1. Introduction
On average, Americans consume 0.43 gallons per capita of grape juice (GJ) annually, making GJ the third most commonly consumed juice in the US [
1]. Native American Concord and Niagara grape cultivars are sources for production of purple and white juice, respectively. Both grapes and their corresponding juices are well established sources of nutrients and bioactive phenolic compounds, including flavan-3-ols, flavonols, stilbenes, phenolic acids and, for Concord grapes, anthocyanins [
2,
3,
4,
5]. With total phenolic and anthocyanin levels reported as high as 2900 mg/L and 880 mg/L, respectively, for 100% Concord GJ and similarly high levels of phenolics in 100% Niagara GJ, these products can be significant contributors to health promoting phytochemicals [
4].
Phenolic rich 100% Concord GJ consumption has been reported to have health promoting activities including improved cardiovascular and cognitive function [
6,
7,
8]. Primary outcomes mediated by Concord GJ include increased flow mediated dilation, decreased platelet aggregation, modulation of low density lipoprotein (LDL) oxidation lag time, and improved memory function and brain signaling (reviewed by Blumberg et al. [
6]; Krikorian et al. [
7,
8]). While promising, these benefits have been observed with consumption of between ~12 and 20 oz of 100% juice and 100% GJ contains ~36 g sugar per 240 mL serving [
9]. While fruit juices have been reported to have similar glycemic response to whole fruits when matched by sugar load [
10], realization of these benefits remains challenged by consumer concern related to the higher sugar content and risk for obesity and diabetes. Therefore, while evidence for the benefits of GJ and GJ phenolics continues to expand, there remains a hesitation in recommending 100% GJ consumption to certain populations due to its natural high sugar content.
Over the past decade, the potential of phenolics to modulate glucose homeostasis has emerged (reviewed by Hanhineva et al. [
11] and Williamson et al. [
12]). Specifically, phenolics derived from foods including berries, juices, tea and coffee have demonstrated the ability to modulate intestinal digestion of starch by inhibition of amylase and glucosidase enzymes as well as intestinal glucose absorption through inhibition of glucose transporters such as GLUT 2 [
13,
14,
15,
16,
17,
18,
19,
20]. While primarily based on experiments with purified phenolics and phenolic extracts, these data would suggest that sugar in the context of a phenolic rich food or beverage may be processed differently in the intestine resulting in a modified glycemic response. Interestingly, while similar glycemic responses have been observed between grape juice and fruit, juice resulted in lower insulin response [
21]. Johnston et al. [
22] previously reported that 3 h glycemic response from both clear and cloudy 100% apple juice was in fact lower in healthy volunteers compared to a phenolic-free, sugar-matched placebo beverage. While no mechanistic test were performed, the authors postulated that phenolics in apple juices, including phloridzin or other polyphenols, may be responsible, in part, for the observed delay in intestinal absorption of glucose. In a related fashion, chronic consumption of 100% Concord GJ (8 weeks) decreased fasting blood glucose levels compared to placebo beverage [
23]. These results do in fact suggest that specific 100% juice components might modify glucose absorption and/or homeostasis in humans.
While these data are promising, additional insight into the ability and mechanisms by which GJ phenolics may modulate glucose absorption is required. Also, considering the potential mechanism of phenolic inhibition of starch digestive enzyme and glucose transport, it is important to consider the potential impact of a complex meal on these effects. Although clinical studies remain the gold standard for investigating health-related outcomes, in vitro models provide an effective screening tool to investigate mechanistic steps and screen a broader set of food matrix factors that impact nutrient and phytonutrient bioavailability prior to refining designs for clinical evaluation [
24,
25,
26,
27,
28]. Leveraging a similar approach, the specific objectives of this study were to (1) examine the potential for 100% GJ phenolics to modulate carbohydrate digestion and intestinal glucose transport in vitro; and (2) determine if bioaccessible phenolics from 100% GJ can alter carbohydrate digestion and intestinal glucose transport in the presence of a starch rich meal using a coupled in vitro digestion/Caco-2 model.
