Impact of Novel Sorghum Bran Diets on DSS-Induced Colitis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Fecal Collections and Processing
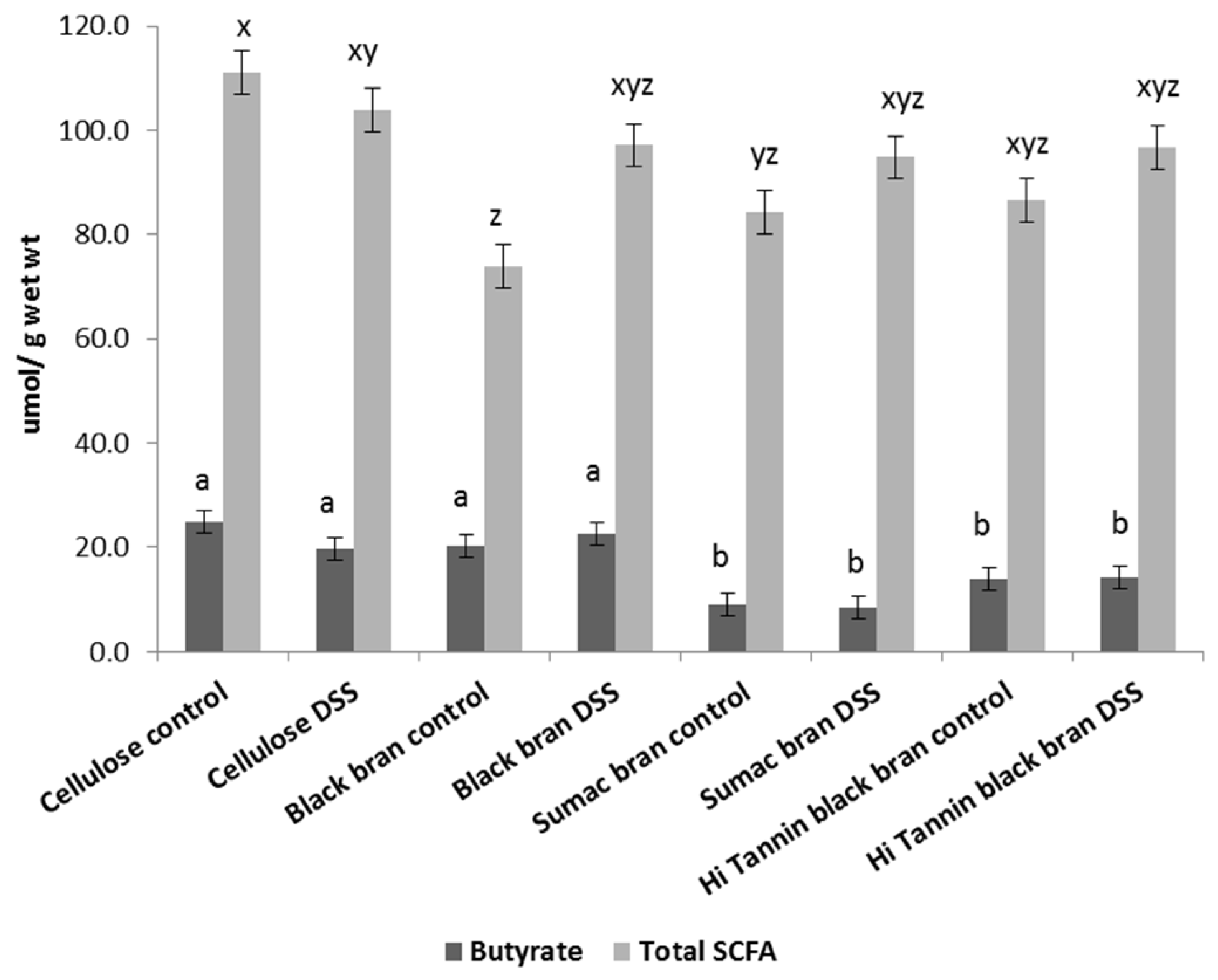
Short Chain Fatty Acid Analysis
2.3. Tissue Fixation for Immunohistological Processing
2.4. Mucosal Samples
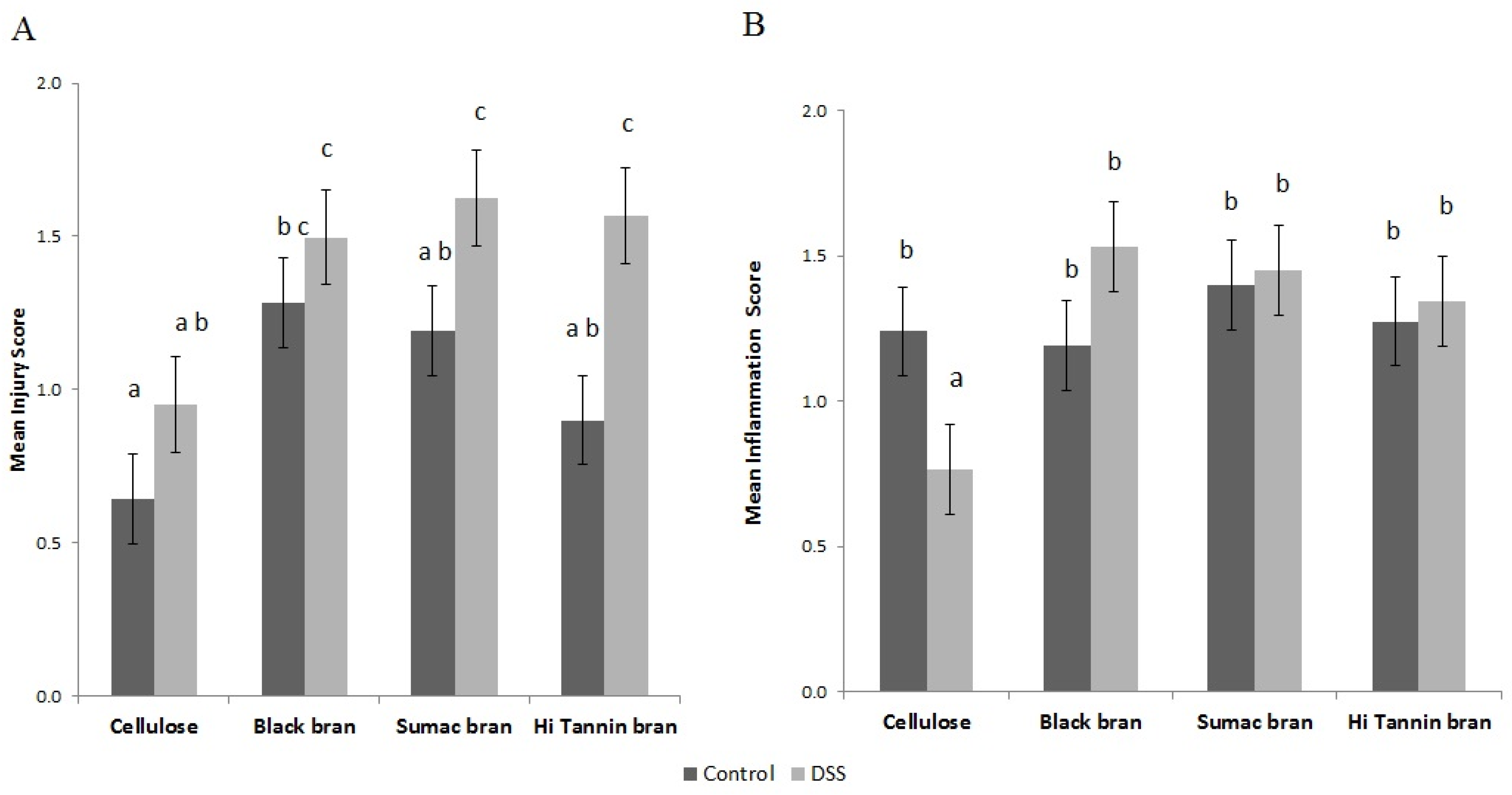
2.5. Inflammation and Injury Histological Scores
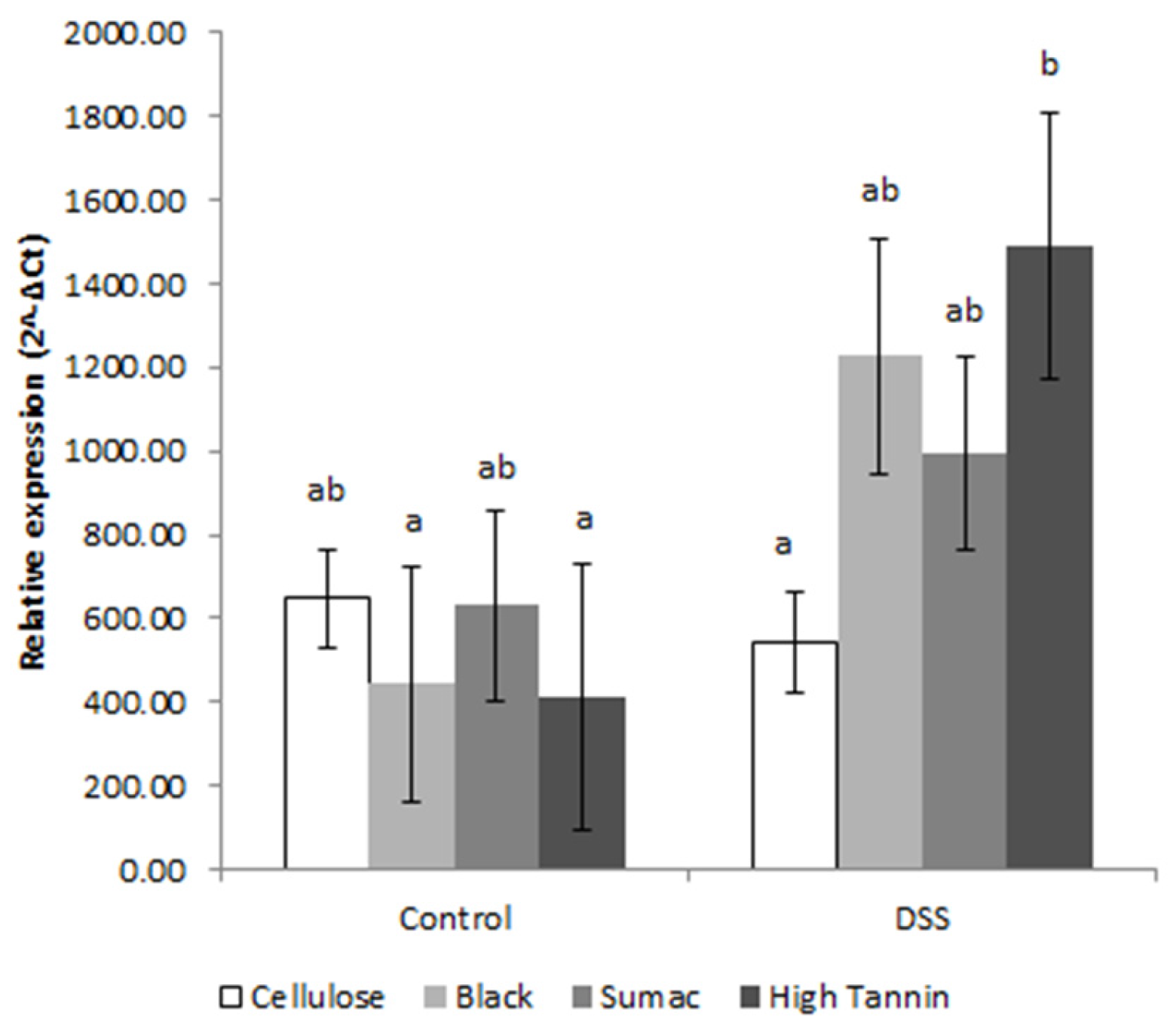
2.6. Activated NF-κB Measurement
2.7. Immunohistological Measurement of Proliferation and Apoptosis
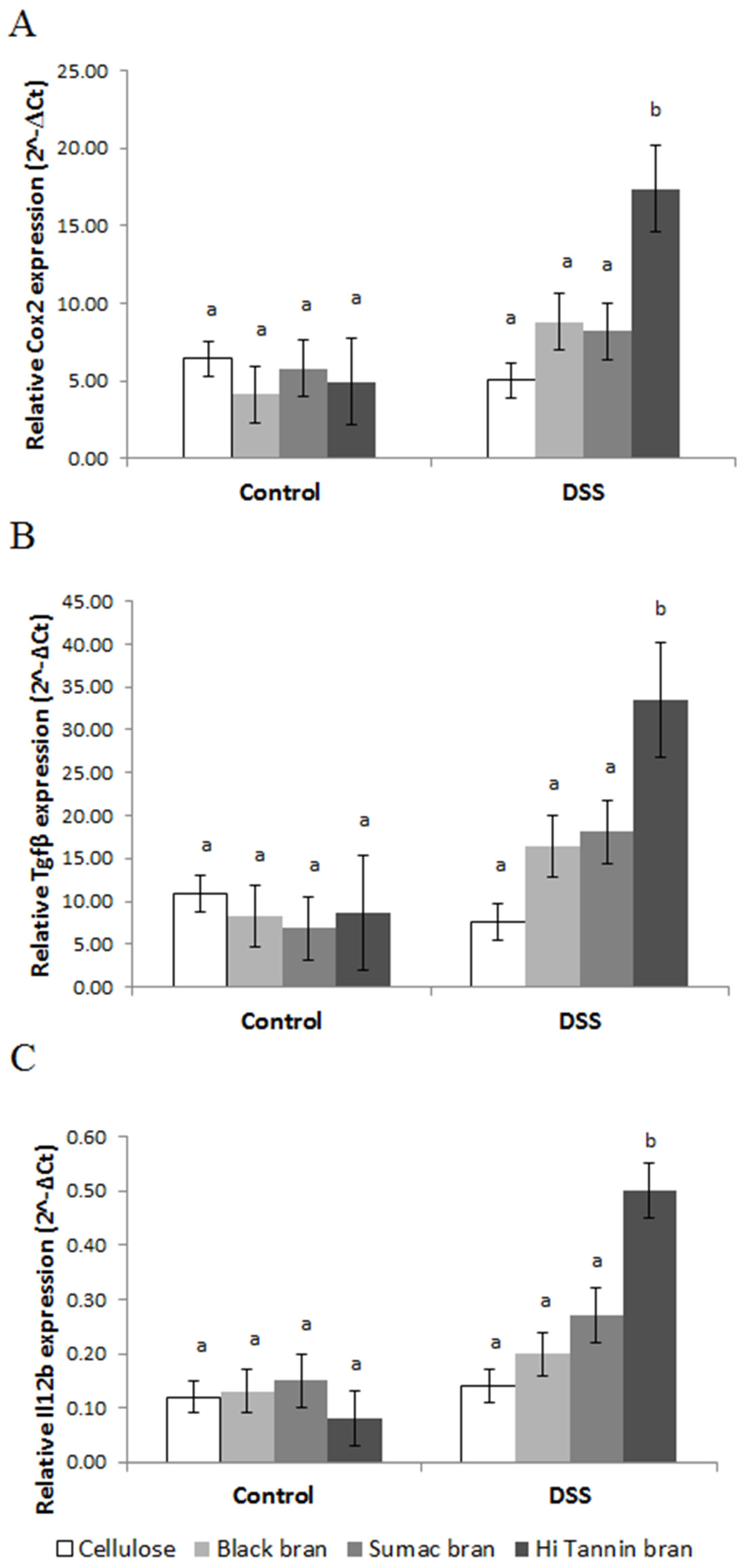
2.8. Measurement of Gene Expression Using Real-Time PCR
2.9. Statistical Analysis
3. Results
3.1. Body Weight, Intake and Fecal Moisture Content
3.2. Immunohistochemical Analysis of Distal Colon
3.3. Fecal SCFA Analysis
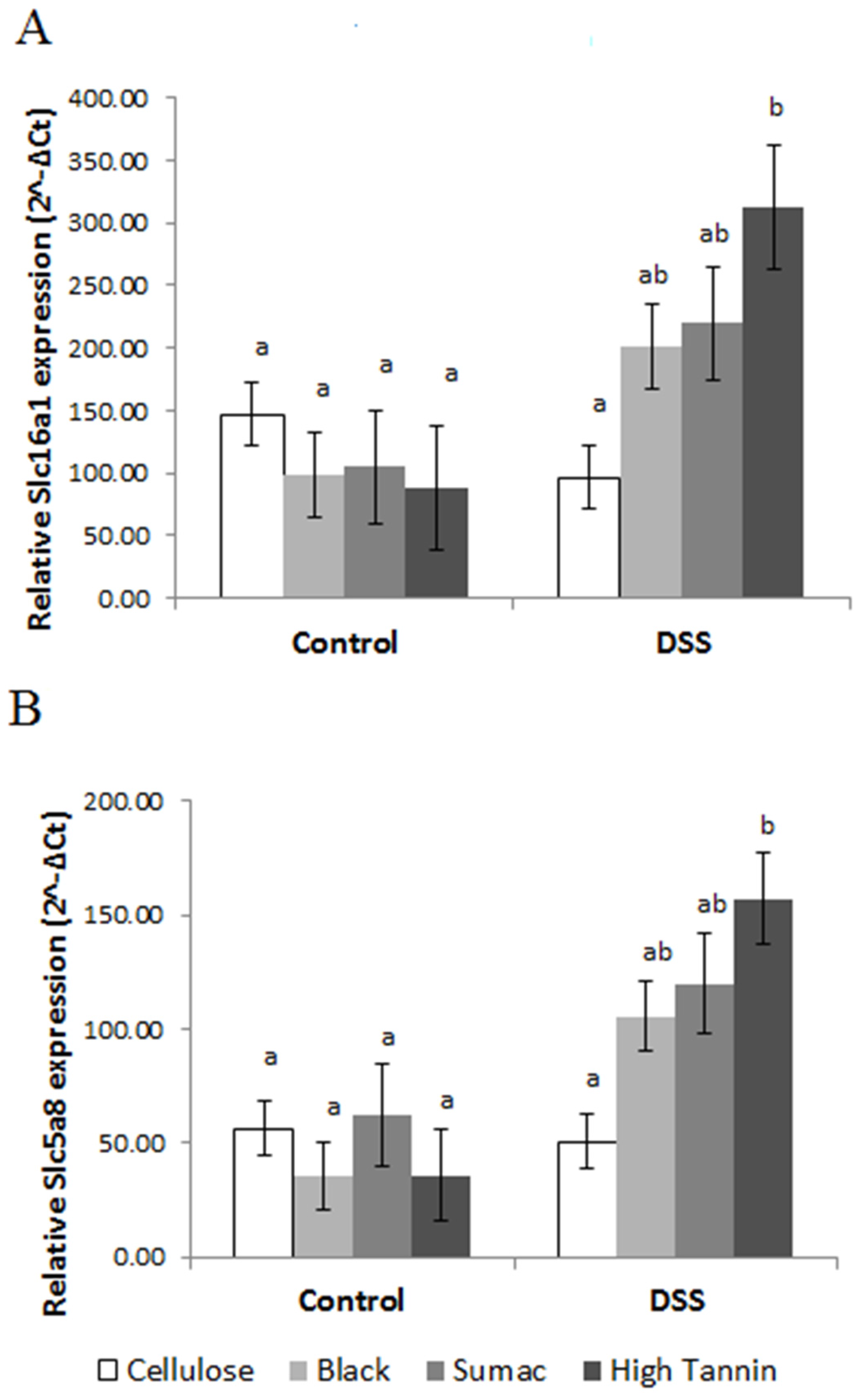
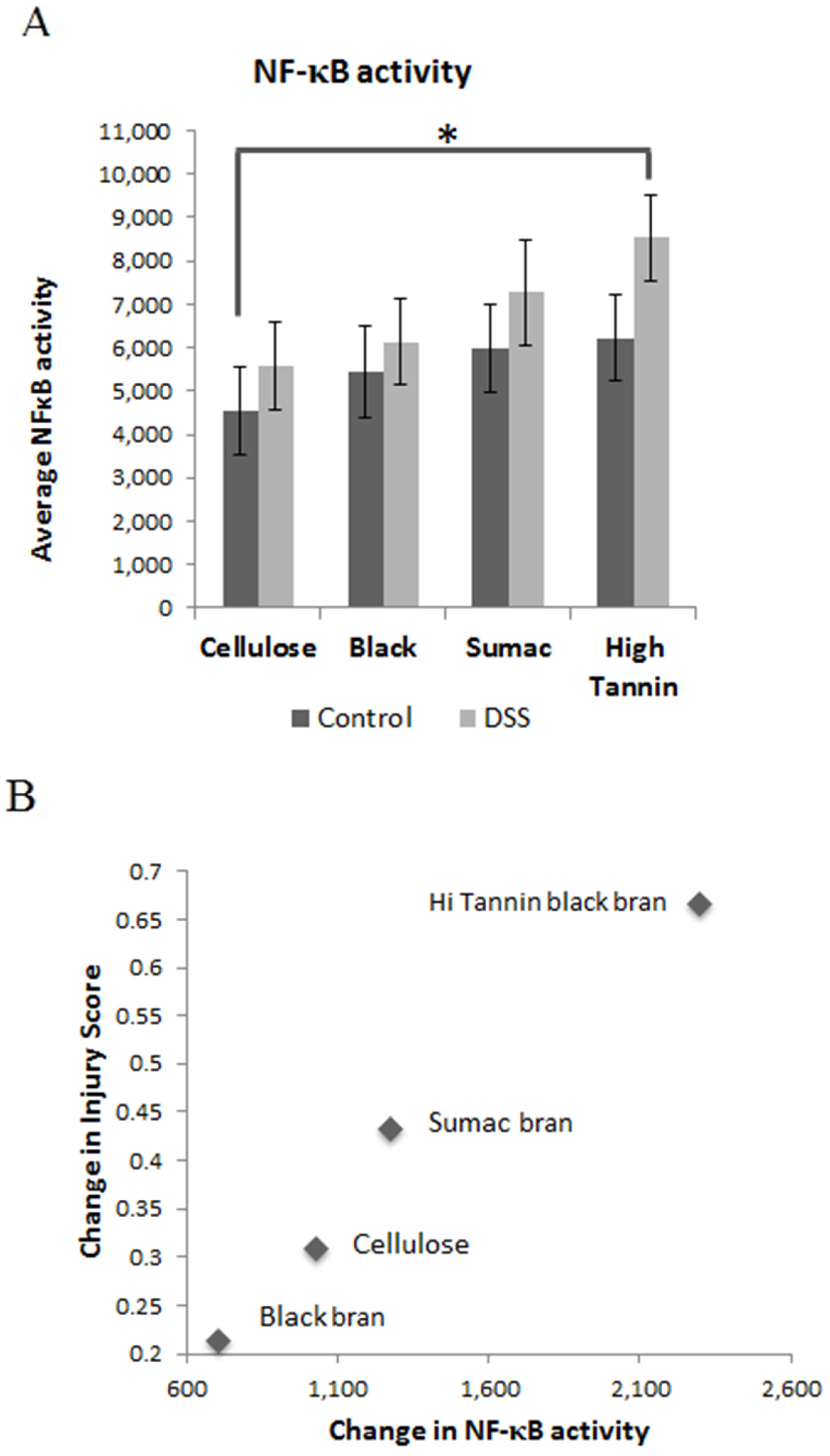
3.4. Mucosal Gene Expression and NF-κB Activity
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| DSS | Dextran sodium sulfate |
| IBD | Inflammatory bowel disease |
| NF-κB | Nuclear factor- κB |
| SCFA | Short chain fatty acids |
| TLR | Toll-like receptor |
| UC | Ulcerative colitis |
References
- Loftus, E.V., Jr. Clinical epidemiology of inflammatory bowel disease: Incidence, prevalence, and environmental influences. Gastroenterology 2004, 126, 1504–1517. [Google Scholar] [CrossRef] [PubMed]
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel disease. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, P.L.; Fischer, S.; Lakatos, L.; Gal, I.; Papp, J. Current concept on the pathogenesis of inflammatory bowel disease-crosstalk between genetic and microbial factors: Pathogenic bacteria and altered bacterial sensing or changes in mucosal integrity take “toll”? World J. Gastroenterol. 2006, 12, 1829–1841. [Google Scholar] [CrossRef] [PubMed]
- Cario, E. Bacterial interactions with cells of the intestinal mucosa: Toll-like receptors and NOD2. Gut 2005, 54, 1182–1193. [Google Scholar] [CrossRef] [PubMed]
- Kanauchi, O.; Matsumoto, Y.; Matsumura, M.; Fukuoka, M.; Bamba, T. The Beneficial Effects of Microflora, Especially Obligate Anaerobes, and Their Products on the Colonic Environment in Inflammatory Bowel Disease. Curr. Pharm. Des. 2005, 11, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Latella, G.; Caprilli, R. Metabolism of large bowel mucosa in health and disease. Int. J. Colorectal Dis. 1991, 6, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Topping, D.L.; Clifton, P.M. Short-Chain Fatty Acids and Human Colonic Function: Roles of Resistant Starch and Nonstarch Polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [PubMed]
