Psoralea corylifolia L. Seed Extract Attenuates Nonalcoholic Fatty Liver Disease in High-Fat Diet-Induced Obese Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of PCS Extract
2.2. Animals
2.3. Measurement of Blood Glucose Levels
2.4. Glucose Tolerance Tests
2.5. Insulin Tolerance Tests
2.6. Measurement of Hemoglobin A1c (HbA1c) Levels
2.7. Serum Lipid Profile
2.8. Oil Red O Staining
2.9. Hematoxylin and Eosin Staining
2.10. Quantification of Liver Triglyceride Content
2.11. Quantitative Real-Time RT-PCR (qRT-PCR) Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward/Reverse Primers |
|---|---|
| cyclophilin | 5′- TGGAGAGCACCAAGACAGACA 5′-TGCCGGAGTCGACAATGAT |
| SREBP1c | 5′-TGCCATCGCCAAGGAGTAG 5′-GGCCCGGGAAGTCACTGT |
| SCD1 | 5′-CCGGAGACCCTTAGATCGA 5′-GCCTGTAAAAGATTTCTGCAAACC |
| FAS | 5′-GCTGCGGAAACTTCAGGAAAT 5′-AGAGACGTGTCACTCCTGGACTT |
| CPT1α | 5′-CAAAGATCAATCGGACCCTAGAC 5′-CGCCACTCACGATGTTCTTC |
| PGC1α | 5′-CACTGACAGATGGAGCCGTGA 5′-TGTTGGCTGGTGCCAGTAAGAG |
| IL1β | 5′-CTACAGGCTCCGAGATGAACAAC 5′-TCCATTGAGGTGGAGAGCTTTC |
| MCP1 | 5′-TTAAAAACCTGGATCGGAACCAA 5′-GCATTAGCTTCAGATTTACGGG |
| SOCS3 | 5′-TCCAGCATCTTTGTCGGAAGA 5′-CCAGGCAGCTGGGTCACTT |
2.12. Western Blotting
2.13. Statistical Analyses
3. Results
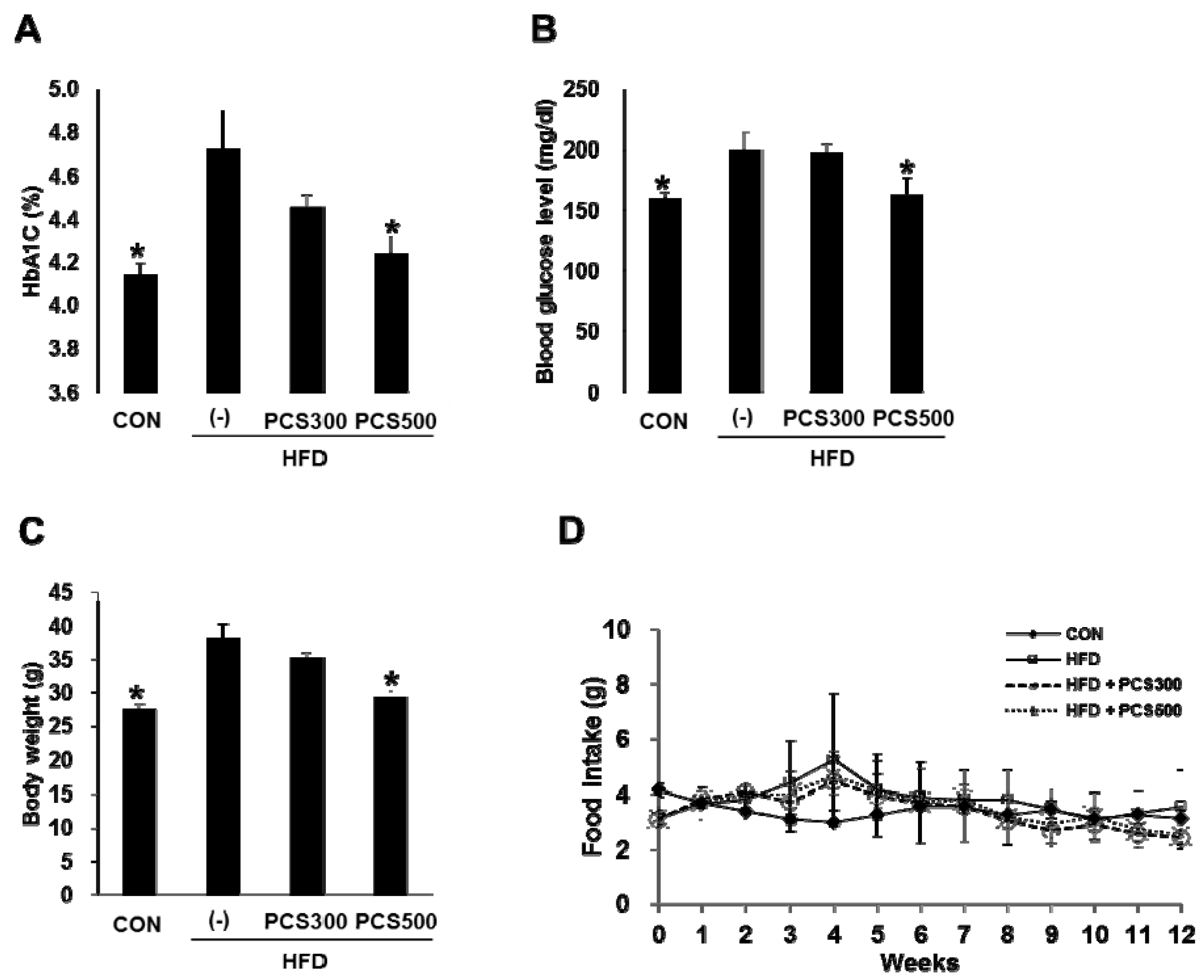
3.1. PCS Extract Treatment Decreased Hemoglobin A1c, Blood Glucose Levels, and Body Weight Gain in HFD-Fed Mice

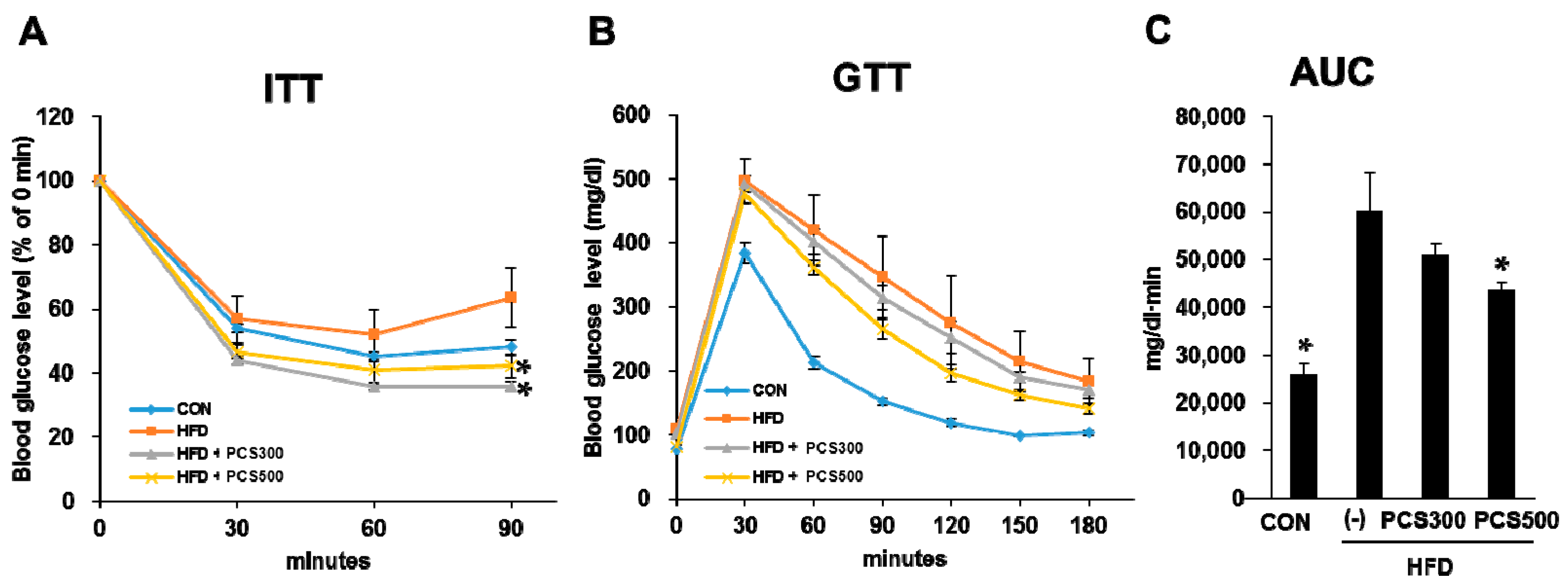
3.2. PCS Extract Treatment Improved Insulin Sensitivity and Glucose Tolerance in HFD-Fed Mice

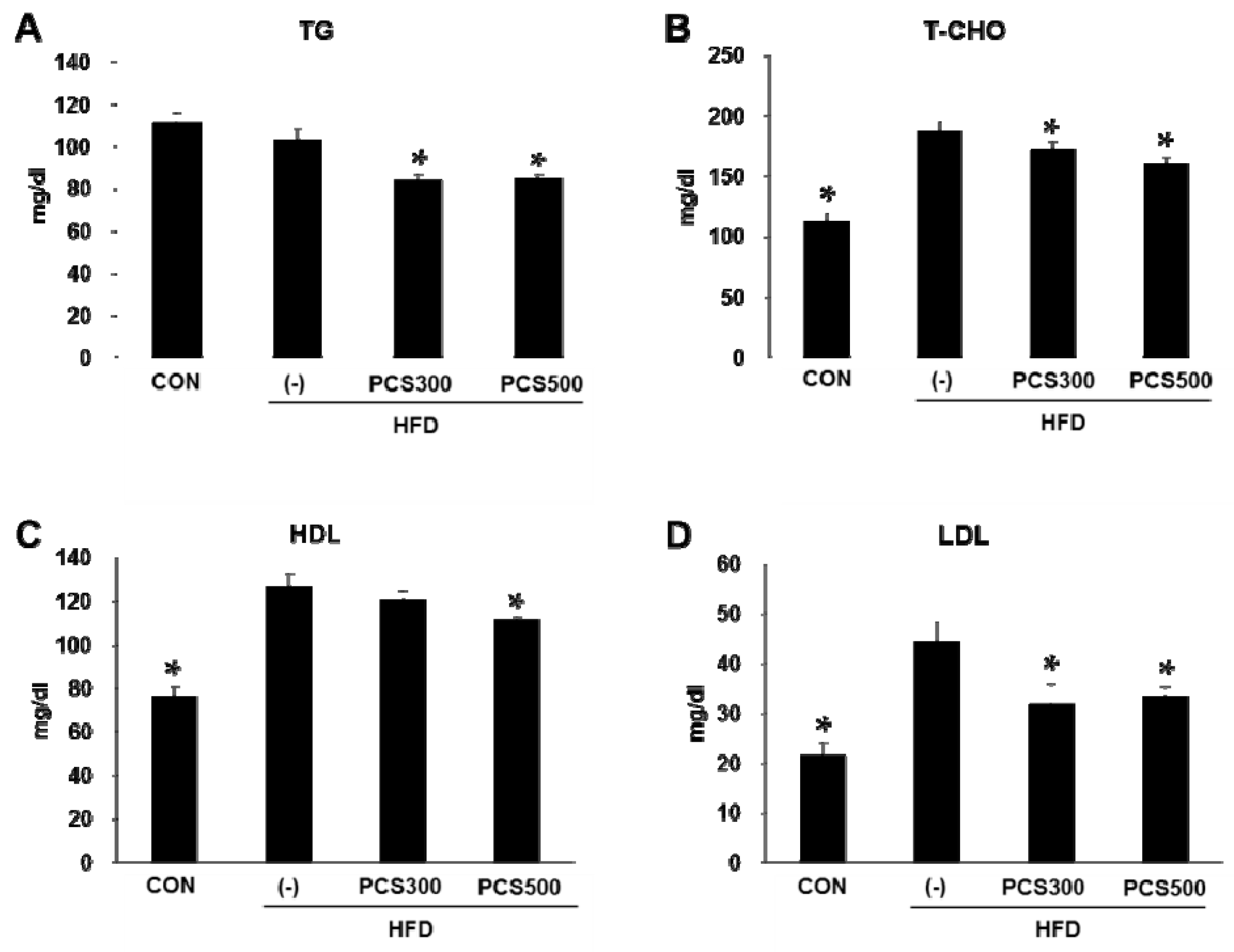
