1. Introduction
Diabetes mellitus (DM) is a severe metabolic disease, and numerous complications are associated with the characteristic hypergly-cemia of this disease [
1]. Of these, diabetic retinopathy (DR) is one of the major microvascular complications amongst diabetic patients, and is the primary cause of visual loss [
1]. Increasing evidence indicates that the chronic uncontrolled hyperglycemic state leads to generation of reactive oxygen species (ROS), which triggers a severe inflammatory state characterized by an elevation of proinflammatory cytokines [
2,
3,
4]. The proinflammatory cytokines tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β) are positively correlated with blood retinal barrier (BRB) breakdown and vascular cell death [
5,
6,
7]. Apart from this, chronic hyperglycemia stimulates synthesis and secretion of vascular endothelial growth factor (VEGF), which was transcriptional regulated by hypoxia-inducible factor-1α (HIF-1α), is the major growth factor mediating retinal vascular leakage and neovascularization [
8,
9].
There is growing evidence that the number of patients with DR keeps growing due to difficulty in achieving tight glycemic control, unresponsive to the current therapeutic approaches and significant side-effects from therapies [
10]. Laser photocoagulation is currently the primary method of treatment for patients with diabetic retinopathy who are at a high risk of vision loss, but unfortunately this is not always effective for improving vision [
11]. There is a great need to develop new therapeutic approaches for this devastating disease. In recent years, clinical and epidemiological evidence suggests that a diet rich in polyphenol may decrease the risk of chronic diseases associated with oxidative stress including diabetes [
12,
13].
Plantaginis semen (Cheqianzi), the dried, ripe seed of
Plantago species (Plantaginaceae), has been traditionally used as medicines and supplements for improving blurred vision or internal oculopathy due to yin deficiency of the liver and kidneys in Asia [
14].
Plantago asiatica L. and
Plantago depressa Willd are the official sources of plantaginis semen in Chinese Pharmacopoeia [
15]. These medicinal materials are produced in all parts of China, collected in summer and autumn when the seed is ripe [
15]. Plantaginis semen has other uses in folk medicine including treating edema, dysuria, stranguria with burning pain and blood, and removing dampness to arrest diarrhea as well as clearing heat from the lungs and resolve phlegm [
16,
17]. One of the major constituents in plantaginis semen is mucilage, which has activity to lower glycemic index of food in human [
18]. In addition, flavonoids are abundant in plantaginis semen [
19]. It is apparent that the polyphenols could improve glucose homeostasis through potential multiple mechanisms of action [
13]. The evidence also suggested that polyphenolic flavonoids are capable of acting on various mechanisms or etiological factors responsible for the development of different sight threatening ocular diseases [
20]. Thus, plantaginis semen seems be valued for adjuvant therapy in the control of free radical-mediated diseases and/or diabetes-related microvascular complications. However, there are no scientifically proven data to show plantaginis semen is responsible for the protective effect on diabetic retinal tissues.
STZ-induced type 1 diabetes in rodents are commonly used as DR animal model. Many researches used STZ-diabetic rats to study DR and other diabetic complications [
21]. The goal of this study was to determine whether plantaginis semen can be used for treatment of STZ-induced DR in rats and its underlying mechanism.
2. Materials and Methods
2.1. Preparation of Plant Extracts
Plantaginis semen were purchased from Jinbaoan Trade Co., Ltd. (Zhunan Township, Miaoli County, Taiwan) in September 2015, and identified by Hong T.Y. (Department of Biotechnology, Collage of Pharmacy and Health Care, Tajen University, Taiwan). The voucher specimen (Lot No. PS 20150926) was deposited in our laboratory. Plantaginis semen (10 kg) were grounded into a 40-mesh powder and extracted with 95% ethanol (5 volumes of ethanol) in a stainless steel extraction tank for 24 h at room temperature. This ethanol mixture was filtered through funnels and centrifuged (4 °C, 1350× g, 20 min), and this was repeated three times. All of the precipitate was eliminated, and the ethanol extract portion was collected and concentrated using a rotary evaporator. Plantaginis semen ethanol extract (PSEE) was then evaporated under reduced pressure conditions, which completely eliminated the alcohol, followed by lyophilization, yielding approximately 1328 g of dry residue (w/w yield: 13.2%). PSEE was kept at −20 °C until use and suspended in distilled water.
2.2. Total Phenolic Content
Polyphenol content of PSEE was determined according to the Folin-Ciocalteu colorimetric method [
22]. Standard gallic acid (Sigma-Aldrich, Inc., Saint Louis, MO, USA) and an aliquot of PSEE were diluted with an ethanol/water (60:40,
v/
v) solution containing 0.3% HCl. Two mL of 2% Na
2CO
3 was mixed into each sample of 100 μL and allowed to equilibrate for 2 min before adding 50% Folin-Ciocalteu’s phenol reagent (Sigma-Aldrich, Inc.). Absorbance at 750 nm was measured at room temperature. The polyphenol content of PSEE was expressed as mg of gallic acid equivalent per gram (g) of PSEE in dry weight (DW), i.e., mg gallic acid/g DW.
2.3. Total Flavonoid Content
The total flavonoid content of PSEE was determined by the aluminium chloride colorimetric method [
23]. Briefly, 0.25 mL of PSEE (100 μg/mL) was added to a tube containing 1 mL of double-distilled water. Next, 0.075 mL of 5% NaNO
2, 0.075 mL of 10% AlCl
3, and 0.5 mL of 1 mmol/L NaOH were added sequentially at 0, 5, and 6 min. Finally, the volume of the reacting solution was adjusted to 2.5 mL with double-distilled water. The solution had an absorbance of 510 nm. The total flavonoid content was calculated from a calibration curve, and the result was expressed as mg rutin (Sigma-Aldrich, Inc.) equivalent per g dry weight, i.e., mg rutin/g DW.
