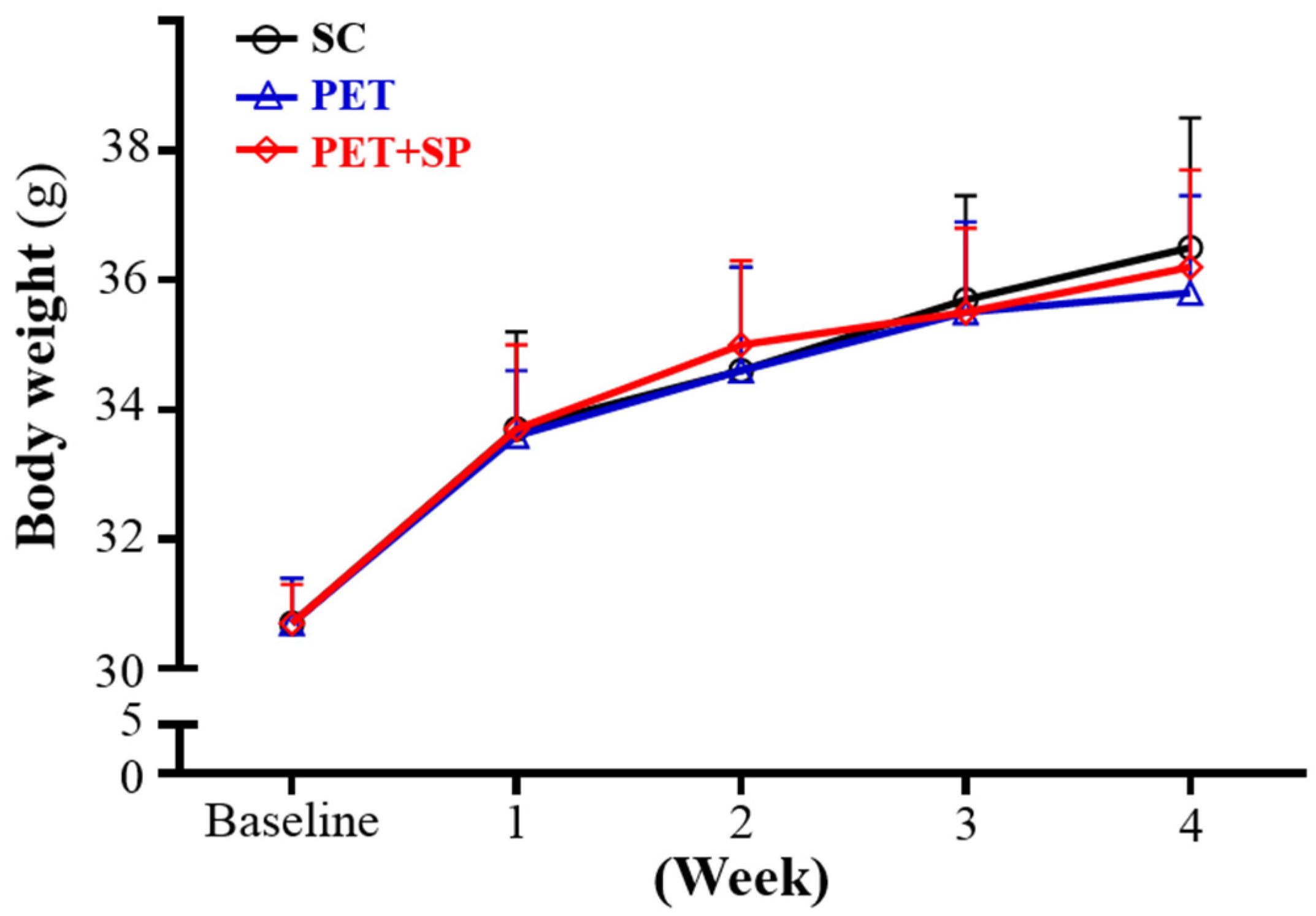
3.2. Effect of SP Supplementation and PET on Body Weight (BW), Skeletal Muscle Mass, and Other Metabolism-Related Organ Weights
3.3. Effect of SP Supplementation with PET Program on Forelimb Grip Strength
3.4. Effect of SP Supplementation with the PET Program on Exercise Performance in a Weight-Loaded Swimming Test
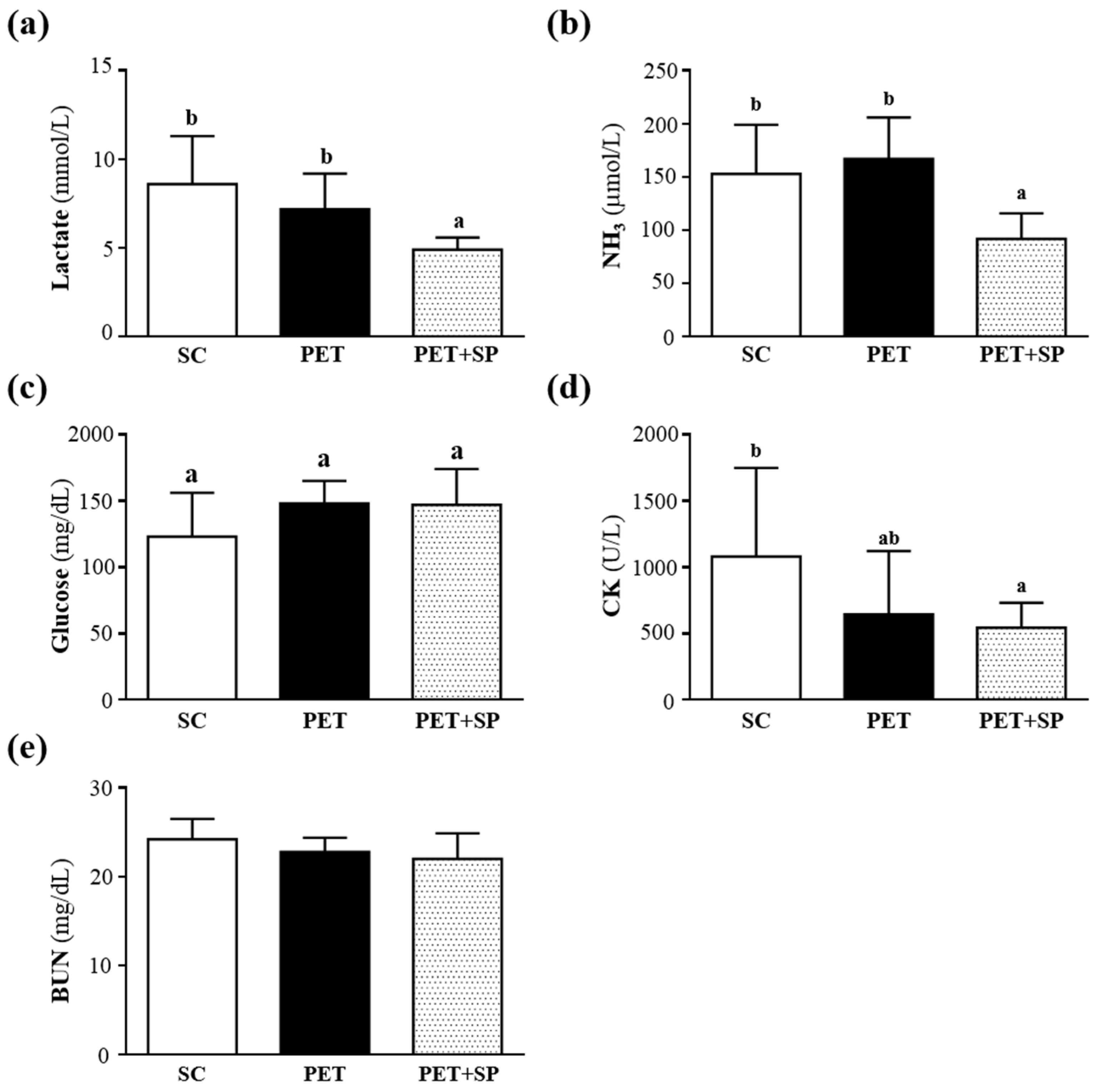
3.5. Effect of SP Supplementation under the PET Program on Serum Lactate, Ammonia, Glucose, CK and BUN Levels after Acute Exercise Challenge
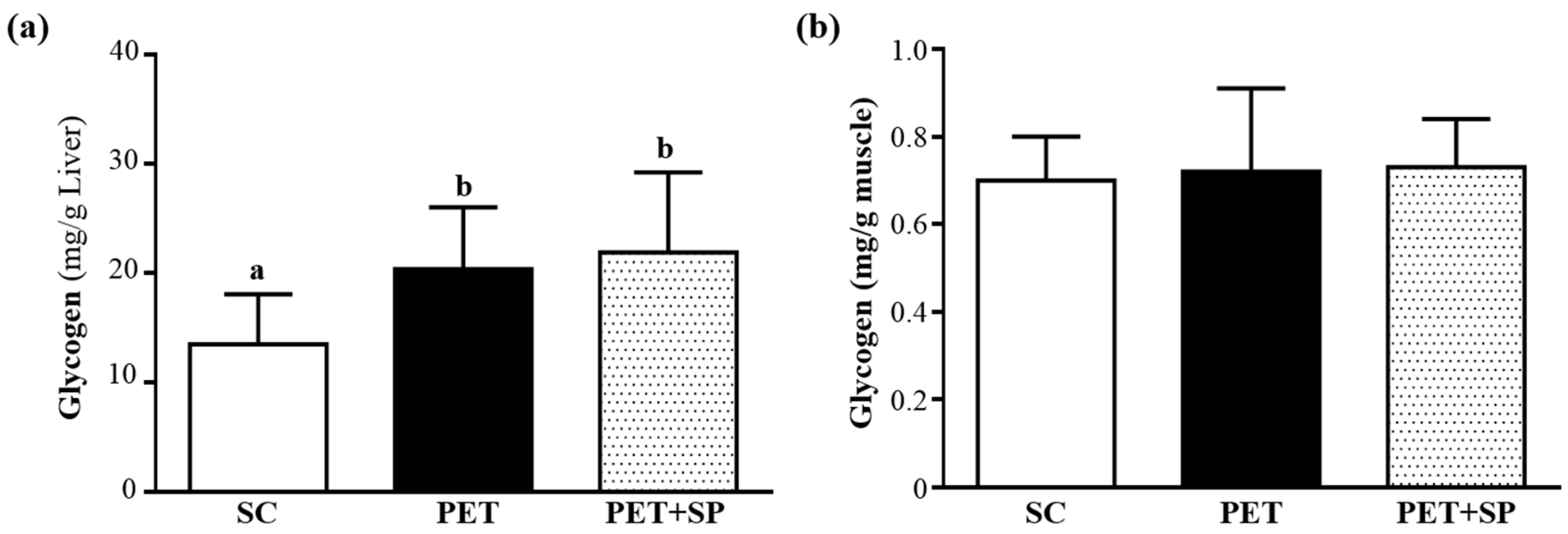
3.6. Effect of PET and PET+SP on Hepatic and Muscle Glycogen Level
3.7. Effect of PET and PET+SP on Biochemical Analyses at the End of the Experiment
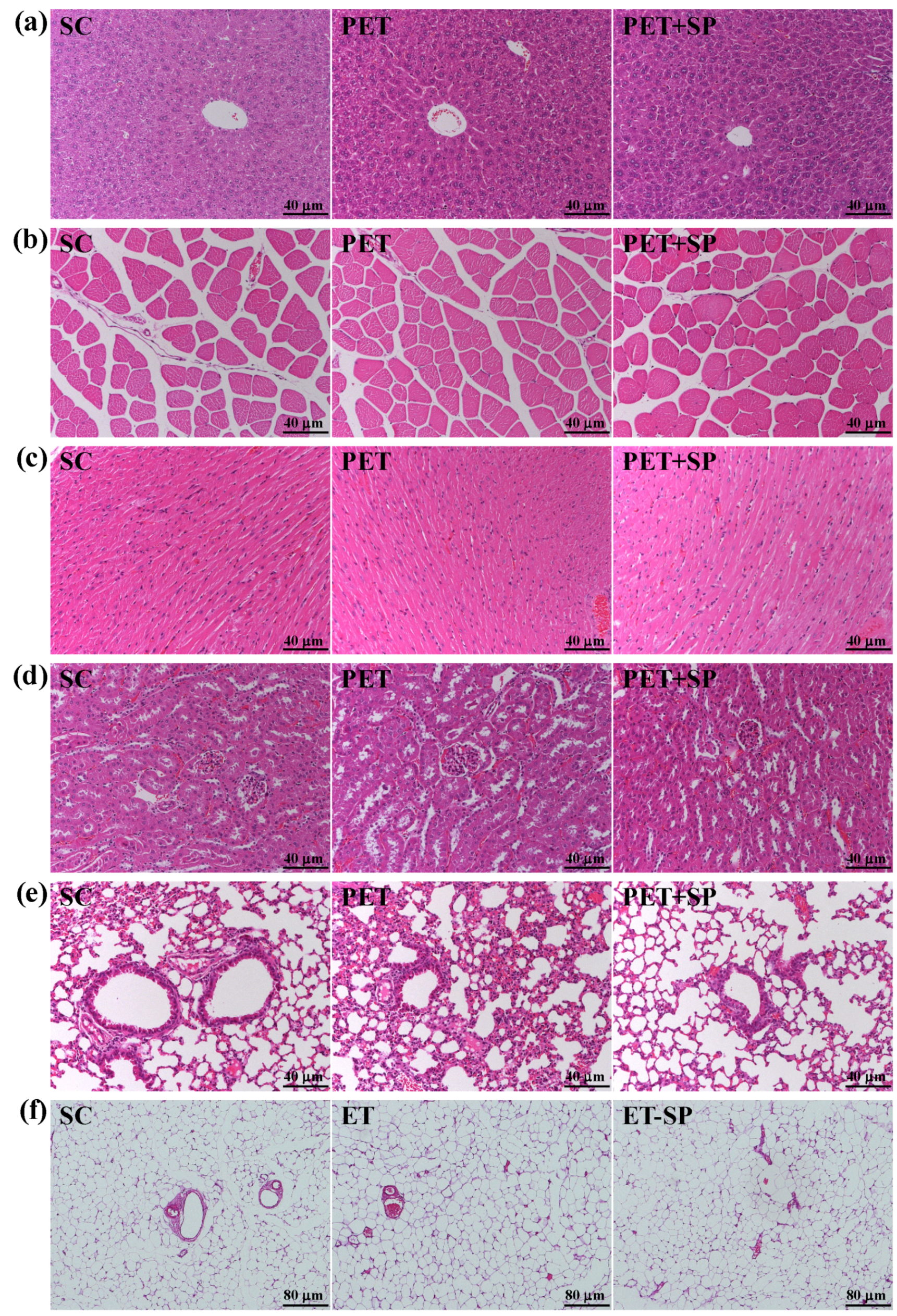
3.8. Histopathological Evaluation of PET and PET + SP Treatments at the End of the Experiment
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Witard, O.C.; Jackman, S.R.; Breen, L.; Smith, K.; Selby, A.; Tipton, K.D. Myofibrillar muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after resistance exercise. Am. J. Clin. Nutr. 2014, 99, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Breen, L.; Burd, N.A.; Hector, A.J.; Churchward-Venne, T.A.; Josse, A.R.; Phillips, S.M. Resistance exercise enhances myofibrillar protein synthesis with graded intakes of whey protein in older men. Br. J. Nutr. 2012, 108, 1780–1788. [Google Scholar] [CrossRef] [PubMed]
- Anthony, J.C.; Anthony, T.G.; Kimball, S.R.; Vary, T.C.; Jefferson, L.S. Orally administered leucine stimulates protein synthesis in skeletal muscle of postabsorptive rats in association with increased eIF4F formation. J. Nutr. 2000, 130, 139–145. [Google Scholar] [PubMed]
- Anthony, J.C.; Yoshizawa, F.; Anthony, T.G.; Vary, T.C.; Jefferson, L.S.; Kimball, S.R. Leucine stimulates translation initiation in skeletal muscle of postabsorptive rats via a rapamycin-sensitive pathway. J. Nutr. 2000, 130, 2413–2419. [Google Scholar] [PubMed]
