Impact of Glycosidic Bond Configuration on Short Chain Fatty Acid Production from Model Fermentable Carbohydrates by the Human Gut Microbiota


Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrates
2.2. Batch Fermentations
2.3. Participants
2.4. SCFA Extraction and Analysis
3. Results
3.1. Effects of Glycosidic Bond Position on Fermenter pH
3.2. Effects of Glycosidic Bond Position on SCFA Production
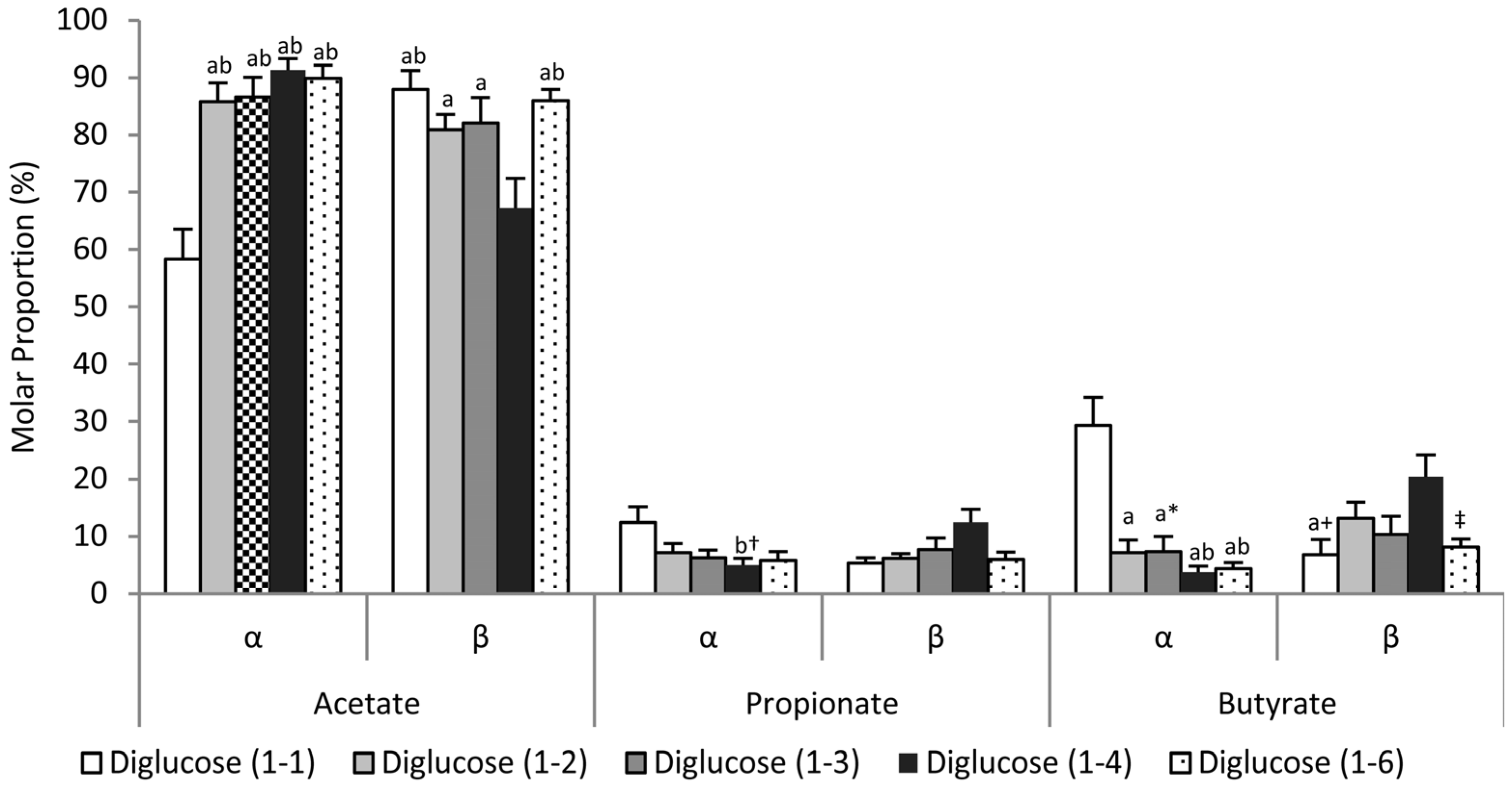
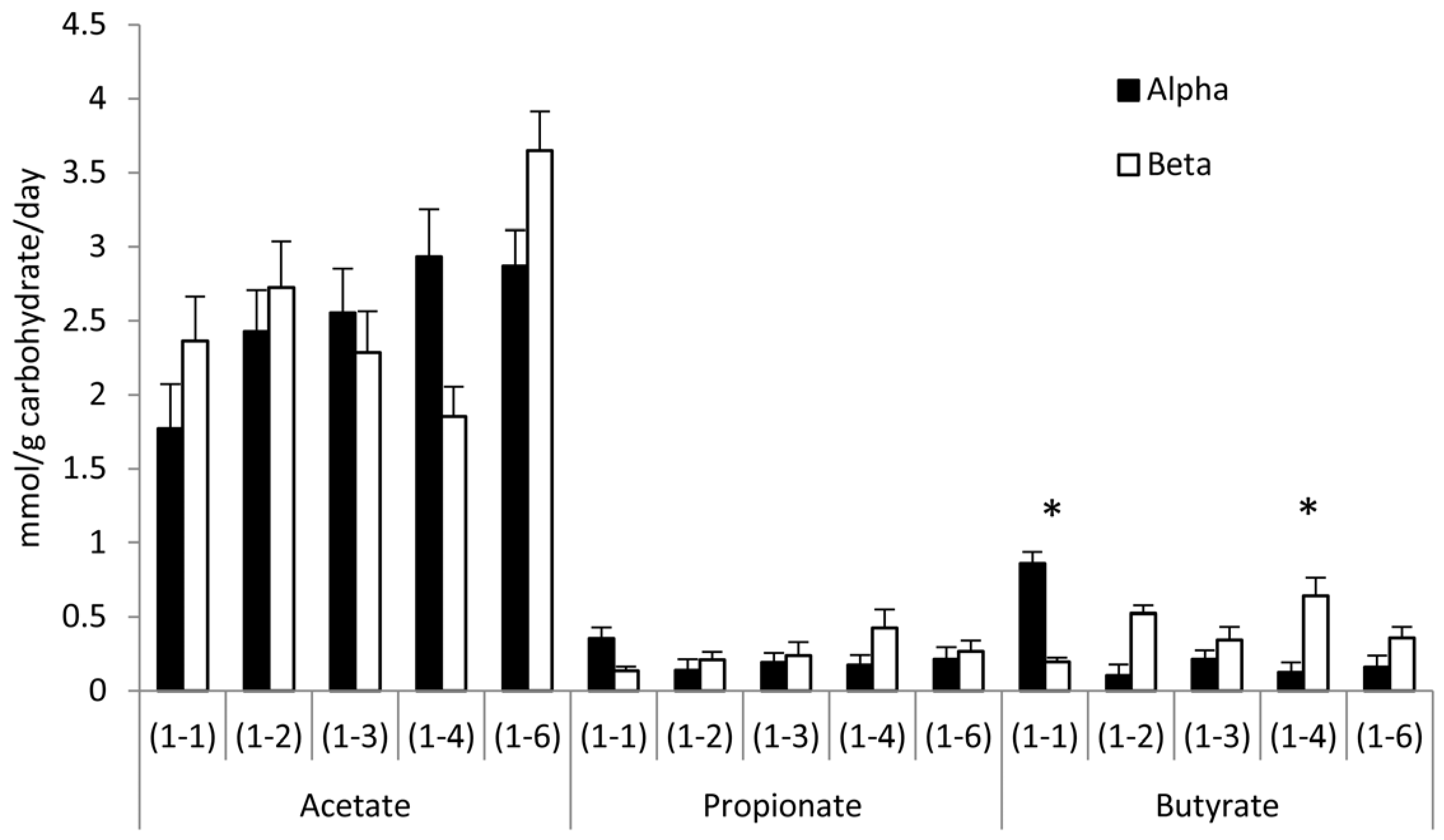
3.3. Effects of Glycosidic Bond Orientation (α or β) on SCFA Production
3.4. Subject SCFA Variability
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| FFAR2/3 | Free fatty acid receptor 2/3 |
| GC-FID | Gas chromatography flame ionization detector |
| GLP-1 | glucagon-like peptide-1 |
| NDC | non-digestible carbohydrate |
| OFN | oxygen free nitrogen |
| PYY | peptide YY |
References
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.K.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Plessier, F.; Gautier-Stein, A.; Mithieux, G. Vasoactive intestinal peptide is a local mediator in a gut-brain neural axis activating intestinal gluconeogenesis. Neurogastroenterol. Motil. 2015, 27, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.W.; de Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, K.A.; Boobis, A.R.; Chiodini, A.; Edwards, C.A.; Franck, A.; Kleerebezem, M.; Nauta, A.; Raes, J.; van Tol, E.A.; Tuohy, K.M. Towards microbial fermentation metabolites as markers for health benefits of prebiotics. Nutr. Res. Rev. 2015, 28, 42–66. [Google Scholar] [CrossRef] [PubMed]
- Wolever, T.M.S.; Spadafora, P.; Eshuis, H. Interaction between colonic acetate and propionate in humans. Am. J. Clin. Nutr. 1991, 53, 681–687. [Google Scholar] [PubMed]
- Den Besten, G.; Lange, K.; Havinga, R.; van Dijk, T.H.; Gerding, A.; van Eunen, K.; Müller, M.; Groen, A.K.; Hooiveld, G.J.; Bakker, B.M.; et al. Gut-derived short-chain fatty acids are vividly assimilated into host carbohydrates and lipids. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G900–G910. [Google Scholar] [CrossRef] [PubMed]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [PubMed]
- Wright, R.S.; Anderson, J.W.; Bridges, S.R. Propionate inhibits hepatocyte lipid synthesis. Exp. Biol. Med. 1990, 195, 26–29. [Google Scholar] [CrossRef]
- Nishina, P.M.; Freedland, R.A. Effects of propionate on lipid biosynthesis in isolated rat hepatocytes. J. Nutr. 1990, 120, 668–673. [Google Scholar] [PubMed]