2.4. Total Antioxidant Capacity
The total antioxidant capacity of PSEE was determined using the horseradish peroxidase catalyzed oxidation of 2,2-azino-bis-(3-ethyl benzothiazoline-6-sulfonicacid) (ABTS) [
24]. The reaction mixture contained 0.5 mL of 1000 μmol/L ABTS (in ddH
2O) and 3.5 mL of 100 μmol/L H
2O
2. The reaction was started by adding 0.5 mL of 44 U/mL peroxidase (in 0.1 mol/L phosphate-buffered saline (PBS)). After 1 h, 0.05 mL of PSEE were added to the mixture. After 5 min, absorbance was measured at 730 nm. Trolox (Sigma-Aldrich, Inc.) standard solution were prepared and assayed under the same conditions. Results were expressed in terms of trolox equivalent antioxidant capacity (TEAC), i.e., mmol/L Trolox/100 g DW.
2.5. Experimental Animals
All experimental methods and animal care procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of Tajen University (approval number, IACUC 104-28; approval date: 12 November, 2015), in accordance with the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health, as well as the guidelines of the Animal Welfare Act. Male Wistar rats (8 weeks aged) weighting 200–250 g, were purchased from National Laboratory Animal Center (Taipei, Taiwan) and housed two per cage in a room under controlled temperature (20–25 °C), humidity (50% ± 5%) and lighting (12 h light/dark cycle) with food and water provided ad libitum. Rats were rendered diabetic by a single intravenous injection of 60 mg/kg streptozotocin (STZ; Sigma-Aldrich, Inc.). Eight-week age-matched control rats were injected with vehicle (sterile saline 0.9%, pH 7.4). After 1 week, rat with non-fasting blood glucose levels >350 mg/dL, polyuria, and glucosuria were defined as diabetic and used for the experiments.
2.6. Treatment Protocols
In the treatment group (𝑛 = 10 per group), STZ-diabetic rats were dosed by oral gavage once per day for 8 weeks with PSEE at dosages of 100, 200, or 300 mg/kg in a volume of 1.5 mL/kg distilled water. The dosages of PSEE were selected based on consideration of tests of Plantago extracts relating to producing preventive effects on oxidative damage in rats [
25]. A vehicle-treated group (𝑛 = 10 per group). Of normal rats and STZ-diabetic rats were treated with 1.5 mL/kg distilled water only over the same treatment period. Animals had free access to standard rat diet (Harlan Teklad, Madison, WI, USA; catalogue number (Cat. No.) 2018) and water throughout the entire treatment period.