- Dreyer, H.C.; Fujita, S.; Cadenas, J.G.; Chinkes, D.L.; Volpi, E.; Rasmussen, B.B. Resistance exercise increases AMPK activity and reduces 4E-BP1 phosphorylation and protein synthesis in human skeletal muscle. J. Physiol. 2006, 576, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Pallafacchina, G.; Calabria, E.; Serrano, A.L.; Kalhovde, J.M.; Schiaffino, S.A. Protein kinase B-dependent and rapamycin-sensitive pathway controls skeletal muscle growth but not fiber type specification. Proc. Natl. Acad. Sci. USA 2002, 99, 9213–9218. [Google Scholar] [CrossRef] [PubMed]
- Grier, T.; Canham-Chervak, M.; McNulty, V.; Jones, B.H. Extreme conditioning programs and injury risk in a US Army Brigade Combat Team. US Army Med. Dep. J. 2013, 36–47. [Google Scholar]
- Simenz, C.J.; Dugan, C.A.; Ebben, W.P. Strength and conditioning practices of National Basketball Association strength and conditioning coaches. J. Strength Cond. Res. 2005, 19, 495–504. [Google Scholar] [PubMed]
- Gibala, M.J.; Little, J.P.; MacDonald, M.J.; Hawley, J.A. Physiological adaptations to low-volume, high-intensity interval training in health and disease. J. Physiol. 2012, 590, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Grier, T.; Canham-Chervak, M.; Anderson, M.K.; Bushman, T.T.; Jones, B.H. The effects of cross-training on fitness and injury in women. US Army Med. Dep. J. 2015, 33–41. [Google Scholar]
- Lawton, T.W.; Cronin, J.B.; McGuigan, M.R. Strength, power, and muscular endurance exercise and elite rowing ergometer performance. J. Strength Cond. Res. 2013, 27, 1928–1935. [Google Scholar] [CrossRef] [PubMed]
- Baechle, T.R.; Earle, R.W. Essentials of Strength Training and Conditioning, 3rd ed.; Human kinetics: Champaign, IL, USA, 2008. [Google Scholar]
- Department of Health and Human Services, Food and Drug Administration. Guidance for Industry on Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers. Available online: https://www.federalregister.gov/articles/2005/07/22/05-14456/guidance-for-industry-on-estimating-the-maximum-safe-starting-dose-in-initial-clinical-trials-for (accessed on 17 February 2016).