- Roediger, W.E. Role of anaerobic bacteria in the metabolic welfare of the colonic mucosa in man. Gut 1980, 21, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Belcheva, A.; Irrazabal, T.; Robertson, S.J.; Streutker, C.; Maughan, H.; Rubino, S.; Moriyama, E.H.; Copeland, J.K.; Kumar, S.; Green, B.; et al. Gut microbial metabolism drives transformation of msh2-deficient colon epithelial cells. Cell 2014, 158, 288–299. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C.; Fan, X.; Dikiy, S.; van der Veeken, J.; deRoos, P.; Liu, H.; Cross, J.R.; Pfeffer, K.; Coffer, P.J.; et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.-Y.; Lannoy, V.; Decobecq, M.-E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Van Damme, J.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [PubMed]
- Stoddart, L.A.; Smith, N.J.; Milligan, G. International Union of Pharmacology. LXXI. Free Fatty Acid Receptors FFA1, -2, and -3: Pharmacology and Pathophysiological Functions. Pharmacology 2008, 60, 405–417. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.V.; Frassetto, A.; Kowalik, E.J.; Nawrocki, A.R.; Lu, M.M.; Kosinski, J.R.; Hubert, J.A.; Szeto, D.; Yao, X.; Forrest, G.; et al. Butyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanisms. PLoS ONE 2012, 7, e35240. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, S.; Macfarlane, G.T. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Gietl, E.; Mengerink, W.; de Slegte, J.; Gibson, G.; Rastall, R.; van den Heuvel, E. Factors involved in the in vitro fermentability of short carbohydrates in static faecal batch cultures. Int. J. Carbohydr. Chem. 2012, 2012, 1–10. [Google Scholar] [CrossRef]
- Salvador, V.; Cherbut, C.; Barry, J.L.; Bertrand, D.; Bonnet, C.; Delort-Laval, J. Sugar composition of dietary fibre and short-chain fatty acid production during in vitro fermentation by human bacteria. Br. J. Nutr. 1993, 70, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, P.B.; Holtug, K.; Rasmussen, H.S. Short-chain fatty acid production from mono- and disaccharides in a fecal incubation system: Implications for colonic fermentation of dietary fiber in humans. J. Nutr. 1988, 118, 321–325. [Google Scholar] [PubMed]