At the end of the 8-week treatment, the rats were weighed, fasted overnight and anesthetized using an intraperitoneal injection of sodium pentobarbital (60 mg/kg). While under anesthesia, they were painlessly sacrificed and blood was collected from the abdominal aorta of each animal into heparin sample bottles. Rat eyes from each group were removed and the retinae were isolated. The diagnostic kit for determination for plasma levels of glucose (Cat. No. COD12503) was purchased from BioSystem (Barcelona, Spain). Commercial enzyme-linked immunosorbent assay (ELISA) kits were used to quantify HbA1c levels (Integrated Bio Ltd., Taipei, Taiwan; Cat. No. CSB-E08140r). All analyses were performed in accordance with the instructions provided by the manufacturers.
2.7. Fundus Photography and Vessel Diameter
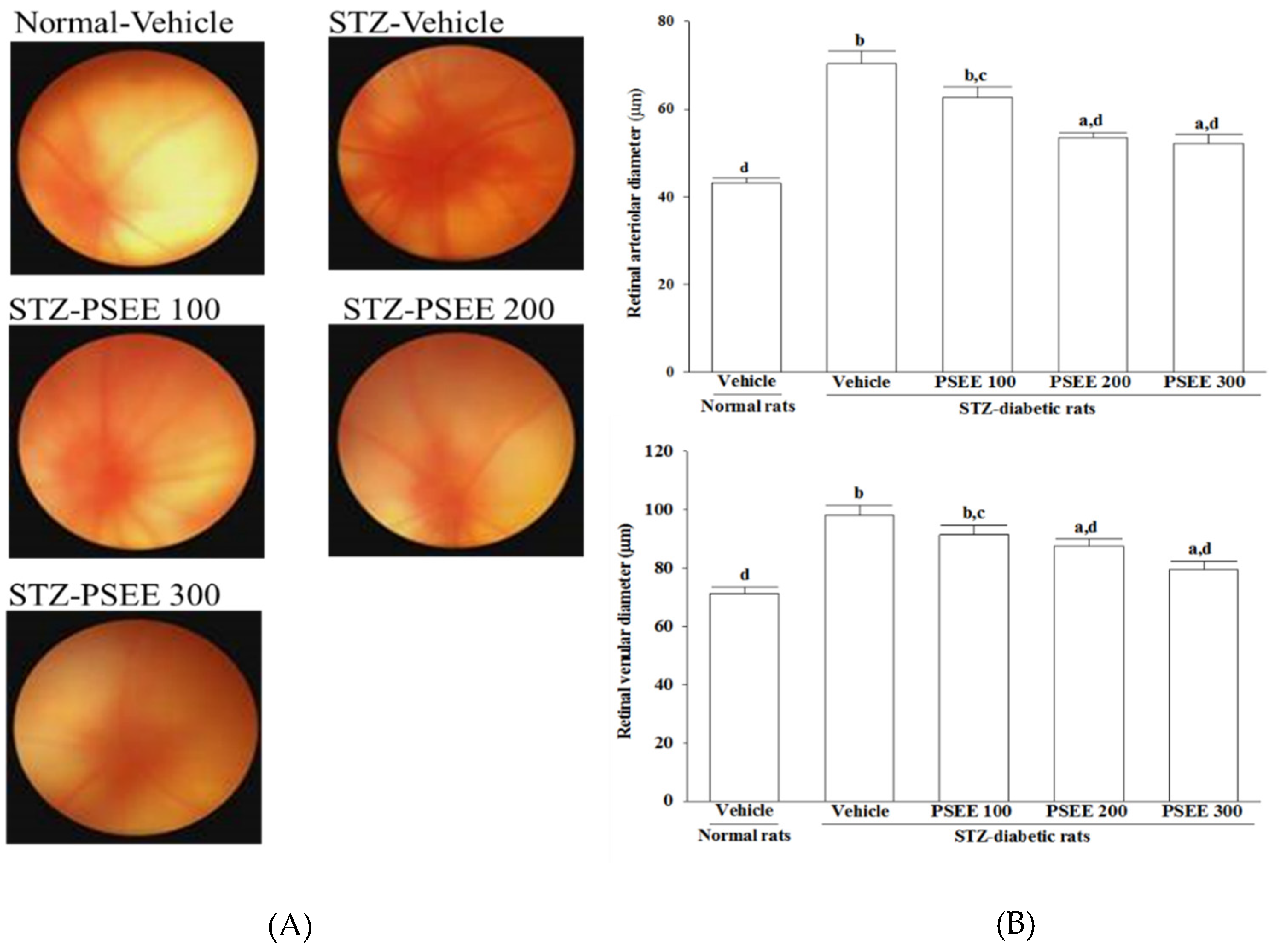
Fundus photography is performed with a retina camera (Kowa Company Ltd., Tokyo, Japan). In order to accustom to the fundus photography procedure, rats were trained before start of the study. Eyes were dilated with a drop of 1% tropicamide (Synpac-Kingdom Pharmaceutical Co., Ltd., Taipei, Taiwan). Moisol eye drops were administered periodically to prevent the cornea from drying out. Fundus photography was done regularly till 8 weeks to monitor the fundus changes.
The diameter of retinal vessels was estimated by previously described method [
26]. Before diameter estimation, the retinal photographs from all groups were randomized. The vessel diameter of 3 most prominent vessels was estimated at 3 sites in its widest portion at equal distance from the center. An average of 3 estimations was taken as the final retinal vessel diameter.
2.8. Quantification of Retinal Leukostasis
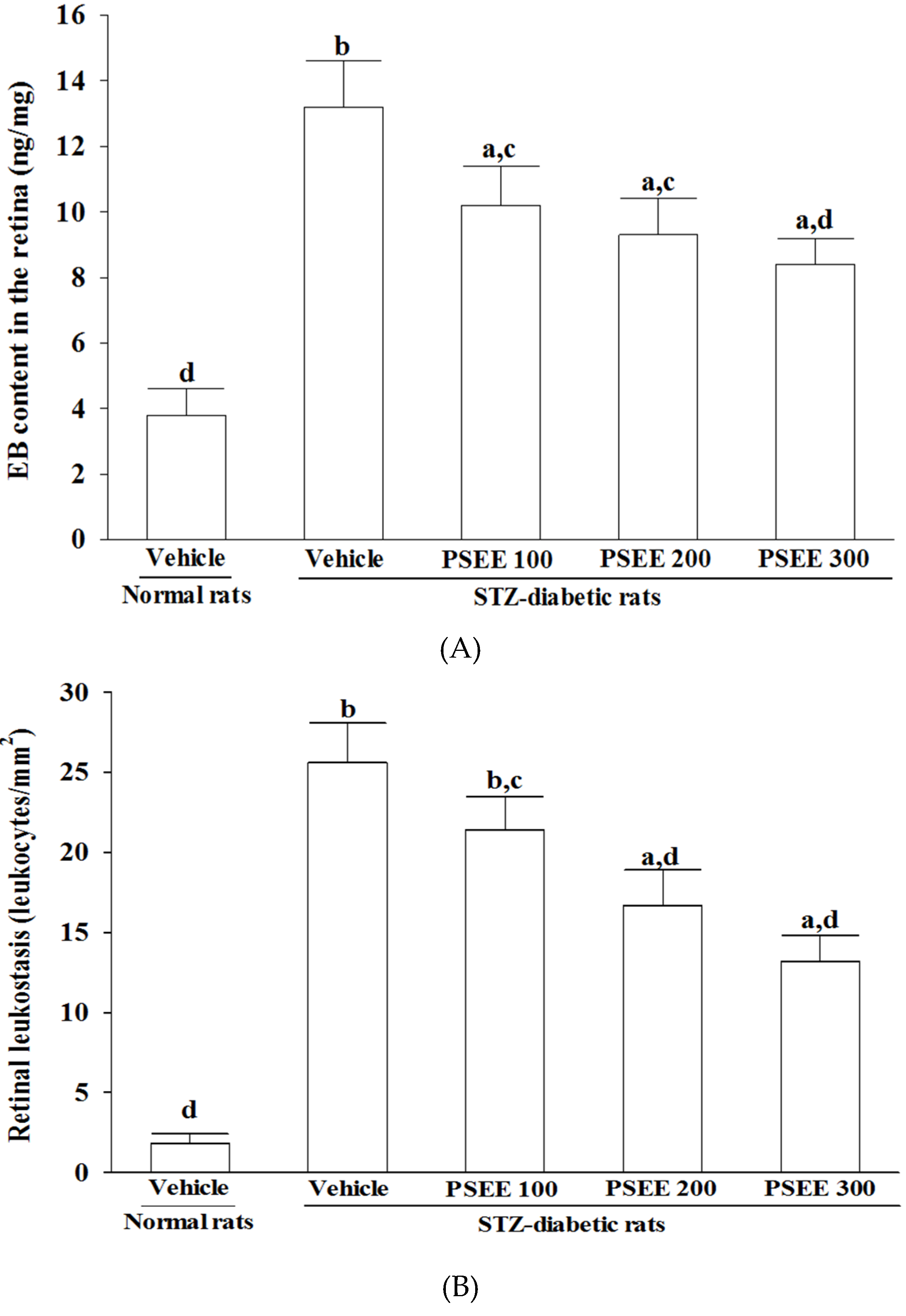
Quantification of leukostasis was performed at the end of the 8-week treatment by previously described method [
27]. The chest cavity of each deeply anesthetized rat was carefully opened and a perfusion needle was inserted into the left ventricle. After cutting the right atrium, the animals were immediately perfused with 500 mL of PBS per kg body weight and heparin (0.1 mg/mL) to wash out nonadherent blood cells. Fluorescein isothiocyanate-coupled Concanavalin A lectin (ConA) (20 μg/mL in PBS; pH 7.4; 5 mg/kg; Vector Laboratories, Burlingame, CA, USA) was then perfused to label adherent leukocytes and vascular endothelial cells. Residual unbound ConA was flushed by PBS perfusion. Eyes were removed and fixed in 4% paraformaldehyde for 1 h. Retinas were dissected and flat mounted on a microscope slide, covered with anti-fading medium and a coverslip, and imaged via fluorescence microscopy. Only whole retinae in which the entire vascular network was stained were used for analysis. The total number of adherent leukocytes within the vessels of each retina was counted.