- Noyes, F.R.; Barber-Westin, S.D.; Smith, S.T.; Campbell, T.; Garrison, T.T. A training program to improve neuromuscular and performance indices in female high school basketball players. J. Strength Cond. Res. 2012, 26, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Simão, R.; Spineti, J.; de Salles, B.F.; Matta, T.; Fernandes, L.; Fleck, S.J.; Strom-Olsen, H.E. Comparison between nonlinear and linear periodized resistance training: Hypertrophic and strength effects. J. Strength Cond. Res. 2012, 26, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.M.; Sommer, A.J.; Starkoff, B.E.; Devor, S.T. Crossfit-based high-intensity power training improves maximal aerobic fitness and body composition. J. Strength Cond. Res. 2013, 27, 3159–3172. [Google Scholar] [CrossRef] [PubMed]
- Yeh, T.S.; Chuang, H.L.; Huang, W.C.; Chen, Y.M.; Huang, C.C.; Hsu, M.C. Astragalus membranaceus improves exercise performance and ameliorates exercise-induced fatigue in trained mice. Molecules 2014, 19, 2793–2807. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Huang, W.C.; Liu, C.C.; Wang, M.F.; Ho, C.S.; Huang, W.P.; Huang, C.C. Pumpkin (Cucurbita moschata) fruit extract improves physical fatigue and exercise performance in mice. Molecules 2012, 17, 11864–11876. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Tseng, T.L.; Huang, W.C.; Chung, Y.H.; Chuang, H.L.; Wu, J.H. Whole-body vibration training effect on physical performance and obesity in mice. Int. J. Med. Sci. 2014, 11, 1218–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, R.E.; Huang, W.C.; Liao, C.C.; Chang, Y.K.; Kan, N.W.; Huang, C.C. Resveratrol protects against physical fatigue and improves exercise performance in mice. Molecules 2013, 18, 4689–4702. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.M.; Tsai, Y.H.; Tsai, T.Y.; Chiu, Y.S.; Wei, L.; Chen, W.C.; Huang, C.C. Fucoidan supplementation improves exercise performance and exhibits anti-fatigue action in mice. Nutrients 2014, 7, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Chiu, W.C.; Chuang, H.L.; Tang, D.W.; Lee, Z.M.; Wei, L.; Huang, C.C. Effect of curcumin supplementation on physiological fatigue and physical performance in mice. Nutrients 2015, 7, 905–921. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.J.; Chiu, C.C.; Li, Y.P.; Huang, W.C.; Te Huang, Y.; Huang, C.C.; Chuang, H.L. Effect of intestinal microbiota on exercise performance in mice. J. Strength Cond. Res. 2015, 29, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, H.K.; Nilsson, P.A.; Nilsson, J.; Chibalin, A.V.; Zierath, J.R.; Blomstrand, E. Branched-chain amino acids increase p70S6k phosphorylation in human skeletal muscle after resistance exercise. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E1–E7. [Google Scholar] [CrossRef] [PubMed]
- Willis, L.H.; Slentz, C.A.; Bateman, L.A.; Shields, A.T.; Piner, L.W.; Bales, C.W.; Kraus, W.E. Effects of aerobic and/or resistance training on body mass and fat mass in overweight or obese adults. J. Appl. Physiol. 2012, 113, 1831–1837. [Google Scholar] [CrossRef] [PubMed]
- Hafstad, A.D.; Lund, J.; Hadler-Olsen, E.; Höper, A.C.; Larsen, T.S.; Aasum, E. High-and moderate-intensity training normalizes ventricular function and mechanoenergetics in mice with diet-induced obesity. Diabetes 2013, 62, 2287–2294. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.M.; Bouchard, C.; Church, T.; Slentz, C.; Kraus, W.E.; Redman, L.M.; Heymsfield, S.B. Why do individuals not lose more weight from an exercise intervention at a defined dose? An energy balance analysis. Obes. Rev. 2012, 13, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Nuutila, P. Functional brown adipose tissue in healthy adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.C.; Huang, W.C.; Chiu, C.C.; Chang, Y.K.; Huang, C.C. Whey protein improves exercise performance and biochemical profiles in trained mice. Med. Sci. Sports Exerc. 2014, 46, 1517–1524. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Lin, C.I.; Chiu, C.C.; Lin, Y.T.; Huang, W.K.; Huang, H.Y.; Huang, C.C. Chicken essence improves exercise performance and ameliorates physical fatigue. Nutrients 2014, 6, 2681–2696. [Google Scholar] [CrossRef] [PubMed]