2. Materials and Methods
2.1. Chemicals, Solutions and Standards
Chromatography solvents, acids and salts including acetonitrile, methanol and water, formic acid and ammonium formate in addition to phenolic standards (gallic acid, caffeic acid, epicatechin, quercetin-3-O-glucoside, quercetin-3-glucuronide, quercetin, resveratrol, and cyanidin-3-O-glucoside) were purchased from Sigma-Aldrich, (St. Louis, MO, USA). Material for test meal including nonfat dry milk (NFDM; Maple Island, North St. Paul, MN, USA) and corn starch (Tate and Lyle) were purchased at a local market. Materials for in vitro digestion including urea (U5378), uric acid (U2625), porcine mucin (M2378), α-amylase (A3176), pepsin (P7125), lipase (L3126), pancreatin (P1750) bile salt (B8631) extract, KH2PO4 (VWR), K2SO4 (Riedel-de Haën), potassium citrate, sodium citrate, KCl, CaCO3, MgCO3 (Sigma-Aldrich) and Tris-HCl were also purchased from Sigma-Aldrich. NaCl, HCl, and NaHCO3 were purchased from Mallinckrodt (Phillipsburg, NJ, USA). Reagents for enzyme inhibition assays including dimethylsulfoxide (DMSO), acarbose, phosphoric acid, α-amylase (A3176), rat intestinal α-glucosidase, NaCl, glucose oxidase-peroxidase, maltose, maltotriose, maltotetrose and maltopentose, were obtained from Sigma-Aldrich. Cell culture reagents including Dulbecco’s Modified Eagles Medium (DMEM), non-essential amino acids (NEAA), penicillin/streptomycin (pen/strep), and phosphate buffered saline (PBS) were purchased from Lonza (Walkersville, MD, USA). Cell culture reagents including 4-(2-hydroxylethyl)-1-piperazineethanes (HEPES), bovine serum albumin (free fatty acid free) (FFA) and glucose-free DMEM were purchased from Sigma-Aldrich. NaHCO3, monosodium phosphate, and disodium phosphate were obtained from J.T. Baker (Center Valley, PA, USA). Fetal bovine serum (FBS) (Atlanta Biologicals, Lawrenceville, GA, USA), gentamycin (J.R. Scientific, Woodland, CA, USA), trypsin (Thermo Scientific, Waltham, MA, USA), glucose and fructose (Research Products International Corps, Mt. Prospect, IL, USA), and d-glucose-1,2,3,4,5,6,6-d7 (d7-glu) and d-Fructose-1,1,3,4,5,6,6-d7 (d7-fru) (Sigma-Aldrich) were used in glucose transport experiments.
2.2. Grape Juice Samples
One hundred percent Niagara and Concord GJ were provided by Welch Foods Inc. (Concord, MA, USA) (
Table 1). One hundred percent juices were produced from two harvest years (2013 and 2014) and were pasteurized and maintained refrigerated at 4 °C until used in experiments. The Niagara GJs were processed with and without the addition of sulfur dioxide.
2.3. Phenolic Extraction
Phenolics were extracted from aliquots (5 mL) of GJ by solid phase extraction (Oasis
® HLB 6cc (150 mg) extraction cartridges) using the method of Song et al. [
29]. Briefly, 5 mL of juice were loaded onto the SPE cartridges (Milford, MA, USA) and rinsed with 2% formic acid in water. Elution of phenolics was completed with 2% formic acid in methanol. Eluates were dried down under nitrogen and kept frozen (−80 °C) until analysis. Total phenolic content of extracts was measured using a modified Folin-Ciocalteau assay as described by Waterhouse et al. [
30].
2.4. Analysis of Polyphenol and Anthocyanin-Rich Fractions by LC-MS
Dried GJ extracts were resolubilized in 2.0% formic acid in water and characterized by LC-MS using methods of Song et al. [
29] with minor modification. Single ion responses (SIRs) were used to quantify individual GJ phenolics. Phenolic acids, flavonoids and stilbenoids were quantified using multi-level response curves constructed with authentic standards of each phenolic species identified by co-chromatography with limited exception. Piceid concentration (a resveratrol glucoside) was estimated using a resveratrol calibration curve. Concentration of all quercetin-
O-glucosides was estimated using quercetin-3-
O-glucoside. Finally, concentrations of anthocyanins were determined using a calibration curve constructed from cyanidin-3-
O-glucoside.
2.5. Impact of GJ Phenolic Extracts on Starch Digestive Enzymes in Vitro
Impact of GJ phenolic extracts on starch digestion by α-amylase and α-glucosidase was determined as described by Lee et al. [
31]. Briefly, GJ phenolic extracts were dissolved in DMSO (5 mM). A waxy maize starch solution (1 g/50 mL) was prepared in 20 mM sodium phosphate buffer and boiled to achieve gelatinization (20 min). Starch solutions (50 µL) were then combined with GJ phenolic extract (15 µL, delivering 10–500 μM of total phenols), pancreatic α-amylase (37 °C, 10 U/37.5 µL) and phosphate buffer (20 mM, pH 6.8). Samples were incubated for 10 min after which the reaction was terminated by boiling. Samples were diluted 10× in water prior to quantification of maltose and maltotriose by HPAEC-ECD. Percent inhibition of α-amylase by GJ phenolics was calculated relative to vehicle control (DMSO with no GJ extract) and compared to positive control acarbose.