2.9. Retinal Permeability Assessment
Retinal vascular permeability was measured using Evans blue (EB) dye extravasation technique at the end of the 8-week treatment [
28]. EB dye (Sigma-Aldrich, Inc.) was dissolved in normal saline at 45 mg/mL and was injected through the tail vein of anesthetized rats over 10 s at a dosage of 45 mg/kg. After the dye had circulated for 2 h, the rats were anesthetized with sodium pentobarbital (40 mg/kg), the chest cavity was opened, and cardiac perfusion was performed via the left ventricle with 1% paraformaldehyde in citrate buffer (0.05 mol/L, pH 3.5) under a constant pressure of 120 mmHg. Immediately after perfusion, the retinas were carefully dissected under an operating microscope. After retinas were fully dried at 4 °C, then the weights of them were measured, EB dye was extracted by incubating each sample in 150 µL formamide for 18 h at 70 °C. The extract was ultracentrifuged at a speed of 14,000 rpm for 60 min. Absorbance was measured using 100 µL of the supernatant at 620 nm and 740 nm. The concentration of EB in the extracts was calculated from a standard curve and normalized by total protein concentration in the tissue.
2.10. Assay of Retinal Antioxidant Enzymes
Retinas from right and left eyes from one rat were pooled as one sample, and then were homogenized in 10 volume of ice cold 0.1 M Tris-HCl, pH 7.4 containing 0.5% Triton X-100, 5 mmol/L β-mercaptoethanol, 0.1 mg/mL phenylmethylsulfonyl fluoride and centrifuged at 14,000× g for 5 min at 4 °C. The supernatant was collected and used for following experiments as described below. Protein concentration of the supernatant was assayed by Bio-Rad protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA). The intracellular activity of superoxide dismutase (SOD; Cat. No. ab65354), catalase (CAT; Cat. No. ab118184) and glutathione (GSH; Cat. No. ab65322) were estimated using commercially available assay kits from Abcam plc. (Cambridge, MA, USA). All assays were carried out in triplicates.
2.11. Protein Extraction and Western Blot Analyses
Retinas from right and left eyes from one rat were pooled as one sample, which were then homogenized in 1 mL of ice-cold hypotonic buffer A (10 mmol/L of HEPES, 10 mmol/L of KCl, 2 mmol/L of MgCl2, 1 mmol/L of dithiothreitol, 0.1 mmol/L of EDTA, and 0.1 mmol/L of phenylmethylsulfonylfluoride; pH 7.8). A solution of 80 μL of 10% Nonidet P-40 was added to the homogenates, and the mixture was centrifuged for 2 min at 14,000× g at 4 °C. Before immunoblotting, and the protein concentration of each sample was determined using a Bio-Rad protein assay kit and bovine serum albumin as a standard, to ensure equal loading among lanes.
The tissue lysates containing 40–50 mg protein were electrophoresed through 8%, 12%, and 15% sodium dodecyl sulfate-polyacrylamide gels. According to the manufacturer’s instructions, separated proteins were electrophoretically transferred to a nitrocellulose membrane, blocked with 5% skim milk solution for 1 h, and incubated with primary antibodies to TNF-α (Cat. No. sc-1348), IL-1β (Cat. No. sc-7884), intercellular adhesion molecule 1 (ICAM-1; Cat. No. sc-8439), vascular cell adhesion molecule 1 (VCAM-1; Cat. No. sc-8304), HIF-1α (Cat. No. sc-1836), VEGF (Cat. No. sc-1836), Akt (Cat. No.sc-5298), pAkt (Ser 473) (Cat. No. sc-135651), pAkt (Thr 308) (Cat. No. sc- sc-16646-R) and β-actin (Cat. No. sc-20357) at 4 °C overnight, respectively. All antibodies were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA) and used at a dilution of 1:1000. After three 5 min washes in Tris-buffered saline with Tween(TBST; 20 mmol/L Tris-HCl, pH 7.5, 150 mmol/L NaCl, and 0.05% Tween 20), membranes were incubated with the appropriate peroxidase-conjugated secondary antibodies. With the ECL advance Western Blotting detection kit (Cat. No. RPN2135; GE Healthcare Life Sciences, Piscataway, NJ, USA), the membranes were washed three times in TBST and visualized on X-ray film. Band densities were determined using ATTO Densitograph Software (ATTO Corporation, Tokyo, Japan) and quantified as the ratio to β-actin. The mean value for samples was adjusted to a value of 1.0 from the vehicle-treated normal rats on each immunoblot, expressed in densitometry units. Then, all experimental sample values were expressed to this adjusted mean value.