- Semann, M.; Bhmig, G.; Zlabinger, G. Short-chain fatty acids: Bacterial mediators of a balanced host-microbial relationship in the human gut. Wien. Klin. Wochenschr. 2002, 114, 289–300. [Google Scholar]
- Crim, K.C.; Sanders, L.M.; Hong, M.Y.; Taddeo, S.S.; Turner, N.D.; Chapkin, R.S.; Lupton, J.R. Upregulation of p21Waf1/Cip1 expression in vivo by butyrate administration can be chemoprotective or chemopromotive depending on the lipid component of the diet. Carcinogenesis 2008, 29, 1415–1420. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Turner, N.D.; Davidson, L.A.; Chapkin, R.S.; Carroll, R.J.; Lupton, J.R. A chemoprotective fish oil/pectin diet enhances apoptosis via Bcl-2 promoter methylation in rat azoxymethane-induced carcinomas. Exp. Biol. Med. 2012, 237, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.; Turner, N.D.; Davidson, L.A.; Chapkin, R.S.; Carroll, R.J.; Lupton, J.R. Colon cancer cell apoptosis is induced by combined exposure to the n-3 fatty acid docosahexaenoic acid and butyrate through promoter methylation. Exp. Biol. Med. 2014, 239, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Toll-like receptors in innate immunity. Int. Immunol. 2005, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Loncar, M.B.; Al-azzeh, E.-D.; Sommer, P.S.M.; Marinovic, M.; Schmehl, K.; Kruschewski, M.; Blin, N.; Stohwasser, R.; Gött, P.; Kayademir, T. Tumour necrosis factor α and nuclear factor κB inhibit transcription of human TFF3 encoding a gastrointestinal healing peptide. Gut 2003, 52, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Taupin, D.R. Intestinal Trefoil Factor Confers Colonic Epithelial Resistance to Apoptosis. Proc. Natl. Acad. Sci. USA 2000, 97, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.W.; Ince, J.; Duncan, S.H.; Webster, L.M.; Holtrop, G.; Ze, X.; Brown, D.; Stares, M.D.; Scott, P.; Bergerat, A. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2010, 5, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The effect of diet on the human gut microbiome: A metagenomic analysis in humanized gnotobiotic mice. Sci. Transl. Med. 2009, 1, 6ra14. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; Luceri, C.; Vivoli, E.; Pagliuca, C.; Lodovici, M.; Moneti, G.; Dolara, P. Polyphenol metabolites from colonic microbiota exert anti-inflammatory activity on different inflammation models. Mol. Nutr. Food Res. 2009, 53, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- Van Duynhoven, J.; Vaughan, E.E.; Jacobs, D.M.; Kemperman, R.A.; van Velzen, E.J.J.; Gross, G.; Roger, L.C.; Possemiers, S.; Smilde, A.K.; Doré, J.; et al. Metabolic fate of polyphenols in the human superorganism. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4531–4538. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, L.E.; Sturino, J.M.; Carroll, R.J.; Rooney, L.W.; Azcarate-Peril, M.A.; Turner, N.D. Polyphenol-rich sorghum brans alter colon microbiota and impact species diversity and species richness after multiple bouts of dextran sodium sulfate-induced colitis. FEMS Microbiol. Ecol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Klinder, A.; Fava, F.; Napolitano, A.; Fogliano, V.; Leonard, C.; Gibson, G.R.; Tuohy, K.M. Whole-grain wheat breakfast cereal has a prebiotic effect on the human gut microbiota: A double-blind, placebo-controlled, crossover study. Br. J. Nutr. 2008, 99, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Tap, J.; Furet, J.P.; Bensaada, M.; Philippe, C.; Roth, H.; Rabot, S.; Lakhdari, O.; Lombard, V.; Henrissat, B.; Corthier, G. Gut microbiota richness promotes its stability upon increased dietary fibre intake in healthy adults. Environ. Microbiol. 2015, 17, 4954–4964. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.T.; Lu, Z.; Chou, M.W. Mechanism of inhibition of tannic acid and related compounds on the growth of intestinal bacteria. Food Chem. Toxicol. 1998, 36, 1053–1060. [Google Scholar] [CrossRef]
- Awika, J.M.; McDonough, C.M.; Rooney, L.W. Decorticating sorghum to concentrate healthy phytochemicals. J. Agric. Food Chem. 2005, 53, 6230. [Google Scholar] [CrossRef] [PubMed]
- Dykes, L.; Rooney, W.L.; Rooney, L.W. Evaluation of phenolics and antioxidant activity of black sorghum hybrids. J. Cereal Sci. 2013, 58, 278–283. [Google Scholar] [CrossRef]
- Dykes, L.; Rooney, L.W. Sorghum and millet phenols and antioxidants. J. Cereal Sci. 2006, 44, 236–251. [Google Scholar] [CrossRef]
- DeVries, J.W.; Prosky, L. Dietary Fiber Analysis and Applications; Aoac Intl: Gaithersburg, MD, USA, 1997. [Google Scholar]
- Seril, D.N.; Liao, J.; Yang, G.-Y.; Yang, C.S. Oxidative stress and ulcerative colitis-associated carcinogenesis: Studies in humans and animal models. Carcinogenesis 2003, 24, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Zoran, D.L.; Turner, N.D.; Taddeo, S.S.; Chapkin, R.S.; Lupton, J.R. Wheat Bran Diet Reduces Tumor Incidence in a Rat Model of Colon Cancer Independent of Effects on Distal Luminal Butyrate Concentrations. J. Nutr. 1997, 127, 2217–2225. [Google Scholar] [PubMed]
- Leonardi, T.; Vanamala, J.; Taddeo, S.S.; Davidson, L.A.; Murphy, M.E.; Patil, B.S.; Wang, N.; Carroll, R.J.; Chapkin, R.S.; Lupton, J.R. Apigenin and naringenin suppress colon carcinogenesis through the aberrant crypt stage in azoxymethane-treated rats. Exp. Biol. Med. 2010, 235, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.A.; Paulhill, K.J.; Davidson, L.A.; Lupton, J.R.; Taddeo, S.S.; Hong, M.Y.; Carroll, R.J.; Chapkin, R.S.; Turner, N.D. Quercetin may suppress rat aberrant crypt foci formation by suppressing inflammatory mediators that influence proliferation and apoptosis. J. Nutr. 2009, 139, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Lupton, J.R.; Smith, R.; Weeks, B.R.; Callaway, E.; Davidson, L.A.; Kim, W.; Fan, Y.