Inhibition of α-glucosidase by GJ phenolics was assessed using a Megazyme glucose assay kit (Megazyme Inc., Chicago, IL, USA). Briefly, rat intestinal α-glucosidase solution (1 g/10 mL, 10 µL) was mixed with inhibitor (10 µL, 100–5000 µM total polyphenols). Sodium phosphate (0.1 M, pH 6.8, 70 µL) was added to the enzyme-inhibitor solution and the mixture was vortexed well. Maltose solution (10 µL, 100 mg/mL) was then added. The reaction mixture was incubated at 37 °C for 90 min. Enzymes were inactivated by placing the samples in boiling water. Samples were then centrifuged. The supernatant was collected and diluted 10×. Glucose content was determined by addition of glucose oxidase-peroxidase reagent and measuring absorbance at 510 nm. Percent inhibition of α-glucosidase by phenolic extracts and acarbose was determined by comparing the difference in absorbance between control and extract relative to absorbance of the control.
2.6. Inhibition of Glucose/Fructose Transport through Caco-2 Human Intestinal Cell Monolayers by GJ Phenolic Extracts
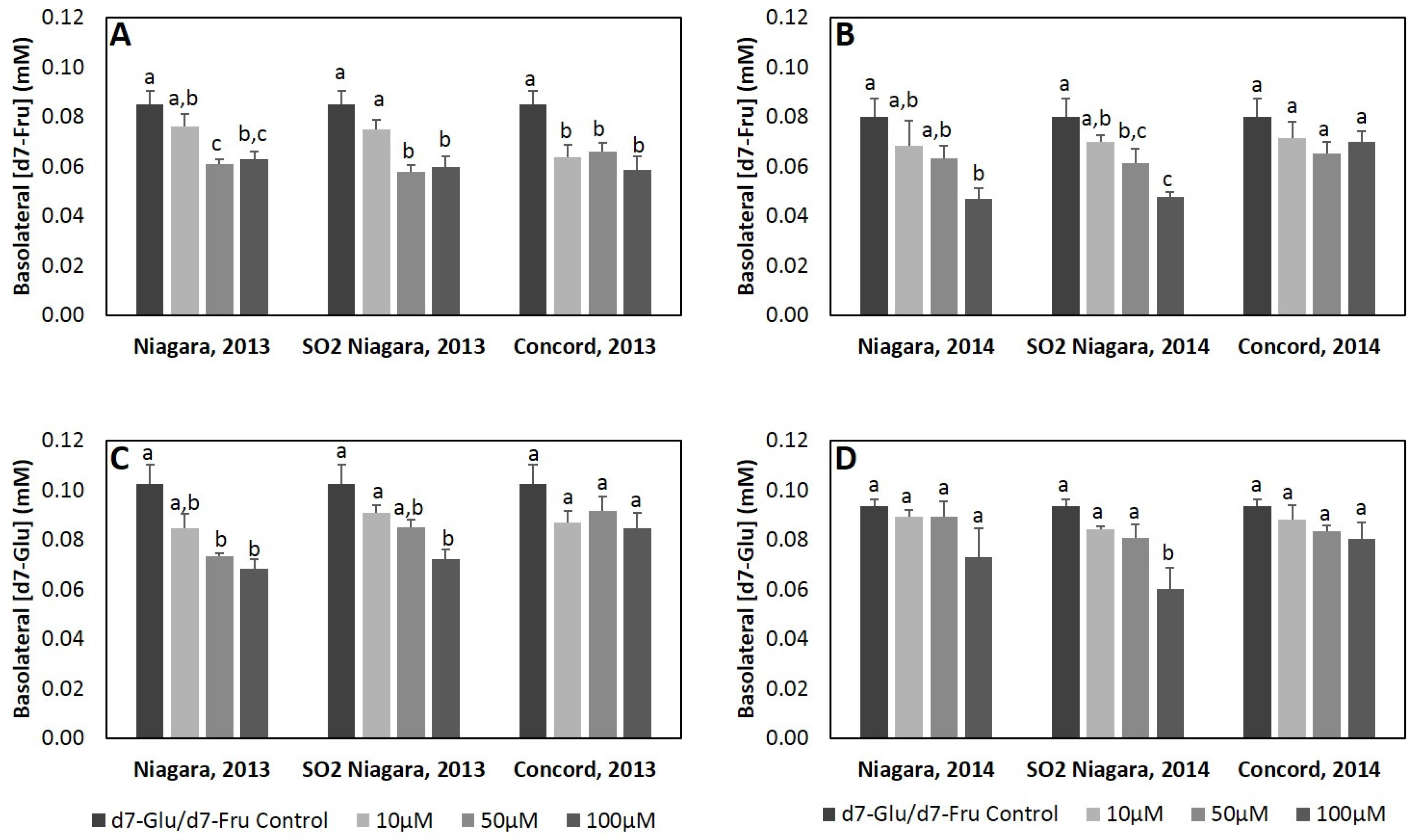
Potential for inhibition of glucose and fructose intestinal transport by GJ phenolic extract was assessed using a three-compartment Caco-2 human intestinal cell culture model. Caco-2 (TC7 clone) cells were maintained in DMEM supplemented with 10% v/v FBS, 1% v/v NEAA, 1% v/v HEPES, 1% v/v pen/strep and 0.1% v/v gentamicin. Cells were seeded (2.12 × 105 cells/cm2), grown and differentiated in 6 well insert plates (Corning® Transwell® polyester membrane, Corning Inc., Oneonta, AL, USA, 24 mm diameter, pore size 0.4 μm) under a humidified atmosphere of air/CO2 (95:5) at 37 °C. All experiments used differentiated monolayers (electrical resistance >250 Ω) at passages 90–95, with