3.3. PCS Extract Treatment Decreased Plasma Lipid Profiles in HFD-Fed Mice

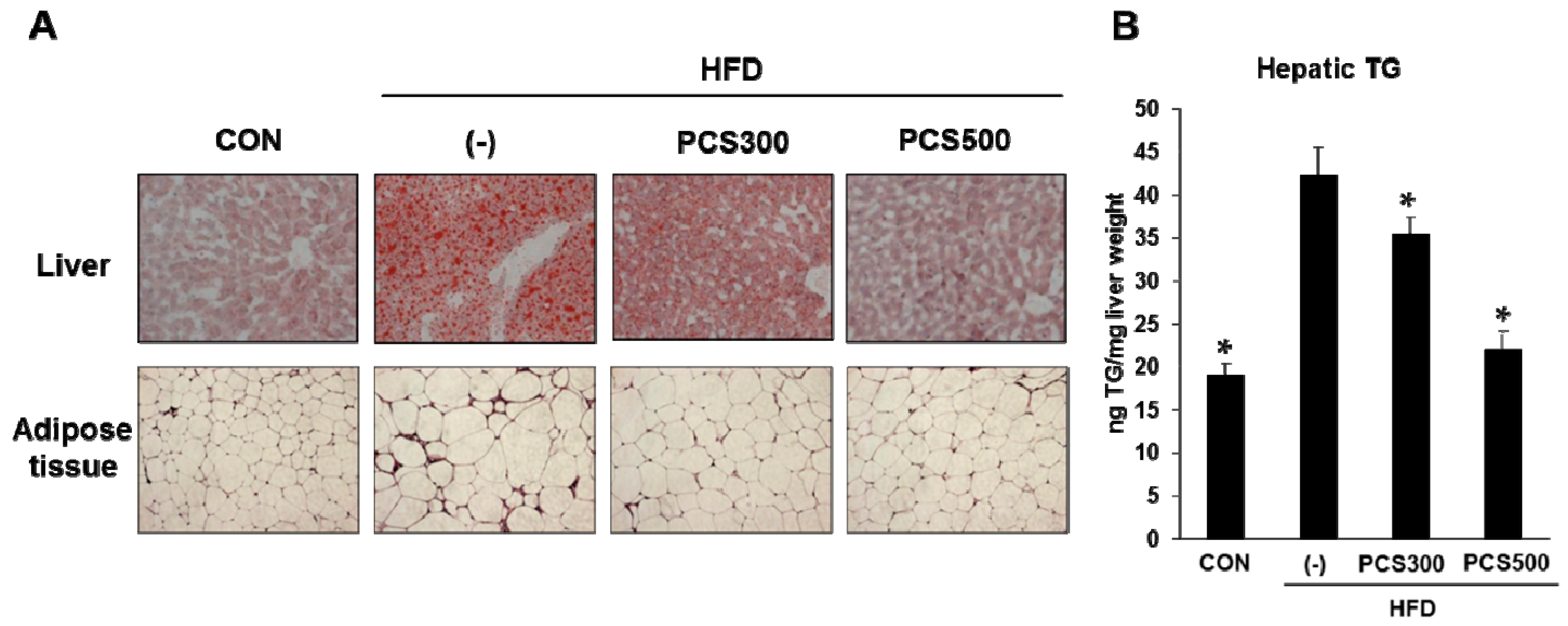
3.4. PCS Extract Treatment Decreased Lipid Accumulation in Liver and Adipose Tissue in HFD-Fed Mice

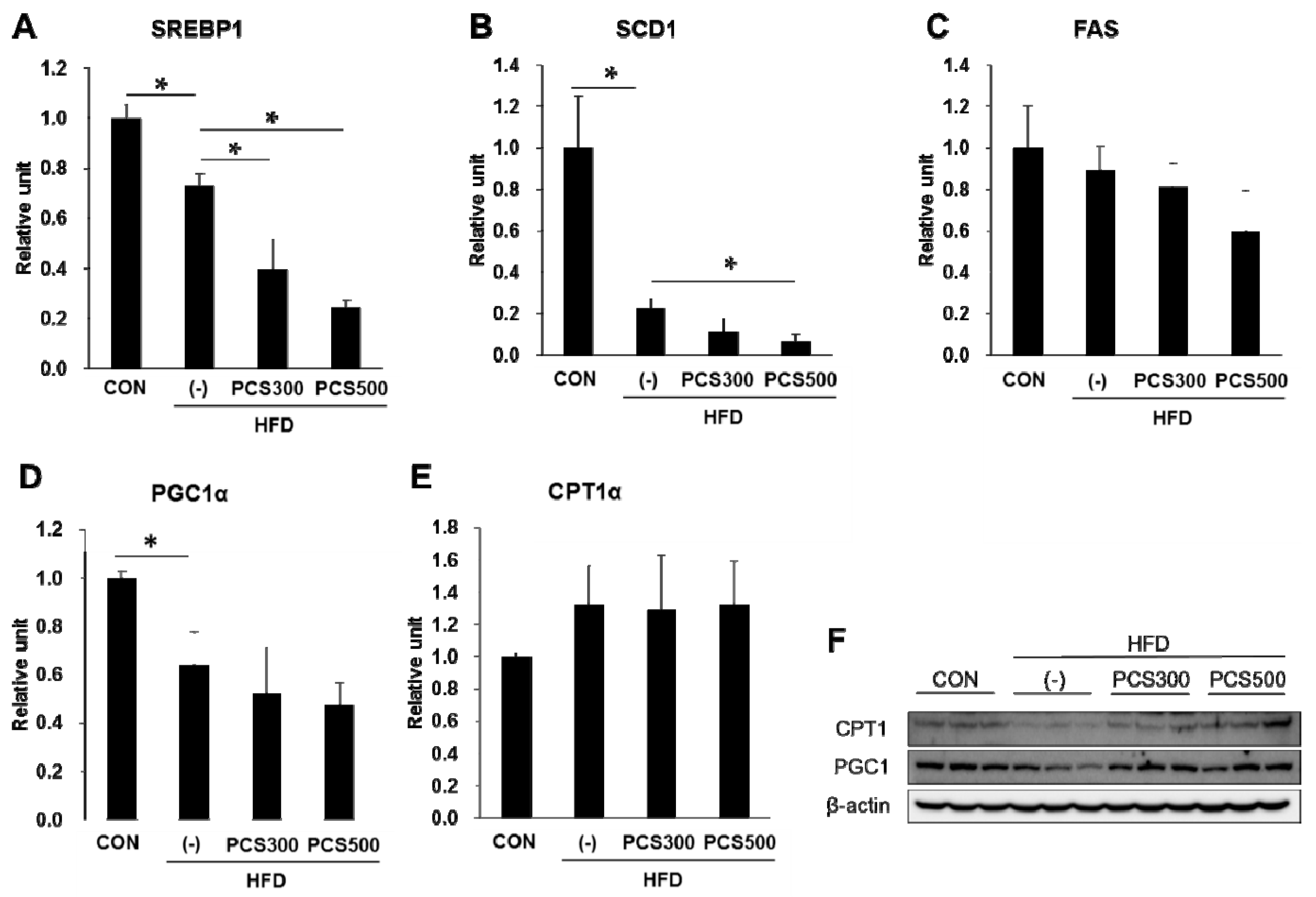
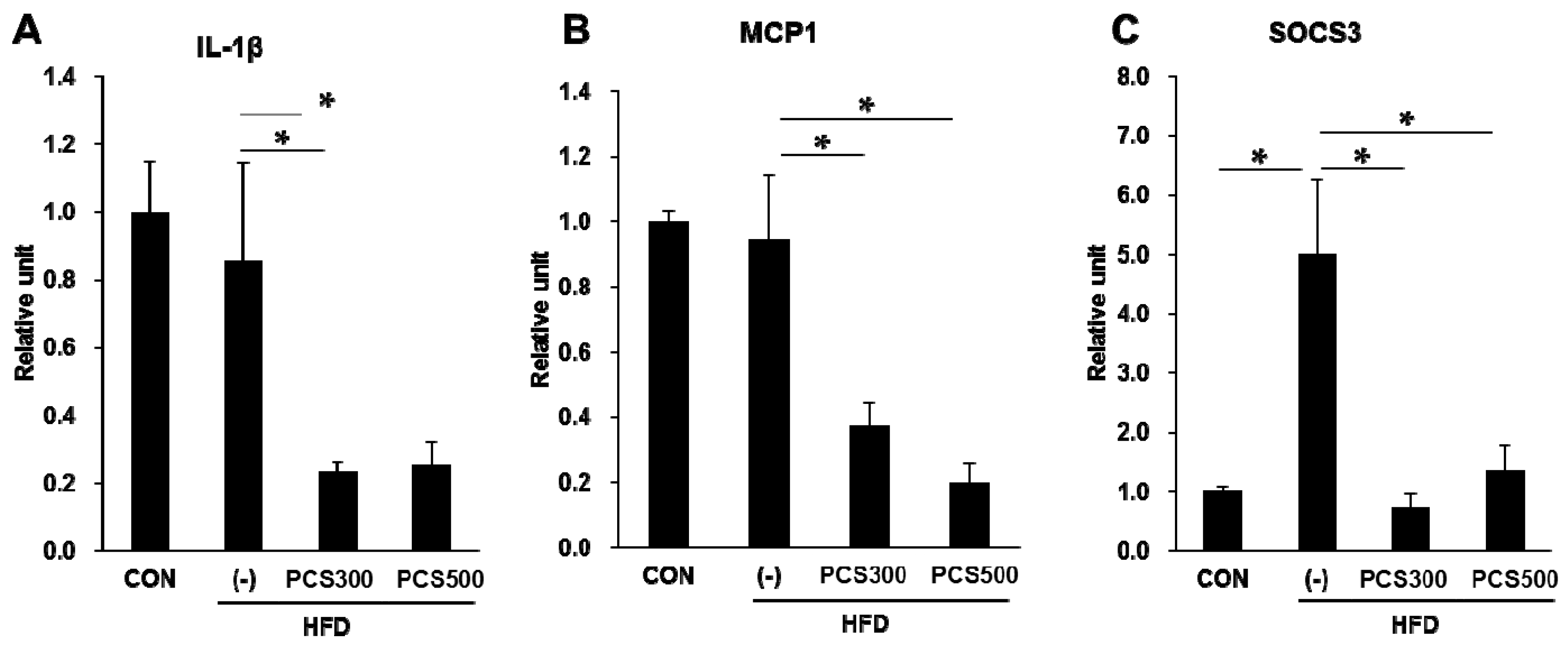
3.5. PCS Extract Treatment Decreased mRNA and Protein Expression of Genes for Lipid Metabolism and Hepatic Inflammation in Liver of HFD-Fed Mice


4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Misra, A.; Bhardwaj, S. Obesity and the metabolic syndrome in developing countries: Focus on south asians. Nestle Nutr. Inst. Workshop Ser. 2014, 78, 133–140. [Google Scholar] [PubMed]
- Smyth, S.; Heron, A. Diabetes and obesity: The twin epidemics. Nat. Med. 2006, 12, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Koo, S.H. Nonalcoholic fatty liver disease: Molecular mechanisms for the hepatic steatosis. Clin. Mol. Hepatol. 2013, 19, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Neves Ribeiro, D.; Goncalves Alfenas Rde, C.; Bressan, J.; Brunoro Costa, N.M. The effect of oilseed consumption on appetite and on the risk of developing type 2 diabetes mellitus. Nutr. Hosp. 2013, 28, 296–305. [Google Scholar] [PubMed]
- Lo, L.; McLennan, S.V.; Williams, P.F.; Bonner, J.; Chowdhury, S.; McCaughan, G.W.; Gorrell, M.D.; Yue, D.K.; Twigg, S.M. Diabetes is a progression factor for hepatic fibrosis in a high fat fed mouse obesity model of non-alcoholic steatohepatitis. J. Hepatol. 2011, 55, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Attar, B.M.; Van Thiel, D.H. Current concepts and management approaches in nonalcoholic fatty liver disease. Sci. World J. 2013, 2013, 481893. [Google Scholar] [CrossRef] [PubMed]