- Brooks, G.A. Intra-and extra-cellular lactate shuttles. Med. Sci. Sports Exerc. 2000, 32, 790–799. [Google Scholar] [CrossRef] [PubMed]
- Carvalho-Peixoto, J.; Alves, R.C.; Cameron, L.C. Glutamine and carbohydrate supplements reduce ammonemia increase during endurance field exercise. Appl. Physiol. Nutr. Metab. 2007, 32, 1186–1190. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.H.; Paik, I.Y.; Jacobs, K. Regulation of blood glucose homeostasis during prolonged exercise. Mol. Cells 2007, 23, 272–279. [Google Scholar] [PubMed]
- Hayashi, T.; Hirshman, M.F.; Kurth, E.J.; Winder, W.W.; Goodyear, L.J. Evidence for 5′ AMP-activated protein kinase mediation of the effect of muscle contraction on glucose transport. Diabetes 1998, 47, 1369–1373. [Google Scholar]
- Klieverik, L.P.; Janssen, S.F.; van Riel, A.; Foppen, E.; Bisschop, P.H.; Serlie, M.J.; Kalsbeek, A. Thyroid hormone modulates glucose production via a sympathetic pathway from the hypothalamic paraventricular nucleus to the liver. Proc. Natl. Acad. Sci. USA 2009, 106, 5966–5971. [Google Scholar] [CrossRef] [PubMed]
- Langenberg, C.; Wan, L.; Egi, M.; May, C.N.; Bellomo, R. Renal blood flow in experimental septic acute renal failure. Kidney Int. 2006, 69, 1996–2002. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Shieh, M.J.; Kuo, S.L.; Lee, C.L.; Pan, T.M. Effect of red mold rice on antifatigue and exercise-related changes in lipid peroxidation in endurance exercise. Appl. Microbiol. Biotechnol. 2006, 70, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Sahlin, K.; Tonkonogi, M.; Söderlund, K. Energy supply and muscle fatigue in humans. Acta Physiol. Scand. 1998, 162, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, C.C.; Jurczak, M.J.; Danos, A.M.; Brady, M.J. Glycogen branches out: New perspectives on the role of glycogen metabolism in the integration of metabolic pathways. Am. J. Physiol. Endocrinol. Metab. 2006, 291, E1–E8. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Hawley, J.A.; Wong, S.H.; Jeukendrup, A.E. Carbohydrates for training and competition. J. Sports Sci. 2011, 29, S17–S27. [Google Scholar] [CrossRef] [PubMed]