transport studies conducted 21–24 days post-confluency. Cells were incubated in glucose-free DMEM for 2 h preceding treatment. Test media for initial experiments was prepared by solubilizing glucose and fructose (9 mM each), d7-glu and d7-fru (3 mM each), and GJ phenolic extracts in PBS pH 5.5 (delivering 10–100 µM total phenolics, respectively). Cellular viability was assessed using the MTT assay (Biotium, Hayward, CA, USA). Highly differentiated cell monolayers treated with phenolic extracts and digesta (at concentrations >100 µM) for 4 h were found to have >95% viability. Test media was applied to the apical surface of cell monolayers. After 60 min incubation, basolateral and apical media were collected and cells were washed twice with 0.1% fatty acid free PBS. Membranes were then washed with ice cold PBS to terminate glucose transport, and cells were collected by scraping and frozen until analysis. All treatments were performed in quadruplicate.
2.7. Analysis of d7-Glucose and d7-Fructose Concentration in Basolateral Media by LC-MS
Basolateral media (100 µL) was extracted using acetone (0.5 mL), dried down under nitrogen, resolubilized in mobile phase (0.6 mL), and centrifuged (14,000 rpm, 5 min) prior to analysis for the chlorine adduct of d7-glu and d7-fru by LC-TOF-MS [
32]. 10 μL of sample was injected on a Waters ACQUITY UPLC H-Class system equipped with an ACQUITY QDa mass detector. Separation was achieved according to a method by Liu et al. [
33] with minor modification. A Waters BEH-amide column (2.1 mm id × 150 mm, 2.5 μM particle size) was heated to 30 °C under isocratic conditions with flow rate of 0.65 mL/min for 6 min and mobile phase 87:13 acetonitrile:water with 8 mM ammonium formate, pH 9.8. Conditions were as follows: ionization mode: ESI (−); mass: 222