- Babu, P.V.; Liu, D.; Gilbert, E.R. Recent advances in understanding the anti-diabetic actions of dietary flavonoids. J. Nutr. Biochem. 2013, 24, 1777–1789. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Fai So, K.; Liong, E.C.; Tipoe, G.L. Recent advances in the herbal treatment of non-alcoholic fatty liver disease. J. Tradit. Complement. Med. 2013, 3, 88–94. [Google Scholar] [PubMed]
- Jiangning, G.; Xinchu, W.; Hou, W.; Qinghua, L.; Kaishun, B. Antioxidants from a chinese medicinal herb—Psoralea corylifolia L. Food Chem. 2005, 91, 287–292. [Google Scholar] [CrossRef]
- Cho, H.; Jun, J.Y.; Song, E.K.; Kang, K.H.; Baek, H.Y.; Ko, Y.S.; Kim, Y.C. Bakuchiol: A hepatoprotective compound of Psoralea corylifolia on tacrine-induced cytotoxicity in Hep G2 cells. Planta Med. 2001, 67, 750–751. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Zhao, Y.Z.; Kim, Y.C.; Sohn, D.H. Protective effect of (S)-bakuchiol from Psoralea corylifolia on rat liver injury in vitro and in vivo. Planta Med. 2005, 71, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Wu, H.; Weng, X.; Yan, J.; Bi, K. Studies on extraction and isolation of active constituents from psoralen corylifolia l. And the antitumor effect of the constituents in vitro. J. Chin. Med. Mater. 2003, 26, 185–187. (In Chinese) [Google Scholar]