2.12. Real-Time Polymerase Chain Reaction (PCR)
Total RNA was extracted from rat retinas using Trizol reagent (Invitrogen; Boston, MA, USA) according to the manufacturer’s protocol. Two retinas from right and left eyes from one rat were pooled as one sample. RNA was quantified by measuring absorbance at 260 nm and its integrity verified by agarose gel electrophoresis using ethidium bromide for visualization. For the reverse transcriptase reaction, 1 μg of total RNA per sample and 8.5 μg/μL random hexamer primers were heated at 65 °C for 5 min and then quenched on ice. This mixture was combined with 500 μmol/L each of dATP, dTTP, dCTP, and dGTP, 10 mmol/L dithiothreitol, 20 mmol/L Tris-HCl (pH 8.4), 50 mmol/L KCl, 5 mmol/L MgCl
2, 40 units of RNaseOUTTM recombinant ribonuclease inhibitor (Invitrogen) and 100 units SuperScript III reverse transcriptase (Invitrogen). Samples were subjected to DNase (Promega; Madison, WI, USA) treatment at 37 °C for 20 min in a GeneAmp 9700 Thermal Cycler (Applied Biosystems; Foster City, CA, USA) and then held at 4 °C. After aliquots were taken for immediate use in PCR, the remainder of the cDNA was stored at −20 °C. mRNA expression was measured by quantitative real-time PCR in a fluorescent temperature Lightcycler 480 (Roche Diagnostics; Mannheim, Germany). The sequences of the primers were as follows: for TNF-α, 5′-ACACCATGAGCACGGAAAGC-3′ (forward) and 5′-CCGCCACGAGCAGGAA-3′ (reverse); for IL-1β, 5′-AATGGACAGAACATAAGCCAACA-3′ (forward) and 5′-CCCAAGGCCACAGGGAT-3′ (reverse); for ICAM-1, 5′-CGGGTTTGGGCTTCTCC-3′ (forward) and 5′-GCCACTGCTCGTCCACATAG-3′ (reverse); for VCAM-1, 5′-ATCTTCGGAGCCTCAACGG-3′ (forward) and 5′-CCAATCTGAGCGAGCGTTT-3′ (reverse); for HIF-1α, 5′-GTCGGACAGCCTCACCAAACAG-3′ (forward) and 5′-TAGGTAGTGAGCCACCAGTCATCCAAGGAA-3′ (reverse); for VEGF, 5′-ACAGGGAAGACAATGGGATGA-3′ (forward) and 5′-GGGCCAGGGATGGGTTT-3′ (reverse); for β-actin, 5′-TGTGATGGTGGGAATGGGTCAG-3′ (forward) and 5′-TTTGATGTCACGCACGATTTCC-3′ (reverse). Primers were designed using Primer Express Software version 2.0 System (Applied Biosystems; Foster City, CA, USA). The PCR reaction was performed using the following cycling protocol: 95 °C for 5 min, followed by 45 cycles of 95 °C for 5 s, 58 °C for 15 s, and 72 °C for 20 s. Dissociation curves were run after amplification to identify the specific PCR products. The mRNA expression levels were normalized to β-actin mRNA levels and calculated according to the delta-delta Ct method [
29].
2.13. Activity of NF-κB, Histone Deacetylases (HDAC) and Histone Acetyltransferases (HAT)
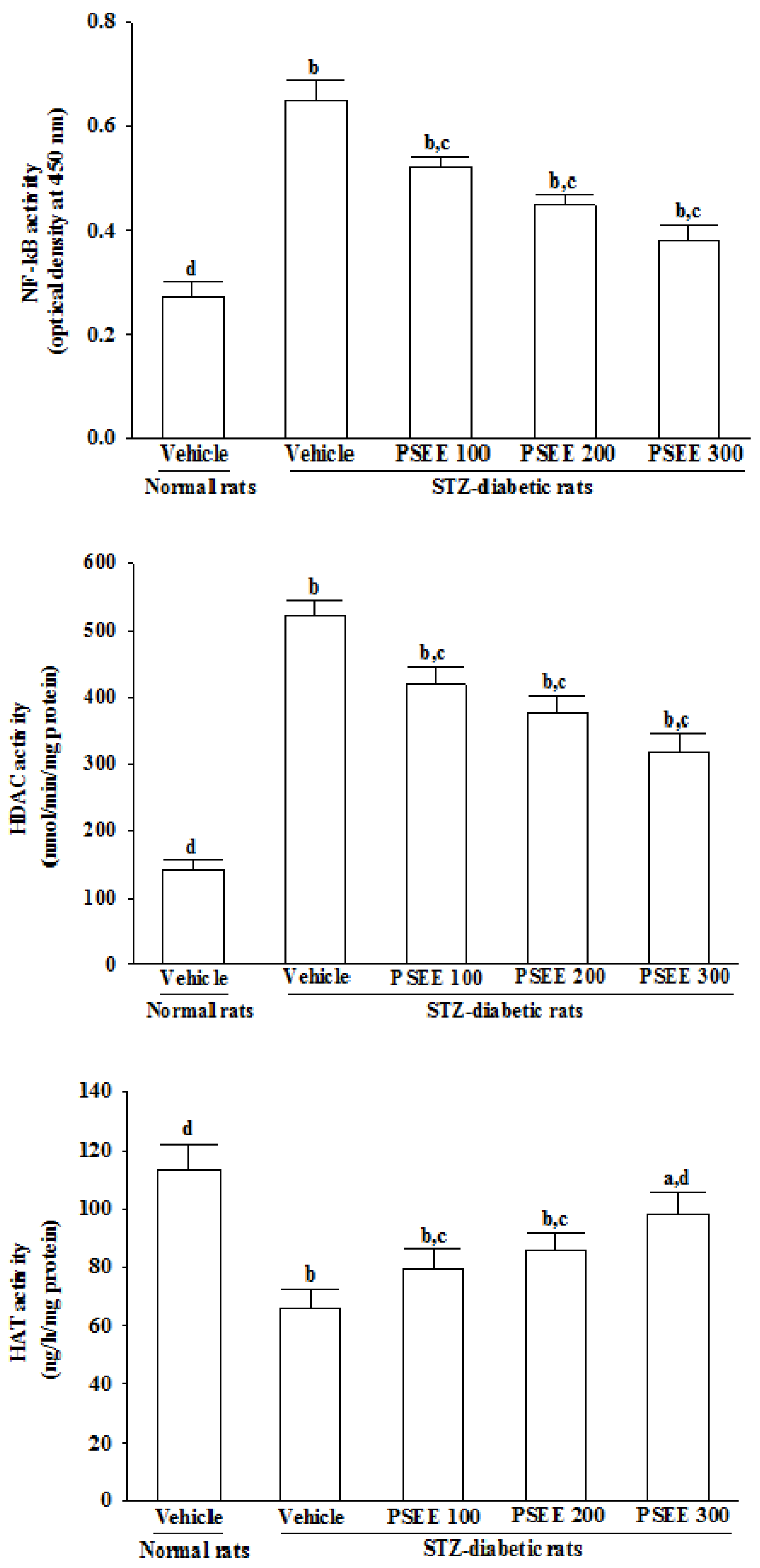
Nuclear extract of retina was prepared using the nuclear extract kit (Cat. No. 40410; Active Motif, Carlsbad, CA, USA) following manufacturer's protocol. Two retinas from right and left eyes from one rat were pooled as one sample. Nuclear factor-κB (NF-κB) activation was determined, TransAM® NF-κB p65 transcription factor assay kit (Cat. No. 40596) implemented under the procedures provided by the manufacturer (Active Motif Inc., Carlsbad, CA, USA). Reaction was quantified at 450 nm. Histone deacetylases (HDAC) activity in retinal nuclear extracts was measured by HDAC activity colorimetric assay kit (Cat. No. K331-100) from BioVision Inc. (Milpitas, CA, USA). The colorimetric readings were measured at 400 nm in a spectrophotometer. Activity of histone acetyltransferases (HAT) was quantified in retinal nuclear fraction by non-radioactive indirect ELISA kit (Cat. No. K332-100) from BioVision Inc. The readings were monitored at 440 nm in a spectrophotometer.
2.14. Statistical Analysis