m/
z; capillary voltage: 0.8 kV; cone voltage: 20 V; probe temp: 350 °C; desolvation temp: 600 °C. Glucose, fructose, d7-glu and d7-fru concentrations were calculated using calibration curves made from authentic standards.
2.8. Impact of 100% GJ on Glucose Release/Transport in a Coupled in Vitro Digestion/Caco-2 Model
To extend beyond GJ extracts, a coupled three-stage in vitro digestion/Caco-2 model was used to determine if bioaccessible GJ phenolics could inhibit starch digestion and/or glucose transport with or without a co-consumed starch rich meal. Initially 100% GJ or sugar matched phenolic free control (~10 mL) was introduced to a three-stage in vitro digestion with oral, gastric and small intestinal phase as described by Moser et al. [
34] and modified according to conditions described by Vermeirssen et al. [
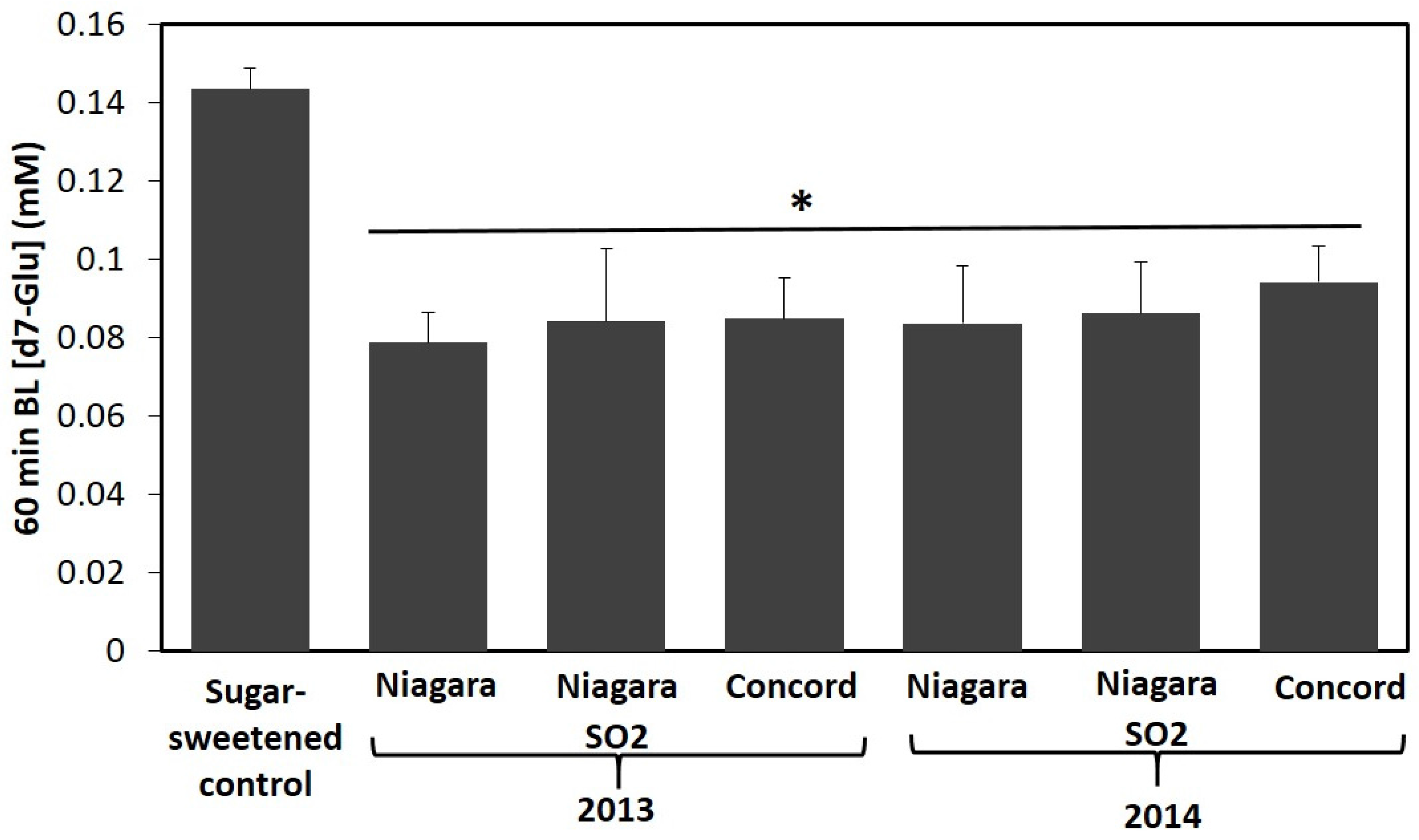
35] to include rat intestinal powder (0.15 g/reaction) as a source of α-glucosidase. Aliquots of undigested beverage starting material (SM), and centrifuged aqueous intestinal digesta (AQ) containing bioaccessible GJ phenolics were collected and acidified with aqueous acetic acid (1% total in sample) and stored frozen at −80 °C until phenolic analysis by HPLC-MS (outlined above). A separate aliquot of AQ digesta was then diluted 2:7 with PBS (pH 5.5) (delivering ~21–56 µM total bioaccessible GJ phenolics, ~24 mM monosaccharides), spiked with d7-glu (6 mM), and applied to the apical surface of Caco-2 monolayers. A matching phenolic-free control was prepared by solubilizing glucose and fructose (24 mM total) with d7-glu (6 mM each) in blank AQ digesta diluted 2:7 with PBS. Treatments were replicated in quadruplicate. Transport of d7-glu was followed as described previously.