-Y.; Yang, P.; Newman, R.A.; et al. Reduced Colitis-Associated Colon Cancer in Fat-1 (n-3 Fatty Acid Desaturase) Transgenic Mice. Cancer Res. 2008, 68, 3985–3991. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Ivanov, I.; Zlatev, Z.Z.; Alaniz, R.C.; Weeks, B.R.; Callaway, E.S.; Goldsby, J.S.; Davidson, L.A.; Fan, Y.-Y.; Zhou, L.; et al. Dietary fish oil and curcumin combine to modulate colonic cytokinetics and gene expression in dextran sodium sulphate-treated mice. Br. J. Nutr. 2011, 106, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Vanamala, J. Suppression of colon carcinogenesis by bioactive compounds in grapefruit. Carcinogenesis 2006, 27, 1257. [Google Scholar] [CrossRef] [PubMed]
- Hijova, E.; Chmelarova, A. Short chain fatty acids and colonic health. Bratisl. Lek. Listy 2007, 108, 354. [Google Scholar] [PubMed]
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; et al. Increasing Incidence and Prevalence of the Inflammatory Bowel Diseases With Time, Based on Systematic Review. Gastroenterology 2012, 142. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.W.; Baird, P.; Davis, R.H., Jr.; Ferreri, S.; Knudtson, M.; Koraym, A.; Waters, V.; Williams, C.L. Health benefits of dietary fiber. Nutr. Rev. 2009, 67, 188–205. [Google Scholar] [CrossRef] [PubMed]
- Perse, M.; Cerar, A. Dextran Sodium Sulphate Colitis Mouse Model: Traps and Tricks. J. Biomed. Biotechnol. 2012. [Google Scholar] [CrossRef]
- Rose, D.J.; DeMeo, M.T.; Keshavarzian, A.; Hamaker, B.R. Influence of dietary fiber on inflammatory bowel disease and colon cancer: Importance of fermentation pattern. Nutr. Rev. 2007, 65, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Kanauchi, O.; Serizawa, I.; Araki, Y.; Suzuki, A.; Andoh, A.; Fujiyama, Y.; Mitsuyama, K.; Takaki, K.; Toyonaga, A.; Sata, M. Germinated barley foodstuff, a prebiotic product, ameliorates inflammation of colitis through modulation of the enteric environment. J. Gastroenterol. 2003, 38, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Kanauchi, O.; Iwanaga, T.; Andoh, A.; Araki, Y.; Nakamura, T.; Mitsuyama, K.; Suzuki, A.; Hibi, T.; Bamba, T. Dietary fiber fraction of germinated barley foodstuff attenuated mucosal damage and diarrhea, and accelerated the repair of the colonic mucosa in an experimental colitis. J. Gastroenterol. Hepatol. 2001, 16, 160–168. [Google Scholar] [CrossRef]