All statistical analyses were performed using SPSS for Windows (version 21.0; IBM Corporation, Armonk, NY, USA). The results are presented as the mean ± standard deviation (SD) for each group of animals at the number (n) indicated. The significance of differences between groups was evaluated by oneway ANOVA with Fisher’s Least Significant Difference post hoc test; and p < 0.05 was considered as indicating statistically significant differences. Relationships between variables were examined using Pearson correlations.
4. Discussion
Data from multicenter prospective studies have shown that an abnormally high glucose concentration in blood is the principal cause of microvascular and macrovascular complications [
1]. Therefore, tight control of blood glucose is the key to preventing or reversing diabetic complications in diabetic patients [
11]. In vitro and in vivo studies have shown that dietary polyphenols may inhibit α-amylase and α-glucosidase, inhibit glucose absorption in the intestine by sodium-dependent glucose transporter 1, stimulate insulin secretion and reduce hepatic glucose output, suggesting that polyphenols could improve glucose homeostasis through potential multiple mechanisms of action and might be one dietary therapy for the prevention and management of diabetes [
13]. In the present study, PSEE treatment showed significant and consistent reduction in fasting blood glucose levels and also improved the body weight loss in STZ-diabetic rats as compared to the vehicle treated diabetic controls, indicating its potent antidiabetic activity on an insulin deficient animal model. PSEE was rich in polyphenolic flavonoids, and its antidiabetic activity may be attributed to the presence of these.
Antioxidant nutrients and phytonutrients have been reported to inhibit the oxidation of living cells by free radicals and result in a decrease in oxidative stress [
30]. In the present study, the antioxidant capacity of PSEE was measured by ABTS radical cation decolorization assay and showing promising results. It is clear that PSEE was rich in polyphenols and shown with antioxidant potential. Actually, long-term hyperglycemia could lead to an increase in ROS generation and decreased antioxidant capacity in diabetes [
3]. The retina is particularly susceptible to oxidative stress because of high energy demands and exposure to light [
31]. Regarding the oxidative stress affects the pathogenesis of DR, correction of oxidant-antioxidant balance may be a powerful approach for preventing vision loss associated with DR [
3]. It is well known that SOD, CAT, and GSH constitute a mutually supportive team of defense against ROS [
4]. In our study, decline in the activities of these enzymes in the retinal tissue of STZ-diabetic rats and attainment of near normalcy in PSEE-treated rats indicate that oxidative stress elicited in the retina of diabetic rats had been nullified due to the effect of PSEE. Thus, our results suggest that PSEE has potential to overcome the hyperglycemia-specific microvascular complications.
The growing evidence has suggested that BRB breakdown, leakage capillaries and vascular structural and functional changes are characteristic for the diabetic retina [
6]. Similar with the previous study [
32], we observed that the number of leukocytes adhered to the retinal vascular endothelium was increased in STZ-diabetic rats; accordingly, vascular permeability and the retinal vessels diameter were increased as well. In consistent with the attenuated leukostasis, PSEE reduced diabetic retinal vascular leakage accompanied by restrained the retinal vascular dilation in STZ-diabetic rats. Prevention of diabetes-related structural disorganization of the retina might be an important contributor of PSEE to the preventing the progression of DR.