In a second experiment, 100% GJ was co-digested with a test meal consisting of a starch/nonfat dry milk model meal. The model meal was prepared by mixing corn starch and non-fat dry milk (NFDM) in double-distilled water (10%
v/
v each). The mixture was then heated (95 °C, 30 min) and cooled slowly to 4 °C. The resulting product was blended and an aliquot (5 g) was combined with 2.5 or 5 g of 100% GJ (Concord or SO
2 Niagara, 2013 harvest) or sugar-matched control beverage (50:50 Glucose:Fructose; 16° Brix) prior to introduction to the oral phase of digestion. Starting materials (GJ plus model meal) and final AQ digesta were collected and stored (−80 °C). Bioaccessibility of phenolics was determined by comparing individual phenolic content of AQ digesta relative to starting material. The extent of starch digestion was determined by comparison of initial glucose content in starting material to that in final AQ digesta. Percentage inhibition of starch digestion by GJ was determined by comparing release of glucose from starch during digestion of model meal with grape juice relative to phenolic-free control. Following digestion, the ability of co-digested GJ to further inhibit glucose transport was determined by diluting AQ digesta 2:7 with PBS (pH 5.5) containing 6 mM d7-glu (delivering ~5–16 µM total bioaccessible phenolics, determined using Folin-Ciocalteu assay [
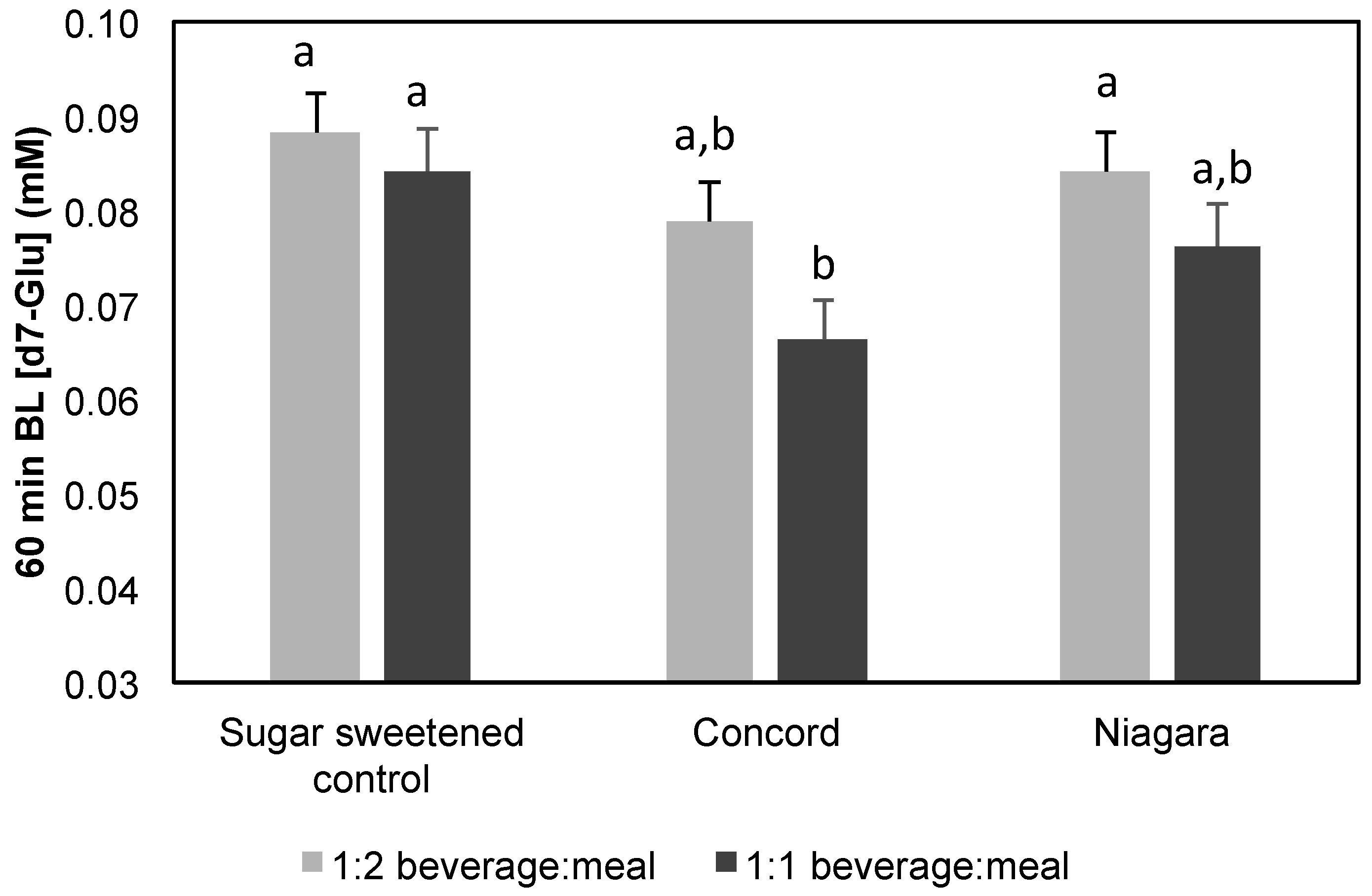
30]) and applying to the apical surface of Caco-2 monolayers. Feeding material containing AQ from high and low level GJ samples contained ~12 mM and 6 mM glucose and fructose. Matching phenolic-free controls were prepared. Treatments were replicated in quadruplicate. Transport of d7-glu (6 mM) was tracked and compared to control matched for sugar content.
