Sugar Metabolism in Hummingbirds and Nectar Bats


Abstract
:1. Introduction
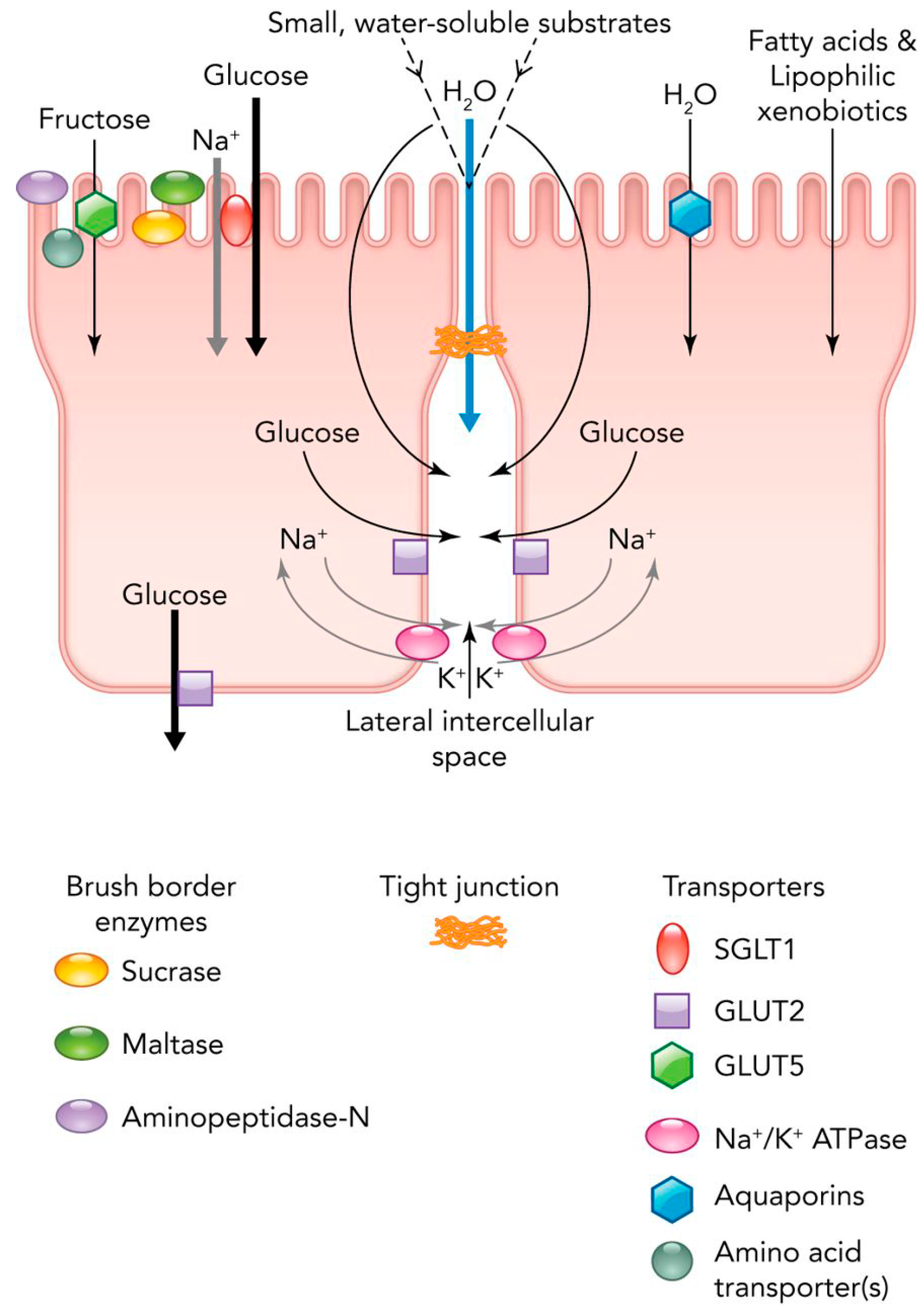