- Rodríguez-Cabezas, M.E.; Gálvez, J.; Lorente, M.D.; Concha, A.; Camuesco, D.; Azzouz, S.; Osuna, A.; Redondo, L.; Zarzuelo, A. Dietary Fiber Down-Regulates Colonic Tumor Necrosis Factor α and Nitric Oxide Production in Trinitrobenzenesulfonic Acid-Induced Colitic Rats. J. Nutr. 2002, 132, 3263–3271. [Google Scholar] [PubMed]
- Piefer, L.A.; Stehm, R.; Krenek, K.; Weeks, B.; Carroll, J.; Byrne, D.; Talcott, S.; Turner, N. Chlorogenic acid reduced DSS-induced injury and NF-kappa B activation in a rat colitis model. FASEB J. 2011, 25, 773.13. [Google Scholar]
- Austin, D.L.; Turner, N.D.; McDonough, C.M.; Rooney, L.W. Effects of Brans from Specialty Sorghum Varieties on In Vitro Starch Digestibility of Soft and Hard Sorghum Endosperm Porridges. Cereal Chem. 2012, 89, 190–197. [Google Scholar] [CrossRef]
- Xiang, J.Y.; Wu, L.G.; Huang, X.L.; Zhang, M.; Pen, L.; Ouyan, Q.; Gan, H.T. Amelioration of murine dextran sulfate sodium-induced colitis by nuclear factor-κb decoy oligonucleotides. Am. J. Surg. 2009, 197, 797–805. [Google Scholar] [CrossRef] [PubMed]
- Osman, N.; Adawi, D.; Ahrné, S.; Jeppsson, B.; Molin, G. Probiotics and blueberry attenuate the severity of dextran sulfate sodium (DSS)-induced colitis. Dig. Dis. Sci. 2008, 53, 2464–2473. [Google Scholar] [CrossRef] [PubMed]
- Paturi, G.; Paturi, T.; Mandimika, C.; Butts, S.; Zhu, N.; Roy, W.; McNabb, J. Influence of dietary blueberry and broccoli on cecal microbiota activity and colon morphology in mdr1 −/− mice, a model of inflammatory bowel diseases. Nutrition 2012, 28, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Shao, J.; Dixon, D.A.; Williams, C.S.; Prescott, S.M.; DuBois, R.N.; Beauchamp, R.D. Transforming growth factor-β1 enhances Ha-ras-induced expression of cyclooxygenase-2 in intestinal epithelial cells via stabilization of mRNA. J. Biol. Chem. 2000, 275, 6628–6635. [Google Scholar] [CrossRef] [PubMed]
- Podolsky, D.K.; Gerken, G.; Eyking, A.; Cario, E. Colitis-associated variant of TLR2 causes impaired mucosal repair because of TFF3 deficiency. Gastroenterology 2009, 137, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.H.; Mackie, R.I. Effect of Condensed Tannins on Bacterial Diversity and Metabolic Activity in the Rat Gastrointestinal Tract. Appl. Environ. Microbiol. 2004, 70, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.N.; St. Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.S. Butyrate and glucose metabolism by colonocytes in experimental colitis in mice. Gut 2000, 46, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Donohoe, D.R.; Collins, L.B.; Wali, A.; Bigler, R.; Sun, W.; Bultman, S.J. The Warburg Effect Dictates the Mechanism of Butyrate-Mediated Histone Acetylation and Cell Proliferation. Mol. Cell 2012, 48, 612–626. [Google Scholar] [CrossRef] [PubMed]