2.9. Data Analysis
Data for polyphenol, anthocyanin, and (d7)-glu and (d7)-fru content of GJ samples, AQ digesta and basolateral material are expressed as mean ± SEM. Relative (%) bioaccessibility is defined as the percentage of polyphenol recovered in final digesta from that in starting material. Absolute bioaccessibility (μM) is the amount of phenolic available in digesta relative to that present in starting material, calculated by multiplying % bioaccessibility by concentration (μM) of phenolic in starting material. Percent (%) glucose release from corn starch by α-amylase was calculated as the fraction of glucose released compared to negative control. Glucose transport is expressed as concentration (μM) of d7-gluc appearing in the basolateral compartment over time. Percent (%) glucose transport was calculated on the basis of initial d7-glu content in the apical compartment at time 0. In order to facilitate comparison between treatments and control for variability between individual replicates, percentage (%) glucose transport was normalized using the d7-glu transport from control. Differences in phenolic profile, bioaccessibility data, enzyme inhibitory activity and glucose transport for each GJ or GJ extract were performed using JMP (Version 12, SAS Institute, Cary, NC, USA), and evaluated using Tukey’s test or t-test. All significant differences testing used α < 0.05.
4. Discussion
Clinical evidence exists to support the notion that certain phenolic-rich foods and beverages may modify glycemic parameters [
22,
23,
41,
42,
43,
44,
45,
46]. One hundred percent GJ is a particularly rich source of dietary phenolics but also naturally high in sugar (~36 g sugar per 240 mL serving) [
9]. While it has been reported that fruit juices have similar glycemic responses to their corresponding whole fruits [
10]), grape juice has also been shown to have a lower insulinemic response than corresponding grapes [
21]. This may be related to the differential profile of grape juice compared to grapes and, thus, a better understanding of the interaction between GJ phenolics and the intrinsic sugar in these products is required. This study was designed to develop insight into the ability and mechanisms by which GJ phenolics may modulate starch digestion and absorption of glucose in the context of a juice matrix with and without a starch rich meal.
One mechanism that has been proposed for these effects is related to phenolic inhibition of starch digestion [
47,
48,
49,
50]. In the present study, GJ phenolic extracts (300 and 500 µM) demonstrated only modest inhibitory capacity for α-amylase and α-glucosidase (
Table 3). Harvest year impacted α-amylase, but not α-glucosidase activity, with 2013 extracts generally having greater impact compared to 2014 extracts. This may be related to subtle differences in qualitative phenolic profiles from 2013 to 2014 harvest GJs as total levels appeared similar between harvest years. Phenolic rich GJ extracts had greater inhibitory activity toward α-glucosidase compared to α-amylase. Similar results were previously reported for wine grape tannins, pomace and skin extracts for inhibition of α-glucosidase (~20% to 85%), with little to no detectable impact on α-amylase activity [
48,
49]. Levels of phenolics required to achieve even modest inhibition were observed to be high (>300 µM). However, it is important to note that concentrations of phenolics in the gut lumen from a serving of phenolic rich food or beverage may in in fact be quite high and approach high µM to even mM levels as previously postulated [
7,
51]. Therefore, results here suggesting a modest ability of GJ phenolics to inhibit α-glucosidase at a 300 µM dose does support the hypothesis that consumption of GJ with starch rich foods may have relevance to starch digestion and liberation of glucose in the gut lumen.
A second mechanism by which phenolics may modulate glycemic response is through alteration of glucose intestinal transport [
18,
19,
20]. In the present study, GJ extracts (10–100 µM total phenolics) reduced intestinal glucose and fructose transport by Caco-2 human intestinal cell monolayers compared to control. These findings are similar to those previously reported with plant-derived phenolic extracts [
18,
19,
20] (
Figure 1;
Table S1). Overall, inhibition by GJ extracts was greater for d7-fru compared to d7-glu transport, with extracts from Niagara GJ generally exhibiting greater inhibitory activity relative to Concord GJ extracts. Since extracts were standardized for total phenolics, these results suggest that the qualitative phenolic profile of Niagara GJ, which is primarily composed of non-anthocyanin flavonoids and phenolic acids and minimal amounts of anthocyanins (
Table 2), may be most critical to consider in selection of juices and therefore merits additional investigation as targeted modifiers of intestinal glucose transport. The mechanism behind this reduction of glucose transport may be related, in part, to the ability of GJ phenolics to inhibit expression of hexose transporters (GLUT2 and SGLT1) or through direct inhibition of these transporters. Alzaid et al. [
20] demonstrated that GLUT2 and SGLT1 mRNA were significantly decreased compared to baseline by up to 85% and 70%, respectively, following treatment of cells with berry extract for over 12 h. However, although GLUT2 protein was significantly reduced compared to control by treatment of blueberry extract for 16 h, SGLT1 protein levels were not affected. In a preliminary experiment, expression of GLUT2 and SGLT1 mRNA was measured in Caco-2 monolayers exposed to 100% GJ phenolic extracts (SO
2 Niagara and Concord 2013) for 4 and 24 h. Interestingly, GLUT2 mRNA was significantly (
p < 0.05) decreased two-fold following treatment by Concord GJ phenolic extract, but no significant change in SGLT1 expression was observed (
Figure S1). While these preliminary results are consistent with previous observations [
18,
19,
20], they cannot fully explain the observed effects in the present study. Previous reports have also demonstrated that bioavailability of select polyphenols may be increased in the presence of carbohydrates [
52,
53,
54] suggesting the potential for additional interrelated mechanism impacting the transport of both phenolics and carbohydrate. Transepithelial flux of grape juice phenolics was not simultaneously monitored in this study, and, as such, it is not possible to speculate to these mechanisms. Further investigations are therefore warranted to better delineate the extent to which phenolic inhibit of natural juice sugars may be related to changes in transporter expression or function, or alternative mechanisms.