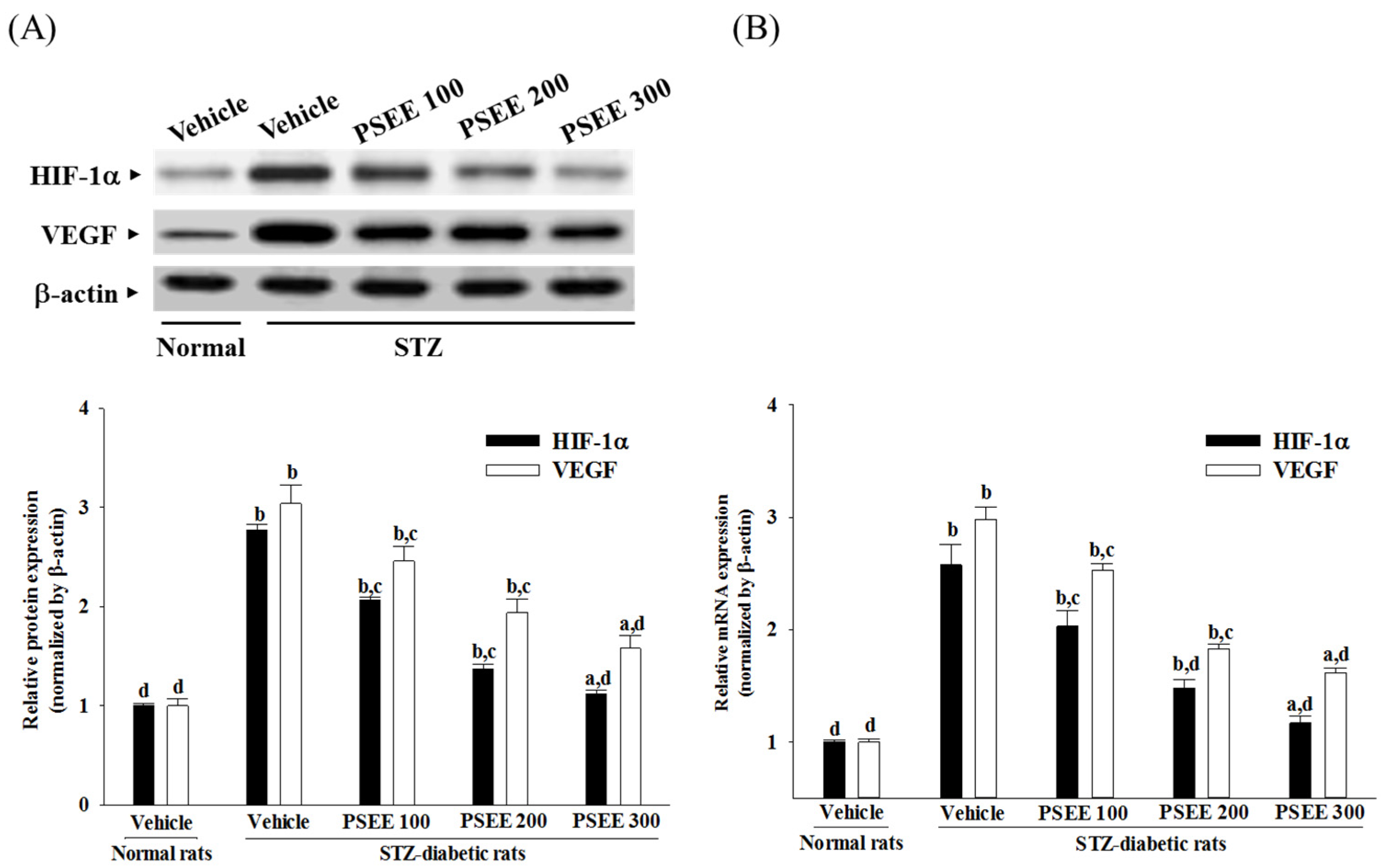
VEGF, an endothelial angiogenic and vasopermeability factor, is known to be a key molecule leading to retinal permeability and breakdown of BRB in diabetes and other retinal diseases [
33]. Regulation of VEGF expression is complex, and HIF-1α is one of the transcription factors that regulate VEGF expression under hyperglycemia [
8,
9]. Furthermore, Akt activation has been recognized as an upstream regulator of HIF-1α expression [
34]. Therefore, promotion of the Akt-HIF-1α-VEGF signaling pathway contributes to the induction of retinal vascularization [
35]. We found that the elevated contents of HIF-1α and VEGF in retinae of STZ-diabetic rats were both reduced in rats receiving PSEE treatment. In addition, the results of the present study revealed an increase in phosphorylation of Thr 308 and Ser 473 of Akt in the retinas of STZ-diabetic rats; the deficit was ameliorated by PSEE treatment. Therefore, it can be considered that PSEE rescued diabetic retinal vasculopathy by downregulation of HIF-1-mediated induction of VEGF expression via suppressing Akt activation in the retina of STZ-diabetic rats. Full activation of Akt requires phosphorylation on Thr 308 and Ser 473 by 3-phosphoinositide-dependent kinase-1 and Ser-473 kinase, respectively [
36]. Thus, the role of PSEE on the alterations in Akt signaling in the development of DR will be identified in future research work.