- Khatune, N.A.; Islam, M.E.; Haque, M.E.; Khondkar, P.; Rahman, M.M. Antibacterial compounds from the seeds of Psoralea corylifolia. Fitoterapia 2004, 75, 228–230. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Fan, C.Q.; Wang, Y.; Dong, L.; Yue, J.M. Antibacterial prenylflavone derivatives from Psoralea corylifolia, and their structure-activity relationship study. Bioorg. Med. Chem. 2004, 12, 4387–4392. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hong, C.; Zhou, C.; Xu, D.; Qu, H.B. Screening antitumor compounds psoralen and isopsoralen from Psoralea corylifolia L. Seeds. Evid. Based Complement. Alternat. Med. 2011, 2011, 363052. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.R.; Chang, C.L.; Hsieh, P.Y.; Lin, L.W.; Ching, H. Psoralen and isopsoralen, two coumarins of psoraleae fructus, can alleviate scopolamine-induced amnesia in rats. Planta Med. 2007, 73, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Lee, E.K.; Lee, C.S.; Chun, K.H.; Lee, M.Y.; Jun, H.S. Psoralea corylifolia l. Seed extract ameliorates streptozotocin-induced diabetes in mice by inhibition of oxidative stress. Oxid. Med. Cell Longev. 2014, 2014, 897296. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Oh, Y.S.; Kim, D.; Lee, M.Y.; Chae, S.; Jun, H.S. Protective role of Psoralea corylifolia L. seed extract against hepatic mitochondrial dysfunction induced by oxidative stress or aging. Evid. Based Complement. Alternat. Med. 2013, 2013, 678028. [Google Scholar] [CrossRef] [PubMed]
