Gut Microbiota as a Target for Preventive and Therapeutic Intervention against Food Allergy


 and
and 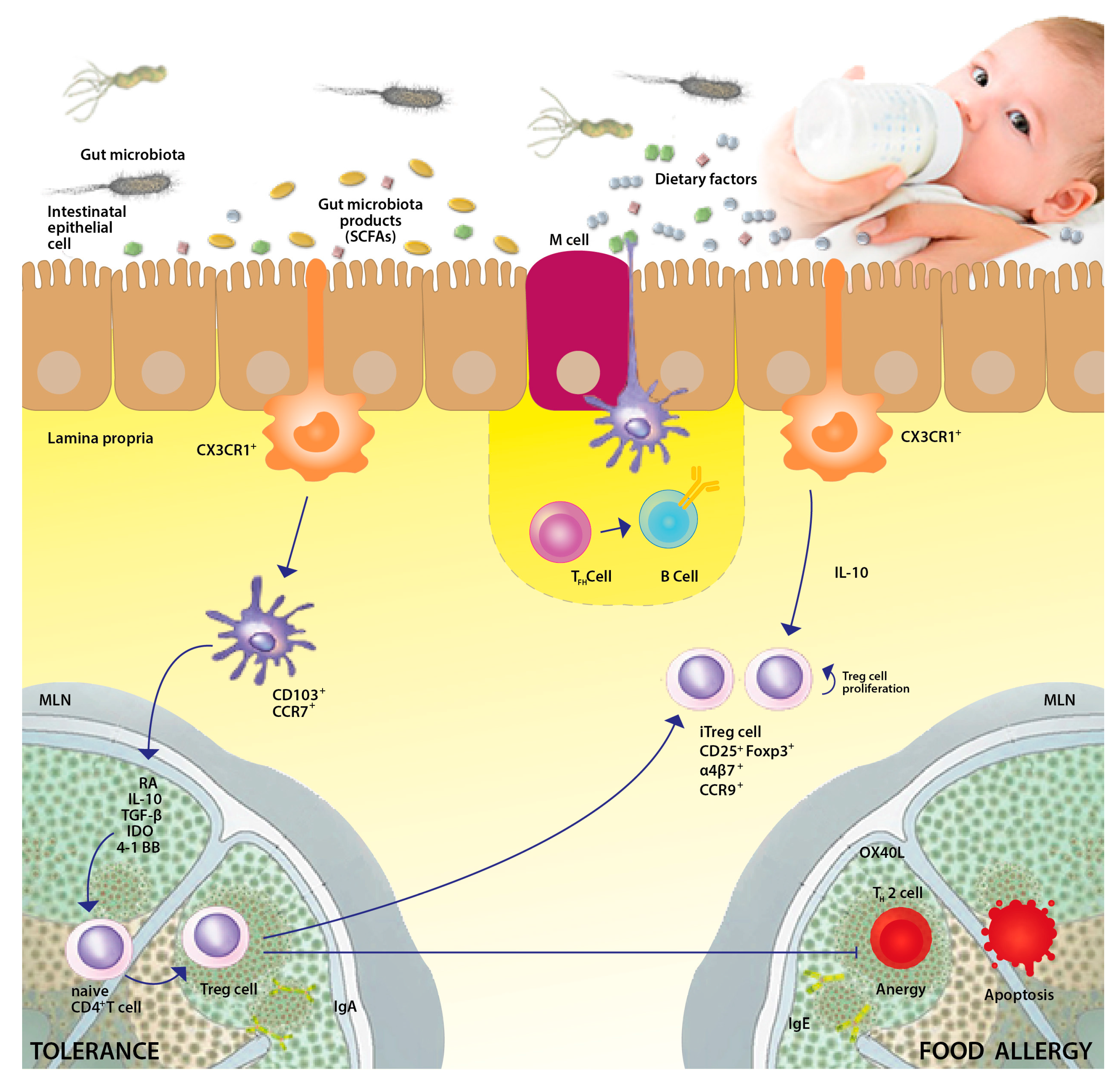{kind=link}
{kind=link}
Abstract
:1. Introduction
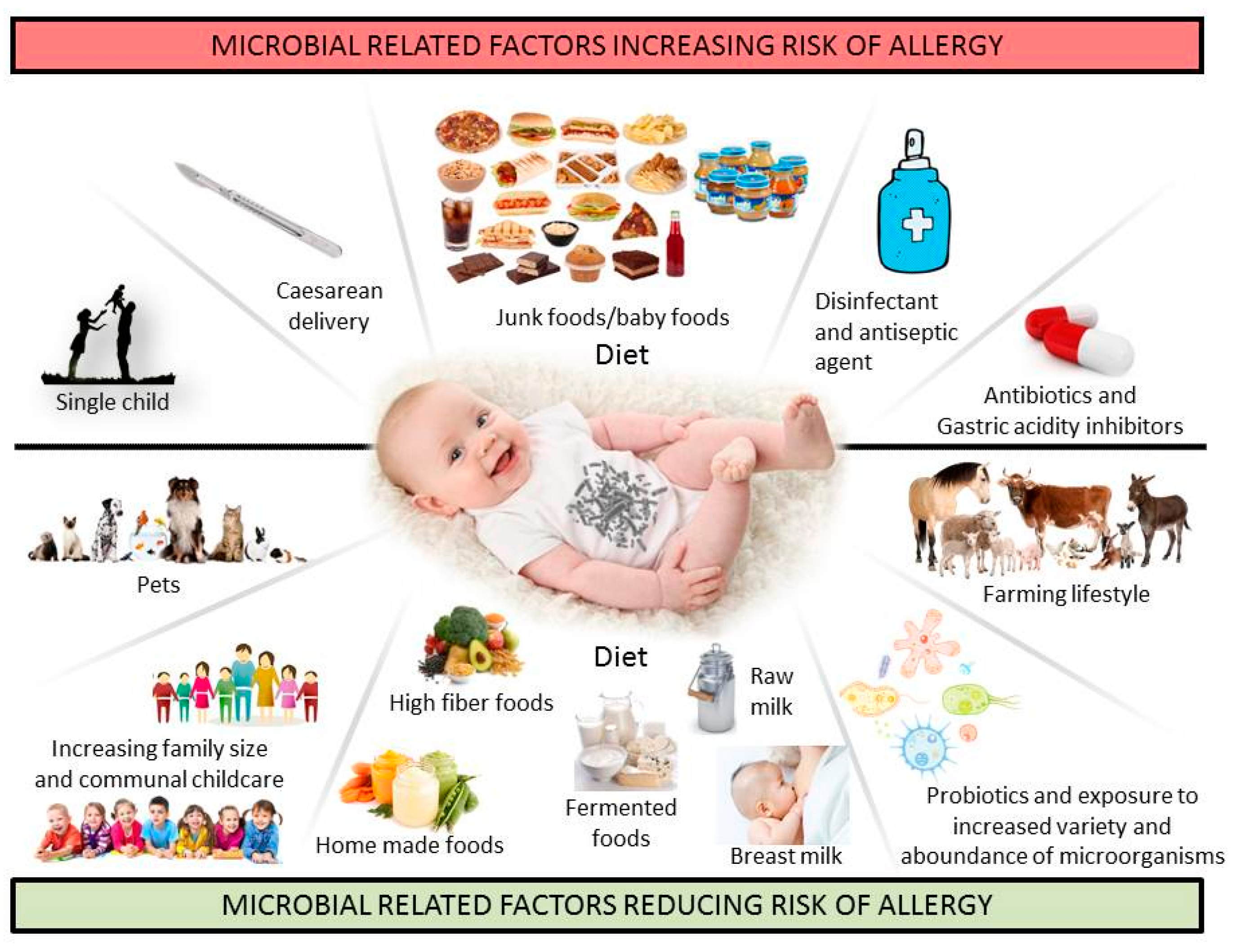
2. Importance of Microbial Exposure for the Development of Immune Tolerance
3. Gut Microbiota in FA
4. Modulation of the Gut Microbiota in FA
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sicherer, S.H.; Sampson, H.A. Food allergy: Epidemiology, pathogenesis, diagnosis, and treatment. J. Allergy Clin. Immunol. 2014, 133, 291–307. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, G.; Tsakok, T. Prevention of food allergy. J. Allergy Clin. Immunol. 2016, 137, 998–1010. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; Quinn, R.A. Microbiome-wide association studies link dynamic microbial consortia to disease. Nature 2016, 535, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Tsakok, T.; Marrs, T. Does atopic dermatitis cause food allergy? A systematic review. J. Allergy Clin. Immunol. 2016, 137, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Hong, S.W. Dietary antigens limit mucosal immunity by inducing regulatory T cells in the small intestine. Science 2016, 351, 858–863. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, G.; Katz, Y. Early consumption of peanuts in infancy is associated with a low prevalence of peanut allergy. J. Allergy Clin. Immunol. 2008, 122, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Koplin, J.J.; Osborne, N.J. Can early introduction of egg prevent egg allergy in infants? A population-based study. J. Allergy Clin. Immunol. 2010, 126, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Katz, Y.; Rajuan, N. Early exposure to cow’s milk protein is protective against IgE-mediated cow’s milk protein allergy. J. Allergy Clin. Immunol. 2010, 126, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, G.; Roberts, G. Randomized trial of peanut consumption in infants at risk for peanut allergy. N. Engl. J. Med. 2015, 372, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, G.; Sayre, P.H.N. Effect of Avoidance on Peanut Allergy after Early Peanut Consumption. N. Engl. J. Med. 2016, 374, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Sekirov, I.; Russell, S.L. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed]
- Tamburini, S.; Shen, N. The microbiome in early life: Implications for health outcomes. Nat. Med. 2016, 22, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Eberl, G. Immunity by equilibrium. Nat. Rev. Immunol. 2016, 16, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Pabst, O.; Mowat, A.M. Oral tolerance to food protein. Mucosal Immunol. 2012, 5, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Stefka, A.T.; Feehley, T. Commensal bacteria protect against food allergen sensitization. Proc. Natl. Acad. Sci. USA 2014, 111, 13145–13150. [Google Scholar] [CrossRef] [PubMed]