- Laurentin, A.; Edwards, C.A. Differential fermentation of glucose-based carbohydrates in vitro by human faecal bacteria—A study of pyrodextrinised starches from different sources. Eur. J. Nutr. 2004, 43, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Mortensen, P.B.; Nordgaard-Andersen, I. The dependence of the in vitro fermentation of dietary fibre to short-chain fatty acids on the contents of soluble non-starch polysaccharides. Scand. J. Gastroenterol. 1993, 28, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Champ, M.M.J. Physiological aspects of resistant starch and in vivo measurements. J. AOAC Int. 2004, 87, 749–755. [Google Scholar] [PubMed]
- Vogt, J.A.; Pencharz, P.B.; Wolever, T.M.S. l-Rhamnose increases serum propionate in humans. Am. J. Clin. Nutr. 2004, 80, 89–94. [Google Scholar] [PubMed]
- Vogt, J.A.; Ishii-schrade, K.B.; Pencharz, P.B.; Wolever, T.M.S. l-Rhamnose increases serum propionate after long-term supplementation, but lactulose does not raise serum acetate. Am. J. Clin. Nutr. 2004, 80, 1254–1261. [Google Scholar] [PubMed]
- Sanz, M.L.; Gibson, G.R.; Rastall, R.A. Influence of disaccharide structure on prebiotic selectivity in vitro. J. Agric. Food Chem. 2005, 53, 5192–5199. [Google Scholar] [CrossRef] [PubMed]
- Deville, C.; Gharbi, M.; Dandrifosse, G.; Peulen, O. Study on the effects of laminarin, a polysaccharide from seaweed, on gut characteristics. J. Sci. Food Agric. 2007, 87, 1717–1725. [Google Scholar] [CrossRef]
- Martens, E.C.; Lowe, E.C.; Chiang, H.; Pudlo, N.A.; Wu, M.; McNulty, N.P.; Abbott, D.W.; Henrissat, B.; Gilbert, H.J.; Bolam, D.N.; et al. Recognition and degradation of plant cell wall polysaccharides by two human gut symbionts. PLoS Biol. 2011, 9, e1001221. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Duncan, S.H.; Leitch, E.C.M.; Child, M.W.; Flint, H.J. pH and peptide supply can radically alter bacterial populations and short-chain fatty acid ratios within microbial communities from the human colon. Appl. Environ. Microbiol. 2005, 71, 3692–3700. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Kettle, H.; Louis, P.; Holtrop, G.; Duncan, S.H.; Flint, H.J. Modelling the emergent dynamics and major metabolites of the human colonic microbiota. Environ. Microbiol. 2015, 17, 1615–1630. [Google Scholar] [CrossRef] [PubMed]
- Adiotomre, J.; Eastwood, M.A.; Edwards, C.A.; Brydon, W.G. Dietary fiber: In vitro methods that anticipate nutrition and metabolic activity in humans. Am. J. Clin. Nutr. 1990, 52, 128–134. [Google Scholar] [PubMed]