- Bernard, J.R.; Liao, Y.H.; Doerner, P.G.; Ding, Z.; Hsieh, M.; Wang, W.; Ivy, J.L. An amino acid mixture is essential to optimize insulin-stimulated glucose uptake and GLUT4 translocation in perfused rodent hindlimb muscle. J. Appl. Physiol. 2012, 113, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.J.C.; Goodpaster, B.H. Increased intramuscular lipid storage in the insulin-resistant and endurance-trained state. Pflugers Arch. 2006, 451, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Chakravarthy, M.V.; Booth, F.W. Eating, exercise, and “thrifty” genotypes: Connecting the dots toward an evolutionary understanding of modern chronic diseases. J. Appl. Physiol. 2004, 96, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, H.; Kawada, N.; Kaimori, J.Y.; Kitamura, H.; Moriyama, T.; Rakugi, H.; Takahara, S.; Isaka, Y. Febuxostat suppressed renal ischemia-reperfusion injury via reduced oxidative stress. Biochem. Biophys. Res. Commun. 2012, 427, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Hamden, K.; Boujbiha, M.A.; Masmoudi, H.; Ayadi, F.M.; Jamoussi, K.; Elfeki, A. Combined vitamins (C and E) and insulin improve oxidative stress and pancreatic and hepatic injury in alloxan diabetic rats. Biomed. Pharmacother. 2009, 63, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, M.; González-Izal, M.; Navarro-Amezqueta, I.; Calbet, J.A.; Ibañez, J.; Malanda, A.; Mallor, F.; Häkkinen, K.; Kraemer, W.J.; Gorostiaga, E.M. Effects of strength training on muscle fatigue mapping from surface EMG and blood metabolites. Med. Sci. Sports Exerc. 2011, 43, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Owen, O.E.; Kalhan, S.C.; Hanson, R.W. The key role of anaplerosis and cataplerosis for citric acid cycle function. J. Biol. Chem. 2002, 277, 30409–30412. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, J.T.; Brosnan, M.E. Branched-chain amino acids: Enzyme and substrate regulation. J. Nutr. 2006, 136, 207S–211S. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrition Facts | Content |