- De Preter, V.; Arijs, I.; Windey, K.; Vanhove, W.; Vermeire, S.; Schuit, F.; Rutgeerts, P.; Verbeke, K. Impaired butyrate oxidation in ulcerative colitis is due to decreased butyrate uptake and a defect in the oxidation pathway. Inflamm. Bowel Dis. 2012, 18, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Thibault, R.; Thibault, P.; de Coppet, K.; Daly, A.; Bourreille, M.; Cuff, C.; Bonnet, J.O.; Mosnier, J.; Galmiche, S.; ShiraziBeechey, J. Down-Regulation of the Monocarboxylate Transporter 1 Is Involved in Butyrate Deficiency During Intestinal Inflammation. Gastroenterology 2007, 133, 1916–1927. [Google Scholar] [CrossRef] [PubMed]
- De Preter, V.; Vandermeulen, G.; Rutgeerts, P.J.; Verbeke, K. T1795 Impaired Butyrate Oxidation in Ulcerative Colitis is Due to Decreased Butyrate Uptake and a Defect in the Oxidation Pathway. Gastroenterology 2010, 138 (Suppl. 1), S580. [Google Scholar] [CrossRef]
- Wullaert, A. Role of NF-κB activation in intestinal immune homeostasis. Int. J. Med. Microbiol. 2010, 300, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Cava, C.F.; Ishihara, S.; Rumi, M.A.K.; Kawashima, K.; Ishimura, N.; Kazumori, H.; Udagawa, J.; Kadowaki, Y.; Kinoshita, Y. Strategic Compartmentalization of Toll-Like Receptor 4 in the Mouse Gut. J. Immunol. 2003, 170, 3977–3985. [Google Scholar] [CrossRef] [PubMed]
- Fukata, M.; Fukata, A.; Chen, A.; Klepper, S.; Krishnareddy, A.; Vamadevan, L.; Thomas, R.; Xu, H.; Inoue, M.; Arditi, A.; et al. Cox-2 Is Regulated by Toll-Like Receptor-4 (TLR4) Signaling: Role in Proliferation and Apoptosis in the Intestine. Gastroenterology 2006, 131, 862–877. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient/Composition | Cellulose | Black Bran | Sumac Bran | Hi Tannin Black Bran |
|---|---|---|---|---|
| INGREDIENT (g/kg) 1 | ||||
| Dextrose | 510.6 | 459.3 | 441.8 | 425.7 |
| Casein | 223.5 | 208.9 | 205.8 | 206.6 |
| DL-methionine | 3.4 | 3.4 | 3.4 | 3.4 |
| Cellulose | 60.0 | 0.00 | 0.00 | 0.00 |
| Sorghum bran | 0.00 | 130.3 | 162.4 | 174.9 |
| Corn oil | 150.0 | 145.8 | 134.6 | 137.1 |
| Mineral mix, AIN-76A | 39.1 | 39.0 | 38.8 | 38.9 |
| Vitamin mix AIN-76A | 11.2 | 11.2 | 11.1 | 11.2 |
| Choline bitartrate | 2.2 | 2.2 | 2.2 | 2.2 |
| BRAN MACRONUTRIENT COMPOSITION (%) * | ||||
| Protein | - | 12.17 | 11.24 | 10.18 |
| Lipid | - | 3.22 | 10.04 | 7.83 |
| Fiber | - | 52.98 | 41.58 | 38.31 |
| Ash | - | 3.37 | 4.94 | 2.81 |
| Carbohydrate | - | 14.87 | 20.44 | 30.04 |
| Moisture | - | 13.39 | 11.76 | 10.83 |
| FIBER CHARACTERIZATION | ||||
| Soluble fiber, % | 6.0 | 6.3 | 11.7 | 10.0 |
| Insoluble fiber, % | 94.0 | 94.0 | 88.3 | 90.0 |