- Maynard, C.L.; Elson, C.O. Reciprocal interactions of the intestinal microbiota and immune system. Nature 2012, 489, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Yatsunenko, T.; Rey, F.E. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Tulic, M.K.; Hodder, M. Differences in innate immune function between allergic and non-allergic children: New insights into immune ontogeny. J. Allergy Clin. Immunol. 2011, 127, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Mazmanian, S.K.; Liu, C.H. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; Gilbert, J.A. The role of the commensal microbiota in the regulation of tolerance to dietary antigens. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Ohnmacht, C.; Park, J.-H. The microbiota regulates type 2 immunity through RORgt1 T cells. Science 2015, 349, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Villablanca, E.J. MyD88-dependent TLR1/2 signals educate dendritic cells with gut-specific imprinting properties. J. Immunol. 2011, 187, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Latcham, F. A consistent pattern of minor immunodeficiency and subtle enteropathy in children with multiple food allergy. J. Pediatr. 2003, 143, 39–47. [Google Scholar] [CrossRef]
- Suzuki, K.; Maruya, M. The sensing of environmental stimuli by follicular dendritic cells promotes immunoglobulin A generation in the gut. Immunity 2010, 33, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C. Dietary Fiber and Bacterial SCFA Enhance Oral Tolerance and Protect against Food Allergy through Diverse Cellular Pathways. Cell Rep. 2016, 15, 2809–2824. [Google Scholar] [CrossRef] [PubMed]
- Diesner, S.C.; Knittelfelder, R. Dose-dependent food allergy induction against ovalbumin under acid-suppression: A murine food allergy model. Immunol. Lett. 2008, 121, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Untersmayr, E.; Diesner, S.C. Nitration of the egg-allergen ovalbumin enhances protein allergenicity but reduces the risk for oral sensitization in a murine model of food allergy. PLoS ONE 2010, 5, e14210. [Google Scholar] [CrossRef] [PubMed]
- Diesner, S.C.; Bergmayr, C. A distinct microbiota composition is associated with protection from food allergy in an oral mouse immunization model. Clin. Immunol. 2016, 173, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, K.E.; Sitarik, A.R. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 2016, 22, 1187–1191. [Google Scholar] [CrossRef] [PubMed]
- Olszak, T.; An, D. Microbial exposure during early life has persistent effects on natural killer T cell function. Science 2012, 336, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Marsland, B.J. The Microbiome in Allergic Disease: Current Understanding and Future Opportunities—2017 PRACTALL Document of the American Academy of Allergy, Asthma & Immunology and the European Academy of Allergy and Clinical Immunology. J. Allergy Clin. Immunol. 2017, 139, 1099–1110. [Google Scholar] [PubMed]