- Khan, K.M.; Edwards, C.A. In vitro fermentation characteristics of a mixture of Raftilose and guar gum by human faecal bacteria. Eur. J. Nutr. 2005, 44, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Morrison, D.J.; Mackay, W.G.; Edwards, C.A.; Preston, T.; Dodson, B.; Weaver, L.T. Butyrate production from oligofructose fermentation by the human faecal flora: What is the contribution of extracellular acetate and lactate? Br. J. Nutr. 2006, 96, 570–577. [Google Scholar] [PubMed]
- Bourriaud, C.; Robins, R.J.; Martin, L.; Kozlowski, F.; Tenailleau, E.; Cherbut, C.; Michel, C. Lactate is mainly fermented to butyrate by human intestinal microfloras but inter-individual variation is evident. J. Appl. Microbiol. 2005, 99, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Polyviou, T.; MacDougall, K.; Chambers, E.S.; Viardot, A.; Psichas, A.; Jawaid, S.; Harris, H.C.; Edwards, C.A.; Simpson, L.; Murphy, K.G.; et al. Randomised clinical study: Inulin short-chain fatty acid esters for targeted delivery of short-chain fatty acids to the human colon. Aliment. Pharmacol. Ther. 2016, 44, 662–672. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, S.J.D.; Li, J.V.; Lahti, L.; Ou, J.; Carbonero, F.; Mohammed, K.; Posma, J.M.; Kinross, J.; Wahl, E.; Ruder, E.; et al. Fat, fibre and cancer risk in African Americans and rural Africans. Nat. Commun. 2015, 6, 6342. [Google Scholar]
- Fässler, C.; Arrigoni, E.; Venema, K.; Brouns, F.; Amadò, R. In vitro fermentability of differently digested resistant starch preparations. Mol. Nutr. Food Res. 2006, 50, 1220–1228. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Rose, D.J.; Rumpagaporn, P.; Patterson, J.A.; Hamaker, B.R. In vitro batch fecal fermentation comparison of gas and short-chain fatty acid production using “slowly fermentable” dietary fibers. J. Food Sci. 2011, 76, H137–H142. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.L.; Timm, D.A.; Slavin, J.L. Fructooligosaccharides exhibit more rapid fermentation than long-chain inulin in an in vitro fermentation system. Nutr. Res. 2008, 28, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Valk, V.; van der Kaaij, R.M.; Dijkhuizen, L. Characterization of the starch-acting MaAmyB enzyme from Microbacterium aurum B8.A representing the novel subfamily GH13_42 with an unusual, multi-domain organization. Sci. Rep. 2016, 6, 36100. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| 0 h | 8 h | 24 h | ||||
|---|---|---|---|---|---|---|