- Rohlfing, C.L.; Little, R.R.; Wiedmeyer, H.M.; England, J.D.; Madsen, R.; Harris, M.I.; Flegal, K.M.; Eberhardt, M.S.; Goldstein, D.E. Use of GHB (HBA1C) in screening for undiagnosed diabetes in the U.S. population. Diabetes Care 2000, 23, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Toye, A.A.; Dumas, M.E.; Blancher, C.; Rothwell, A.R.; Fearnside, J.F.; Wilder, S.P.; Bihoreau, M.T.; Cloarec, O.; Azzouzi, I.; Young, S.; et al. Subtle metabolic and liver gene transcriptional changes underlie diet-induced fatty liver susceptibility in insulin-resistant mice. Diabetologia 2007, 50, 1867–1879. [Google Scholar] [CrossRef] [PubMed]
- Stojsavljevic, S.; Gomercic Palcic, M.; Virovic Jukic, L.; Smircic Duvnjak, L.; Duvnjak, M. Adipokines and proinflammatory cytokines, the key mediators in the pathogenesis of nonalcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 18070–18091. [Google Scholar] [CrossRef] [PubMed]
- Ravussin, E.; Smith, S.R. Increased fat intake, impaired fat oxidation, and failure of fat cell proliferation result in ectopic fat storage, insulin resistance, and type 2 diabetes mellitus. Ann. N.Y. Acad. Sci. 2002, 967, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; DiNatale, D.A.; Chung, M.Y.; Park, Y.K.; Lee, J.Y.; Koo, S.I.; O′Connor, M.; Manautou, J.E.; Bruno, R.S. Green tea extract attenuates hepatic steatosis by decreasing adipose lipogenesis and enhancing hepatic antioxidant defenses in OB/OB mice. J. Nutr. Biochem. 2011, 22, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Catalano, D.; Martines, G.F.; Tonzuso, A.; Pirri, C.; Trovato, F.M.; Trovato, G.M. Protective role of coffee in non-alcoholic fatty liver disease (NAFLD). Digest. Dis. Sci. 2010, 55, 3200–3206. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Lu, F.E.; Zhao, L. Chinese herbal medicine in the treatment of nonalcoholic fatty liver disease. Chin. J. Integrat. Med. 2012, 18, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Polyzos, S.A.; Kountouras, J.; Zavos, C. Nonalcoholic fatty liver disease: The pathogenetic roles of insulin resistance and adipocytokines. Curr. Mol. Med. 2009, 9, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Vanni, E.; Bugianesi, E.; Kotronen, A.; De Minicis, S.; Yki-Jarvinen, H.; Svegliati-Baroni, G. From the metabolic syndrome to nafld or vice versa? Dig. Liver Dis. 2010, 42, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Karagianni, P.; Talianidis, I. Transcription factor networks regulating hepatic fatty acid metabolism. Biochim. Biophys. Acta 2014, in press. [Google Scholar] [CrossRef] [PubMed]
- Guturu, P.; Duchini, A. Etiopathogenesis of nonalcoholic steatohepatitis: Role of obesity, insulin resistance and mechanisms of hepatotoxicity. Int. J. Hepatol. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.H.; Byrne, C.D. Modulation of sterol regulatory element binding proteins (SREBPS) as potential treatments for non-alcoholic fatty liver disease (NAFLD). Drug Discov Today 2007, 12, 740–747. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Z.; Berk, M.; McIntyre, T.M.; Feldstein, A.E. Hepatic lipid partitioning and liver damage in nonalcoholic fatty liver disease: Role of stearoyl-COA desaturase. J. Biol. Chem. 2009, 284, 5637–5644. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Sohn, I.; Ahn, J.I.; Lee, K.H.; Lee, Y.S.; Lee, Y.S. Hepatic gene expression profiles in a long-term high-fat diet-induced obesity mouse model. Gene 2004, 340, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Wang, M.; Lu, M.; Xi, B.; Sheng, H.; Zang, Y.Q. Rhein ameliorates fatty liver disease through negative energy balance, hepatic lipogenic regulation, and immunomodulation in diet-induced obese mice. Am. J. Physiol. Endocrino. Metabol. 2011, 300, E886–E893. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Ohtake, T.; Motomura, W.; Takahashi, N.; Hosoki, Y.; Miyoshi, S.; Suzuki, Y.; Saito, H.; Kohgo, Y.; Okumura, T. Increased expression of ppargamma in high fat diet-induced liver steatosis in mice. Biochem. Biophys. Res. Commu. 2005, 336, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hong, S.W.; Chae, S.W.; Kim, D.H.; Choi, J.H.; Bae, J.C.; Park, S.E.; Rhee, E.J.; Park, C.Y.; Oh, K.W.; et al. Exendin-4 improves steatohepatitis by increasing SIRT1 expression in high-fat diet-induced obese C57Bl/6J mice. PloS ONE 2012, 7, e31394. [Google Scholar] [CrossRef] [PubMed]
- McGarry, J.D.; Brown, N.F. The mitochondrial carnitine palmitoyltransferase system. From concept to molecular analysis. Eur. J. Biochem. 1997, 244, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Puigserver, P.; Andersson, U.; Zhang, C.; Adelmant, G.; Mootha, V.; Troy, A.; Cinti, S.; Lowell, B.; Scarpulla, R.C.; et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell 1999, 98, 115–124. [Google Scholar] [CrossRef]