|---|---|
| Nutrition Facts | /100 g SP |
| Protein | 30.2 g |
| Fat | 2.8 g |
| Saturated fat | 1.36 g |
| Trans fat | 0 |
| Carbohydrate | 58.3 g |
| Sugar | 4 g |
| Moisture | 7.9 g |
| Ash | 0.8 g |
| Sodium | 11.8 mg |
| Total calories | 379.2 Kcal |
| Hydrolyzed amino acid profiles | g/100g |
| Serine | 1.32 |
| Aspartic Acid | 2.07 |
| Glutamic Acid | 3.95 |
| Glycine | 1.26 |
| Histidine | 0.67 |
| Arginine | 1.90 |
| Threonine | 1.12 |
| Alanine | 1.53 |
| Proline | 1.22 |
| Cystine | 0.59 |
| Tyrosine | 1.22 |
| Phenylalanine | 1.39 |
| Methionine | 0.63 |
| Lysine | 0.99 |
| Tryptophan | 0.37 |
| Valine | 1.73 |
| Isoleucine | 1.30 |
| Leucine | 2.25 |
| Characteristic | SC | PET | PET + SP | p-value between PET & SC | p-value between PET + SP & SC |
|---|---|---|---|---|---|
| Initial BW (g) | 30.7 ± 0.7 | 30.7 ± 0.7 | 30.7 ± 0.6 | 0.8594 | 0.9435 |
| Final BW (g) | 37.1 ± 2.1 | 36.8 ± 1.2 | 37.6 ±1.4 | 0.7197 | 0.5039 |
| Food intake (g/day) | 6.3 ± 0.5 | 6.2 ± 0.8 | 6.3 ± 0.8 | 0.5911 | 0.8673 |
| Water intake (mL/day) | 7.9 ± 1.5 | 7.6 ± 1.0 | 7.9 ± 1.3 | 0.3929 | 0.9326 |
| Weight (g) | |||||
| Liver | 1.88 ± 0.16 | 1.87 ± 0.06 | 1.89 ± 0.09 | 0.7745 | 0.9297 |
| Kidney | 0.58 ± 0.04 | 0.58 ± 0.03 | 0.58 ± 0.03 | 0.1000 | 0.9407 |
| Heart | 0.26 ± 0.04 | 0.25 ± 0.05 | 0.26 ± 0.04 | 0.6924 | 0.8210 |
| Lung | 0.25 ± 0.02 | 0.24 ± 0.02 | 0.24 ± 0.02 | 0.3043 | 0.3673 |
| Muscle | 0.39 ± 0.02 | 0.39 ± 0.02 | 0.39 ± 0.02 | 0.9183 | 0.6823 |
| EFP | 0.50 ± 0.11 b | 0.34 ± 0.11 a | 0.34 ± 0.06 a | 0.0034 | 0.0025 |
| BAT | 0.123 ± 0.014 a | 0.139 ± 0.016 b | 0.135 ± 0.012 ab | 0.0326 | 0.0930 |
| Relative Weight (%) | |||||
| Liver | 5.08 ± 0.41 | 5.08 ± 0.23 | 5.02 ± 0.25 | 0.9741 | 0.6799 |
| Kidney | 1.57 ± 0.12 | 1.58 ± 0.11 | 1.54 ± 0.07 | 0.8485 | 0.5678 |
| Heart | 0.69 ± 0.11 | 0.67 ± 0.15 | 0.69 ± 0.10 | 0.7614 | 0.9838 |
| Lung | 0.67 ± 0.07 | 0.65 ± 0.04 | 0.64 ± 0.06 | 0.4179 | 0.2288 |
| Muscle | 1.05 ± 0.07 | 1.06 ± 0.10 | 1.04 ± 0.03 | 0.8129 | 0.8657 |