Finally, in order to better understand the extent to which these effects would be extendable to 100% GJ, Niagara and Concord GJ or matching phenolic free control were formulated with and without a starch rich model meal and digested in vitro. Differences observed in phenolic relative bioaccessibility from juice alone are suggestive of variations in grape phenolic components between harvest years (
Table 2 and
Table 4). On the other hand, SO
2 treatment did not impact relative bioaccessibility of phenolics from Niagara GJ. These results logically suggest that starting concentration of phenolics in 100% GJ have a direct impact on concentration of phenolics available for interactions in the gut and ultimate stability and accessibility of phenolics in the small intestine. Following assessment of phenolic bioaccessibility, transport of glucose from AQ digesta of 100% GJ and sugar match controls by Caco-2 intestinal cells was assessed. All GJ AQ digesta reduced d7-glu transport efficiency compared to phenolic-free sugar matched control up to 45% over 60 min (
Figure 2;
Table 5). These results were similar to those observed from extract screening and suggest that reduced efficiency of intestinal glucose transport may be a factor to consider in assessing glycemic response from GJ relative to a phenolic free sugar sweetened beverage. Interestingly, this observation is consistent with the glycemic response of phenolic rich apple juice that has previously been shown to cause a modest reduction in glycemic response relative to sugar matched controls in healthy humans [
22,
41].
To build on these findings with the awareness that GJ is commonly consumed with meals, it is important to consider the consequences of co-consumed food on ability of GJ phenolics to modulate carbohydrate digestion and glucose transport. It is well known that phenolics interact non-covalently with protein and carbohydrate (reviewed by Bordenave et al. [
55] and Jakobek et al. [
56]). These interactions in the context of a co-consumed meal may result in changes to the activity of phenolics relative to endpoints critical to glycemic response, namely carbohydrate digestion and glucose transport. Therefore, digestion of carbohydrate from a starch and protein-rich test meal co-formulated with GJ compared to phenolic-free control was determined. Although the level of GJ phenolics in AQ digesta resulting from co-digestion of GJ with test meal was lower compared to level of phenolics from GJ extracts used in enzyme inhibition assays (
Table 3 and
Table 5), results for inhibition of starch digestion were in fact similar. Therefore, it appears bioaccessible phenolics in the context of a complex meal still have the ability to impact digestion of carbohydrates derived from the meal. The extent to which this may be due to the proximity of phenolics and macronutrients in the meal and specific interactions that may develop through preparation and/or digestion remains to be explored.
Modifying glucose transport by GJ in the context of a digested meal was also determined. The effect of AQ digesta from GJ containing meals demonstrated only modest inhibitory effects (
Figure 3) which reach significance only for 100% Concord GJ. While in contrast with extract screening that found Niagara phenolics to be more effective, this observation was not totally unexpected as the phenolic concentrations resulting from digestion of mixed meals were lower than extract and juice experiments, especially for Niagara (
Table 5). Considering that in humans concentrations in the gut lumen may reach high μM to mM levels [
12] from typical food doses and gut dilution, responses within a meal such as those observed with berries by Törrönen et al. [
57] may be expected. While requiring additional clinical insights as to the direction and extent of this effect in vivo are required, current results, while in vitro, do reflect the modest but important changes in glycemic response observed in recent clinical trials involving phenolic-rich foods and beverages [
22,
23,
41,
42,
43,
44,
45,
46]. This is relevant considering that moderate post-prandial hyperglycemia blood glucose levels (148–199 mg/dL) have been shown to be indicative of the development of negative health effects including atherosclerosis and endothelial damage [
58]. This range represents a ~6% increase compared to the recommended <140 mg/dL post-prandial (2 h) blood glucose target [
59], suggesting that even subtle improvements to post-prandial blood glucose level may prevent development of negative health outcomes. Therefore, while subtle, the current observations that 100% GJ may impact both carbohydrate digestion and glucose transport both from juice and in the context of a model meal suggest that the benefits of 100% GJ may be extendable beyond the glucose derived from the juice and to the response of a full meal and, thus, have positive impacts on health.

 ,
, 



{kind=link}
{kind=link}
{kind=link}