| ANTIOXIDANT CAPACITY AND PHENOLS | ||||
| ABTS † | ND | 64.2 | 182.4 | 136.3 |
| Total phenols ‡ | ND | 5.4 | 11.6 | 8.8 |
| Tannins ‡ | ND | 0.2 | 14.0 | 13.5 |
| Time Relative to DSS | Sample Day | Cellulose | Black Bran | Sumac Bran | Hi Tannin Black Bran | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | DSS | Control | DSS | Control | DSS | Control | DSS | Diet | DSS | Diet*DSS | ||
| Pre DSS#1 | 38 | 39.81 ± 0.89 a | 38.87 ± 1.00 a | 47.60 ± 0.49 b | 46.47 ± 0.75 b | 52.25 ± 0.57 c | 53.05 ± 0.43 c | 48.40 ± 0.30 b | 47.89 ± 0.34 b | <0.0001 | ||
| Post DSS#1 | 44 | 38.68 ± 0.83 a | 39.98 ± 0.75 a | 47.74 ± 0.20 b | 47.74 ± 0.20 b | 50.28 ± 0.55 c | 54.98 ± 1.43 d | 49.43 ± 0.92 b,c | 48.90 ± 0.65 b,c | <0.0001 | 0.0163 | 0.006 |
| Post DSS#2 | 58 | 40.41 ± 0.89 a | 41.53 ± 0.42 a | 47.91 ± 0.21 b | 48.28 ± 0.62 b | 51.41 ± 1.03 c | 53.79 ± 0.76 d | 49.63 ± 0.56 b,c | 50.34 ± 0.77 c | <0.0001 | 0.0241 | |
| Post DSS#3 | 81 | 43.25 ± 1.11 a | 40.89 ± 0.47 a | 48.05 ± 0.51 b | 49.57 ± 0.41 b,c | 53.23 ± 0.54 d | 55.61 ± 0.89 d | 50.06 ± 0.39 c | 49.97 ± 0.51 c | <0.0001 | 0.003 | |
| Variable | Cellulose | Black Bran | Sumac Bran | Hi Tannin Black Bran | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | DSS | Control | DSS | Control | DSS | Control | DSS | Diet | DSS | Diet*DSS | |
| Proliferative index | 9.59 ± 2.88 a,b | 9.35 ± 3.44 a,b | 8.15 ± 3.20 a | 11.18 ± 3.00 b | 9.83 ± 3.15 a,b | 10.75 ± 3.79 b | 9.17 ± 3.02 a,b | 10.89 ± 3.35 b | 0.0112 | ||
| Proliferative zone | 36.96 ± 6.54 b | 34.45 ± 6.50 a,b | 31.49 ± 5.39 a | 35.18 ± 5.08 a,b | 36.29 ± 7.26 a,b | 37.52 ± 5.57 b | 34.91 ± 5.95 a,b | 37.72 ± 7.04 b | |||
| Apoptotic index | 0.18 ± 0.01 a,b | 0.18 ± 0.01 a,b | 0.15 ± 0.01 a,b | 0.18 ± 0.01 a,b | 0.29 ± 0.01 b | 0.22 ± 0.01 a,b | 0.23 ± 0.01 a,b | 0.09 ± 0.01 a | |||
| Crypt height | 30.00 ± 0.74 b | 27.73 ± 0.76 a | 30.74 ± 0.77 b | 26.62 ± 0.88 a | 32.02 ± 0.72 b | 27.08 ± 1.01 a | 31.23 ± 0.68 b | 26.29 ± 0.98 a | <0.0001 | ||
| SCFA | Cellulose | Black Bran | Sumac Bran | Hi Tannin Black Bran | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | DSS | Control | DSS | Control | DSS | Control | DSS | Diet | DSS | Diet*DSS | ||
| Day 38 | Acetic | 43.01 ± 6.70 a | 32.55 ± 6.34 a,b,c | 29.63 ± 2.76 b,c | 26.95 ± 2.16 c | 32.52 ± 2.97 a,b,c | 39.96 ± 5.52 a,b | 28.52 ± 2.05 c | 25.94 ± 2.59 c | 0.0299 | ||
| Propionic | 8.72 ± 0.92 c | 7.85 ± 1.11 c | 11.78 ± 1.22 b | 11.27 ± 1.20 b,c | 18.53 ± 1.42 a | 17.60 ± 1.82 a | 13.64 ± 0.86 b | 12.41 ± 1.01 b | 0.0001 | |||
| Isobutyric | 1.44 ± 0.16 b | 1.35 ± 0.13 b | 1.49 ± 0.13 b | 1.48 ± 0.15 b | 2.91 ± 0.30 a | 2.82 ± 0.28 a | 3.16 ± 0.18 a | 3.02 ± 0.27 a | 0.0001 | |||