| α | β | α | β | α | β | |
| Diglucose (1-1) | 7.13 (0.14) | 7.08 (0.24) | 5.10 (0.17) a,* | 4.32 (0.25) | 5.51 (0.06) a,* | 3.91 (0.12) |
| Diglucose (1-2) | 7.04 (0.21) | 7.23 (0.18) | 4.43 (0.27) b | 4.29 (0.10) | 3.93 (0.10) b | 3.85 (0.08) |
| Diglucose (1-3) | 7.04 (0.20) | 6.98 (0.22) | 4.18 (0.18) b | 4.50 (0.11) | 3.92 (0.10) b | 4.05 (0.12) b |
| Diglucose (1-4) | 7.06 (0.21) | 7.16 (0.17) | 3.93 (0.09) b,* | 4.49 (0.09) | 3.64 (0.06) b,* | 4.15 (0.14) b |
| Diglucose (1-6) | 7.08 (0.21) | 7.19 (0.08) | 4.11 (0.25) b | 4.24 (0.23) | 3.81 (0.06) b | 3.77 (0.11) a |
| Acetate | Propionate | Butyrate | Total | |||||
|---|---|---|---|---|---|---|---|---|
| Glucose Bond Orientation | α | β | α | β | α | β | α | β |
| Diglucose (1-1) | 1.8 (0.3) a | 2.4 (0.3) | 0.4 (0.1) | 0.1 (0.0) | 0.9 (0.2) a,* | 0.2 (0.1) | 3.0 (0.4) | 2.7 (0.3) |
| Diglucose (1-2) | 2.4 (0.3) | 2.7 (0.3) | 0.1 (0.1) | 0.2 (0.1) | 0.1 (0.1) | 0.5 (0.1) | 2.9 (0.3) | 3.5 (0.5) |
| Diglucose (1-3) | 2.6 (0.3) | 2.3 (0.3) | 0.2 (0.1) | 0.2 (0.1) | 0.2 (0.1) b | 0.3 (0.1) | 3.0 (0.3) | 2.9 (0.4) |
| Diglucose (1-4) | 2.9 (0.3) b | 1.9 (0.2) a | 0.2 (0.1) | 0.4 (0.1) | 0.1 (0.0) b,* | 0.6 (0.1) | 3.2 (0.4) | 2.9 (0.3) |
| Diglucose (1-6) | 2.9 (0.2) b | 3.7 (0.3) b | 0.2 (0.1) | 0.3 (0.1) | 0.2 (0.0) b | 0.4 (0.1) | 3.2 (0.3) | 4.3 (0.3) |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harris, H.C.; Edwards, C.A.; Morrison, D.J. Impact of Glycosidic Bond Configuration on Short Chain Fatty Acid Production from Model Fermentable Carbohydrates by the Human Gut Microbiota. Nutrients 2017, 9, 26. https://doi.org/10.3390/nu9010026
Harris HC, Edwards CA, Morrison DJ. Impact of Glycosidic Bond Configuration on Short Chain Fatty Acid Production from Model Fermentable Carbohydrates by the Human Gut Microbiota. Nutrients. 2017; 9(1):26. https://doi.org/10.3390/nu9010026
Chicago/Turabian StyleHarris, Hannah C., Christine A. Edwards, and Douglas J. Morrison. 2017. "Impact of Glycosidic Bond Configuration on Short Chain Fatty Acid Production from Model Fermentable Carbohydrates by the Human Gut Microbiota" Nutrients 9, no. 1: 26. https://doi.org/10.3390/nu9010026
APA StyleHarris, H. C., Edwards, C. A., & Morrison, D. J. (2017). Impact of Glycosidic Bond Configuration on Short Chain Fatty Acid Production from Model Fermentable Carbohydrates by the Human Gut Microbiota. Nutrients, 9(1), 26. https://doi.org/10.3390/nu9010026