| EFP | 1.35 ± 0.29 b | 0.93 ± 0.29 a | 0.90 ± 0.19 a | 0.0042 | 0.0024 |
| BAT | 0.33 ± 0.04 a | 0.38 ± 0.04 b | 0.36 ± 0.04 a | 0.0578 | 0.2373 |
| Variable | SC | PET | PET+SP | p-value between PET & SC | p-value between PET+SP & SC |
|---|---|---|---|---|---|
| AST (U/L) | 94 ± 37 a | 156 ± 46 b | 125 ± 22 ab | 0.0027 | 0.1063 |
| ALT (U/L) | 62 ± 9 a | 95 ± 34 b | 59 ± 10 a | 0.0045 | 0.8160 |
| CK (U/L) | 413 ± 149 | 418 ± 205 | 412 ± 114 | 0.9582 | 0.9840 |
| Albumin (g/dL) | 3.81 ± 0.20 | 3.90 ± 0.32 | 3.90 ± 0.21 | 0.4910 | 0.4910 |
| TP (g/dL) | 5.2 ± 0.2 | 5.2 ± 0.2 | 5.2 ± 0.2 | 1.0000 | 0.7999 |
| BUN (mg/dL) | 21.1 ± 2.4 | 21.1 ± 3.3 | 21.5 ± 1.6 | 0.9922 | 0.7481 |
| Creatinine (mg/dL) | 0.27 ± 0.04 a | 0.35 ± 0.05 b | 0.25 ± 0.04 a | 0.0008 | 0.5291 |
| UA (mg/dL) | 1.4 ± 0.2 a | 1.8 ± 0.4 b | 1.7 ± 0.4 ab | 0.0180 | 0.0892 |
| Glucose (mg/dL) | 146 ± 13 | 143 ± 16 | 143 ± 16 | 0.6976 | 0.7216 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-M.; Lin, C.-L.; Wei, L.; Hsu, Y.-J.; Chen, K.-N.; Huang, C.-C.; Kao, C.-H. Sake Protein Supplementation Affects Exercise Performance and Biochemical Profiles in Power-Exercise-Trained Mice. Nutrients 2016, 8, 106. https://doi.org/10.3390/nu8020106
Chen Y-M, Lin C-L, Wei L, Hsu Y-J, Chen K-N, Huang C-C, Kao C-H. Sake Protein Supplementation Affects Exercise Performance and Biochemical Profiles in Power-Exercise-Trained Mice. Nutrients. 2016; 8(2):106. https://doi.org/10.3390/nu8020106
Chicago/Turabian StyleChen, Yi-Ming, Che-Li Lin, Li Wei, Yi-Ju Hsu, Kuan-Neng Chen, Chi-Chang Huang, and Chin-Hsung Kao. 2016. "Sake Protein Supplementation Affects Exercise Performance and Biochemical Profiles in Power-Exercise-Trained Mice" Nutrients 8, no. 2: 106. https://doi.org/10.3390/nu8020106
APA StyleChen, Y. -M., Lin, C. -L., Wei, L., Hsu, Y. -J., Chen, K. -N., Huang, C. -C., & Kao, C. -H. (2016). Sake Protein Supplementation Affects Exercise Performance and Biochemical Profiles in Power-Exercise-Trained Mice. Nutrients, 8(2), 106. https://doi.org/10.3390/nu8020106