| Butyric | 13.34 ± 2.54 a,b | 12.79 ± 2.30 a,b | 16.13 ± 1.17 a | 14.36 ± 1.08 a,b | 8.15 ± 0.77 c | 8.18 ± 0.63 c | 12.99 ± 0.85 ab | 11.84 ± 1.16 b,c | 0.0001 | |||
| Isovaleric | 2.89 ± 0.39 b | 2.74 ± 0.28 b | 3.70 ± 0.31 b | 3.65 ± 0.36 b | 7.73 ± 0.81 a | 7.49 ± 0.81 a | 8.46 ± 0.50 a | 8.18 ± 0.78 a | 0.0001 | |||
| Valeric | 4.40 ± 0.40 b | 4.17 ± 0.48 b | 5.60 ± 0.70 b | 5.55 ± 1.04 b | 1.75 ± 0.96 b | 1.50 ± 0.73 b | 8.39 ± 0.99 b | 16.52 ± 7.90 a | 0.0035 | |||
| Total | 73.80 ± 9.62 a | 61.46 ± 9.35 a | 68.33 ± 5.81 a | 63.26 ± 5.52 a | 71.59 ± 4.67 a | 81.32 ± 7.49 a | 75.16 ± 3.67 a | 77.90 ± 8.97 a | ||||
| Day 81 | Acetic | 49.19 ± 6.70 a | 48.45 ± 6.33 a | 26.04 ± 2.76 b | 33.71 ± 2.17 a,b | 40.33 ± 2.98 a,b | 50.24 ± 5.52 a | 51.30 ± 2.05 b | 37.20 ± 2.59 a,b | |||
| Propionic | 10.79 ± 0.92 b | 12.59 ± 1.11 a,b | 9.10 ± 1.22 b | 22.43 ± 1.20 a | 22.40 ± 1.42 a | 23.98 ± 1.82 a | 22.06 ± 0.86 a | 17.14 ± 1.01 a,b | 0.0550 | |||
| Isobutyric | 1.52 ± 0.16 c | 1.53 ± 0.13 c | 1.14 ± 0.13 c | 1.54 ± 0.15 c | 3.75 ± 0.30 b | 3.77 ± 0.28 b | 5.85 ± 0.18 a | 4.53 ± 0.27 a,b | 0.0001 | |||
| Butyric | 19.67 ± 2.54 a | 16.45 ± 2.30 a,b | 15.98 ± 1.17 a,b | 20.07 ± 1.08 a | 9.20 ± 0.77 b | 8.80 ± 0.63 b | 20.24 ± 0.85 a | 14.08 ± 1.16 a,b | 0.0065 | |||
| Isovaleric | 2.83 ± 0.39 c | 2.94 ± 0.28 c | 2.99 ± 0.31 c | 3.78 ± 0.36 c | 9.48 ± 0.81 b | 9.28 ± 0.58 b | 14.91 ± 0.50 a | 11.35 ± 0.78 a,b | 0.0001 | |||
| Valeric | 4.17 ± 0.40 b | 4.46 ± 0.48 b | 3.98 ± 0.70 b | 5.04 ± 1.04 b | 0.71 ± 0.96 b | 0.55 ± 0.73 b | 14.39 ± 0.99 a | 12.11 ± 7.90 a | 0.0001 | |||
| Total | 88.08 ± 9.62 a,b | 86.41 ± 9.35 a,b | 59.22 ± 5.81 a | 86.55 ± 5.53 a,b | 85.88 ± 4.67 a,b | 96.61 ± 7.48 a,b | 128.74 ± 3.67 a | 96.41 ± 8.97 a,b | ||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ritchie, L.E.; Taddeo, S.S.; Weeks, B.R.; Carroll, R.J.; Dykes, L.; Rooney, L.W.; Turner, N.D. Impact of Novel Sorghum Bran Diets on DSS-Induced Colitis. Nutrients 2017, 9, 330. https://doi.org/10.3390/nu9040330
Ritchie LE, Taddeo SS, Weeks BR, Carroll RJ, Dykes L, Rooney LW, Turner ND. Impact of Novel Sorghum Bran Diets on DSS-Induced Colitis. Nutrients. 2017; 9(4):330. https://doi.org/10.3390/nu9040330
Chicago/Turabian StyleRitchie, Lauren E., Stella S. Taddeo, Brad R. Weeks, Raymond J. Carroll, Linda Dykes, Lloyd W. Rooney, and Nancy D. Turner. 2017. "Impact of Novel Sorghum Bran Diets on DSS-Induced Colitis" Nutrients 9, no. 4: 330. https://doi.org/10.3390/nu9040330
APA StyleRitchie, L. E., Taddeo, S. S., Weeks, B. R., Carroll, R. J., Dykes, L., Rooney, L. W., & Turner, N. D. (2017). Impact of Novel Sorghum Bran Diets on DSS-Induced Colitis. Nutrients, 9(4), 330. https://doi.org/10.3390/nu9040330