- Pae, H.O.; Cho, H.; Oh, G.S.; Kim, N.Y.; Song, E.K.; Kim, Y.C.; Yun, Y.G.; Kang, C.L.; Kim, J.D.; Kim, J.M.; et al. Bakuchiol from psoralea corylifolia inhibits the expression of inducible nitric oxide synthase gene via the inactivation of nuclear transcription factor-kappab in raw 264.7 macrophages. Int. Immunopharmacol. 2001, 1, 1849–1855. [Google Scholar] [CrossRef]
- Ma, K.L.; Ruan, X.Z.; Powis, S.H.; Chen, Y.; Moorhead, J.F.; Varghese, Z. Inflammatory stress exacerbates lipid accumulation in hepatic cells and fatty livers of apolipoprotein e knockout mice. Hepatology 2008, 48, 770–781. [Google Scholar]
- Zhao, L.; Zhong, S.; Qu, H.; Xie, Y.; Cao, Z.; Li, Q.; Yang, P.; Varghese, Z.; Moorhead, J.F.; Chen, Y.; et al. Chronic inflammation aggravates metabolic disorders of hepatic fatty acids in high-fat diet-induced obese mice. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, E.; Oh, Y.S.; Jun, H.-S. Psoralea corylifolia L. Seed Extract Attenuates Nonalcoholic Fatty Liver Disease in High-Fat Diet-Induced Obese Mice. Nutrients 2016, 8, 83. https://doi.org/10.3390/nu8020083
Seo E, Oh YS, Jun H-S. Psoralea corylifolia L. Seed Extract Attenuates Nonalcoholic Fatty Liver Disease in High-Fat Diet-Induced Obese Mice. Nutrients. 2016; 8(2):83. https://doi.org/10.3390/nu8020083
Chicago/Turabian StyleSeo, Eunhui, Yoon Sin Oh, and Hee-Sook Jun. 2016. "Psoralea corylifolia L. Seed Extract Attenuates Nonalcoholic Fatty Liver Disease in High-Fat Diet-Induced Obese Mice" Nutrients 8, no. 2: 83. https://doi.org/10.3390/nu8020083
APA StyleSeo, E., Oh, Y. S., & Jun, H. -S. (2016). Psoralea corylifolia L. Seed Extract Attenuates Nonalcoholic Fatty Liver Disease in High-Fat Diet-Induced Obese Mice. Nutrients, 8(2), 83. https://doi.org/10.3390/nu8020083