- Wesemann, D.R.; Portuguese, A.J. Microbial colonization influences early B-lineage development in the gut lamina propria. Nature 2013, 501, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Rosser, E.C.; Mauri, C. Regulatory B cells: Origin, phenotype, and function. Immunity 2015, 42, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Van de Veen, W.; Stanic, B. IgG4 production is confined to human IL-10-producing regulatory B cells that suppress antigen-specific immune responses. J. Allergy Clin. Immunol. 2013, 131, 1204–1212. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Q.; Wu, Y.; Song, J.P. Tolerogenic CX3CR1+ B cells suppress food allergy-induced intestinal inflammation in mice. Allergy 2013, 68, 1241–1248. [Google Scholar] [CrossRef] [PubMed]
- Amu, S.; Saunders, S.P. Regulatory B cells prevent and reverse allergic airway inflammation via FoxP3-positive T regulatory cells in a murine model. J. Allergy Clin. Immunol. 2010, 125, 1114–1124. [Google Scholar] [CrossRef] [PubMed]
- Tait Wojno, E.D.; Artis, D. Emerging concepts and future challenges in innate lymphoid cell biology. J. Exp. Med. 2016, 213, 2229–2248. [Google Scholar] [CrossRef] [PubMed]
- Eyerich, K.; Dimartino, V. IL-17 and IL-22 in immunity: Driving protection and pathology. Eur. J. Immunol. 2017, 47, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Plunkett, C.H.; Nagler, C.R. The Influence of the Microbiome on Allergic Sensitization to Food. J. Immunol. 2017, 198, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Papathoma, E.; Triga, M. Cesarean section delivery and development of food allergy and atopic dermatitis in early childhood. Pediatr. Allergy Immunol. 2016, 27, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Muraro, A.; Halken, S. EAACI food allergy and anaphylaxis guidelines. Primary prevention of food allergy. Allergy 2014, 69, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Anita Trikha, M.D.; Jacques, G. Baillargeon Development of food allergies in patients with Gastroesophageal Reflux Disease treated with gastric acid suppressive medications. Pediatr Allergy Immunol. 2013, 24, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Grimshaw, K.E.; Maskell, J. Diet and food allergy development during infancy: Birth cohort study findings using prospective food diary data. J. Allergy Clin. Immunol. 2014, 133, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Yamanishi, S. Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences. Cell 2014, 158, 705–721. [Google Scholar] [CrossRef] [PubMed]
- Metsälä, J.; Lundqvist, A. Mother’s and offspring’s use of antibiotics and infant allergy to cow’s milk. Epidemiology 2013, 24, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Marrs, T.; Bruce, K.D. Is there an association between microbial exposure and food allergy? A systematic review. Pediatr. Allergy Immunol. 2013, 24, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Thompson-Chagoyan, O.C.; Vieites, J.M. Changes in faecal microbiota of infants with cow’s milk protein allergy—a Spanish prospective case-control 6-month follow-up study. Pediatr. Allergy Immunol. 2010, 21 Pt 2, e394–e400. [Google Scholar] [CrossRef] [PubMed]