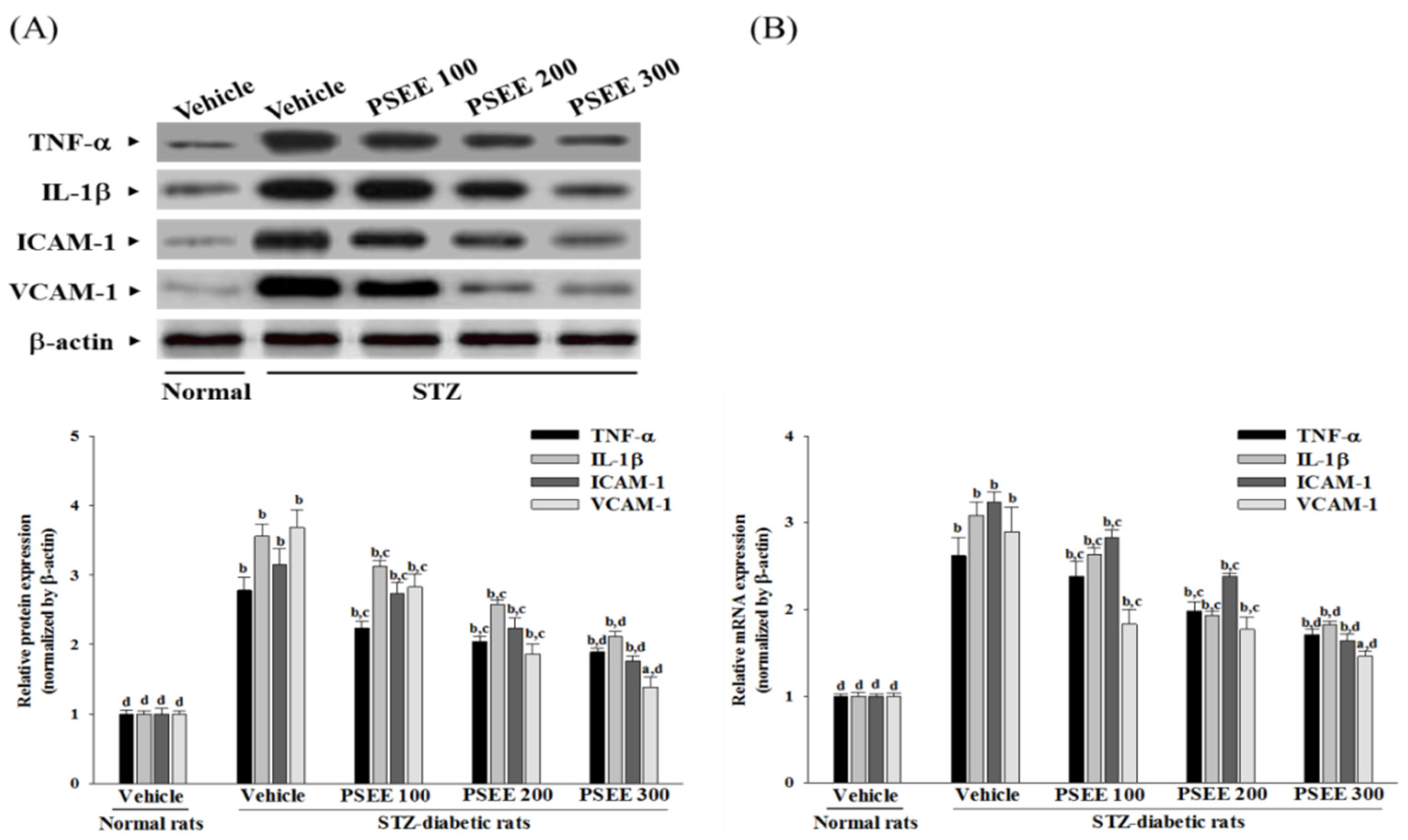
Numerous studies show that hyperglycemia leads to oxidative stress in the diabetic retinas, which has been associated with cellular inflammation and release of inflammatory cytokines [
2]. One of these mediators is TNF-α, a proinflammatory cytokine which is known as an initiator of inflammatory reactions [
7]. Similarly, IL-1β can be up-regulated in the retina in diabetes [
5]. In addition to increases in the above-mentioned inflammatory mediators, both molecules ICAM-1 and VCAM-1 promote chemoattraction of leukocytes into the vascular walls and their migration into retinal tissues, which accounts for the majority of diabetes-associated retinal vascular leakage [
37]. Actually, plantaginis semen significantly inhibited lipopolysaccharide-induced cyclooxygenase-2 (concentration required for 50% inhibition [IC(50)] = 8.61 μg/mL, TNF-α [IC(50)] = 9.63 μg/mL, and nitric oxide [IC(50)] = 8.65 μg/mL) production in RAW 264.7 cells; anti-inflammatory activity of plantaginis semen has been reported [
38]. In the present study, retinae from PSEE-treated STZ-diabetic rats showed lower levels of inflammatory cytokines and chemokines, suggesting that the extract acted against inflammatory response triggered by hyperglycemia. These results support the proposition that protection of PSEE from retinal damage in STZ-diabetic rats was mediated by blockade of diabetes-induced production of inflammatory molecules in retinal tissue, and attenuates retinal vascular leakage.
NF-κB plays a critical role in diabetes complications as it regulates transcription of a number of genes involved in inflammatory response [
39]. Actually, it has been demonstrated that the subunits of the NF-κB signaling pathway, including the inhibitory transcription factor IκB, are acetylated/deacetylated by the HAT and HDAC, respectively [
40]. High HDAC activity may therefore maintain deacetylated such inhibitory factor allowing for NF-κB activation [
41]. Recent studies have shown that diabetes induced increase HDAC activity in the retina and kidney, that are the tissues associated with microvascular complications [
42]. It has also been reported that hyperglycemia-induced superoxide overproduction activates HDAC activity and decreases HAT activity [
43]. PSEE treatment results in a significant increase in HAT activity and a parallel decrease in activies of HDAC and NF-κB in retinae of STZ-diabetic rats. The effects of PSEE seems to play a role in controlling NF-κB activation and modulation of HDAC and HAT activity, consequently affecting the expression of inflammatory response genes.
Medicinal plants produced several useful biological activities; however, the inclusion of toxicological evaluation at preclinical stage will assure its safe usage in humans as a medicine [
44]. Further studies are needed to clarify toxicity of PSEE to rat at the effective dosage used for treating DR. Whether PSEE is effective in human for DR improvement also need further evaluate in clinical studies.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}