- Adlerberth, I.; Strachan, D.P. Gut microbiota and development of atopic eczema in 3 European birth cohorts. J. Allergy Clin. Immunol. 2007, 120, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Kendler, M.; Uter, W. Comparison of fecal microflora in children with atopic eczema/dermatitis syndrome according to IgE sensitization to food. Pediatr. Allergy Immunol. 2006, 17, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Arrieta, M.C.; Stiemsma, L.T. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci. Transl. Med. 2015, 7, 307ra152. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, J.; Kobayashi, T. Aberrant structures of fecal bacterial community in allergic infants profiled by 16S rRNA gene pyrosequencing. FEMS Immunol. Med. Microbiol. 2011, 63, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Louis, P. The role of pH in determining the species composition of the human colonic microbiota. Environ. Microbiol. 2009, 11, 2112–2122. [Google Scholar]
- Azad, M.B.; Konya, T. Infant gut microbiota and food sensitization: Associations in the first year of life. Clin. Exp. Allergy 2015, 45, 623–643. [Google Scholar] [CrossRef] [PubMed]
- Hua, X.; Goedert, J.J. Allergy associations with the adult fecal microbiota: Analysis of the American Gut Project. EBioMedicine 2015, 3, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Li, Z. Altered fecal microbiota composition associated with food allergy in infants. Appl. Environ. Microbiol. 2014, 80, 2546–2554. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Chen, K.J. Alterations in the gut microbiotas of children with food sensitization in early life. Pediatr. Allergy Immunol. 2016, 27, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Bunyavanich, S.; Shen, N. Early-life gut microbiome composition and milk allergy resolution. J. Allergy Clin. Immunol. 2016, 138, 1122–1130. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.D.; Lawson, P.A. The phylogeny of the genus Clostridium: Proposal of five new genera and eleven new species combinations. Int. J. Syst. Bacteriol. 1994, 44, 812–826. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Stubbington, M.J. The Regulatory T Cell Lineage Factor Foxp3 Regulates Gene Expression through Several Distinct Mechanisms Mostly Independent of Direct DNA Binding. PLoS Genet. 2015, 11, e1005251. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Arpaia, N.; Campbell, C. Metabolites produced by commensal bacteria promote peripheral regulatory T cell generation. Nature 2013, 504, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y. Commensal microbe-derived butyrate induces differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Mackay, C.R. Diet, gut microbiota and immune responses. Nat. Immunol. 2011, 12, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.J.; Zheng, L. Crosstalk between Microbiota-Derived Short-Chain Fatty Acids and Intestinal Epithelial HIF Augments Tissue Barrier Function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Sandin, A.; Bråbäck, L. Faecal short chain fatty acid pattern and allergy in early childhood. Acta Paediatr. 2009, 98, 823–827. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; Sangwan, N. Lactobacillus rhamnosus GG supplemented formula expands butyrate producing bacterial strains in food allergic infants. ISME J. 2016, 10, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Geuking, M.B.; McCoy, K.D. Metabolites from intestinal microbes shape Treg. Cell Res 2013, 23, 1339–1340. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Yoo, Y. Faecalibacterium prausnitzii subspecies-level dysbiosis in the human gut microbiome underlying atopic dermatitis. J. Allergy Clin. Immunol. 2015, 137, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Franzosa, E.A. Relating the metatranscriptome and metagenome of the human gut. Proc. Natl. Acad. Sci. USA 2014, 111, e2329–e2338. [Google Scholar] [CrossRef] [PubMed]
- Sellitto, M. Proof of concept of microbiome–metabolome analysis and delayed gluten exposure on celiac disease autoimmunity in genetically at-risk infants. PLoS ONE 2012, 7, e33387. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.J. Preterm gut microbiota and metabolome following discharge from intensive care. Sci. Rep. 2015, 5, 17141. [Google Scholar] [CrossRef] [PubMed]
- Riedler, J.; Braun-Fahrländer, C. Exposure to farming in early life and development of asthma and allergy: A cross-sectional survey. Lancet 2001, 358, 1129–1133. [Google Scholar] [CrossRef]
- Schuijs, M.J.; Willart, M.A. Farm dust and endotoxin protect against allergy through A20 induction in lung epithelial cells. Science 2015, 349, 1106–1110. [Google Scholar] [CrossRef] [PubMed]
- Dicksved, J.; Flöistrup, H. Molecular fingerprinting of the fecal microbiota of children raised according to different lifestyles. Appl. Environ. Microbiol. 2007, 73, 2284–2289. [Google Scholar] [CrossRef] [PubMed]
- Peters, R.L.; Allen, K.J. Differential factors associated with challenge-proven food allergy phenotypes in a population cohort of infants: A latent class analysis. Clin. Exp. Allergy 2015, 45, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, K.E.; Johnson, C.C. Man’s best friend? The effect of pet ownership on house dust microbial communities. J. Allergy Clin. Immunol. 2010, 126, 410–412. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Peano, C. Diversity of bifidobacteria within the infant gut microbiota. PLoS ONE 2012, 7, e36957. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, H.J.; Wildeboer-Veloo, A.C. Analysis of intestinal flora development in breast-fed and formula-fed infants by using molecular identification and detection methods. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Sitarik, A.R.; Bobbitt, K.R. Breast Milk TGF [beta] is Associated with Neonatal Gut Microbial Composition. J. Pediatr. Gastroenterol. Nutr. 2017. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Osborn, D.A.; Sinn, J.K. Probiotics in infants for prevention of allergic disease and food hypersensitivity. Cochrane Database Syst. Rev. 2007, CD006475. [Google Scholar]
- Hill, C.; Guarner, F. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Fiocchi, A.; Pawankar, R. World Allergy Organization-McMaster University Guidelines for Allergic Disease Prevention [GLAD-P]: Probiotics. WAO J. 2015, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Elazab, N.; Mendy, A. Probiotic administration in early life, atopy, and asthma: A meta-analysis of clinical trials. Pediatrics 2013, 132, e666–e676. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.L.; Ponsonby, A.L. Administration of a probiotic with peanut oral immunotherapy: A randomized trial. J. Allergy Clin. Immunol. 2015, 135, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Hol, J.; van Leer, E.H. The acquisition of tolerance toward cow’s milk through probiotic supplementation: A randomized, controlled trial. J. Allergy Clin. Immunol. 2008, 121, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; Nocerino, R. Effect of Lactobacillus GG on tolerance acquisition in infants with cow’s milk allergy: A randomized trial. J. Allergy Clin. Immunol. 2012, 129, 580–582. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; Nocerino, R. Formula selection for management of children with cow’s milk allergy influences the rate of acquisition of tolerance: A prospective multicenter study. J. Pediatr. 2013, 163, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; Di Costanzo, M. Extensively hydrolyzed casein formula containing Lactobacillus rhamnosus GG reduces the occurrence of other allergic manifestations in children with cow’s milk allergy: 3-year randomized controlled trial. J. Allergy Clin. Immunol. 2017, 139, 1906–1913. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; de Zoeten, E.F. Deacetylase inhibition promotes the generation and function of regulatory T cells. Nat. Med. 2007, 13, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; Paparo, L. Differences in DNA methylation profile of Th1 and Th2 cytokine genes are associated with tolerance acquisition in children with IgE-mediated cow’s milk allergy. Clin. Epigenetics 2015, 31, 7–38. [Google Scholar] [CrossRef] [PubMed]
- Paparo, L.; Nocerino, R. Epigenetic features of FoxP3 in children with cow’s milk allergy. Clin. Epigenetics 2016, 12, 8–86. [Google Scholar] [CrossRef] [PubMed]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aitoro, R.; Paparo, L.; Amoroso, A.; Di Costanzo, M.; Cosenza, L.; Granata, V.; Di Scala, C.; Nocerino, R.; Trinchese, G.; Montella, M.; et al. Gut Microbiota as a Target for Preventive and Therapeutic Intervention against Food Allergy. Nutrients 2017, 9, 672. https://doi.org/10.3390/nu9070672
Aitoro R, Paparo L, Amoroso A, Di Costanzo M, Cosenza L, Granata V, Di Scala C, Nocerino R, Trinchese G, Montella M, et al. Gut Microbiota as a Target for Preventive and Therapeutic Intervention against Food Allergy. Nutrients. 2017; 9(7):672. https://doi.org/10.3390/nu9070672
Chicago/Turabian StyleAitoro, Rosita, Lorella Paparo, Antonio Amoroso, Margherita Di Costanzo, Linda Cosenza, Viviana Granata, Carmen Di Scala, Rita Nocerino, Giovanna Trinchese, Mariangela Montella, and et al. 2017. "Gut Microbiota as a Target for Preventive and Therapeutic Intervention against Food Allergy" Nutrients 9, no. 7: 672. https://doi.org/10.3390/nu9070672
APA StyleAitoro, R., Paparo, L., Amoroso, A., Di Costanzo, M., Cosenza, L., Granata, V., Di Scala, C., Nocerino, R., Trinchese, G., Montella, M., Ercolini, D., & Berni Canani, R. (2017). Gut Microbiota as a Target for Preventive and Therapeutic Intervention against Food Allergy. Nutrients, 9(7), 672. https://doi.org/10.3390/nu9070672