2. Diet and Digestion
3. Biochemical Capacities for Substrate Oxidation
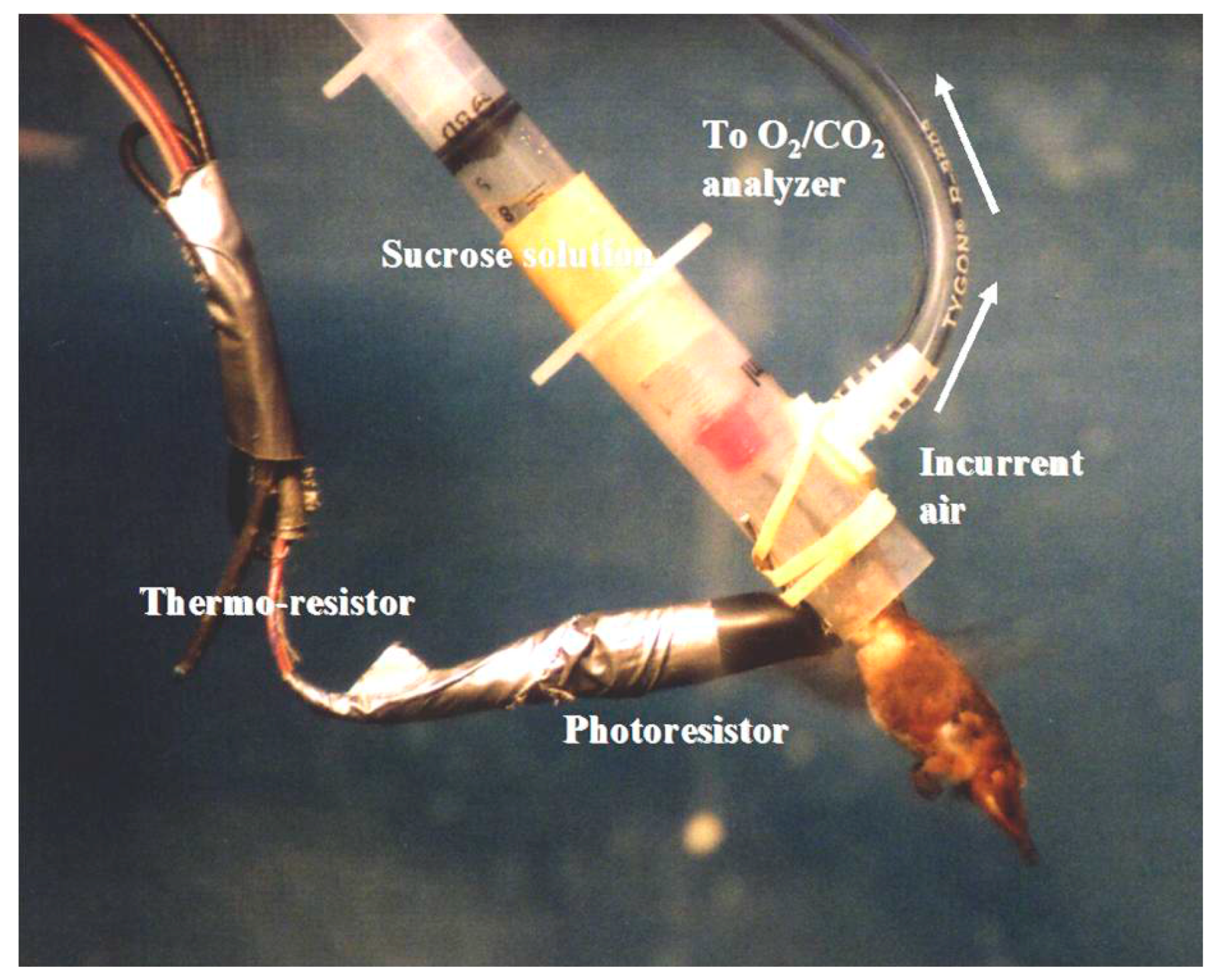
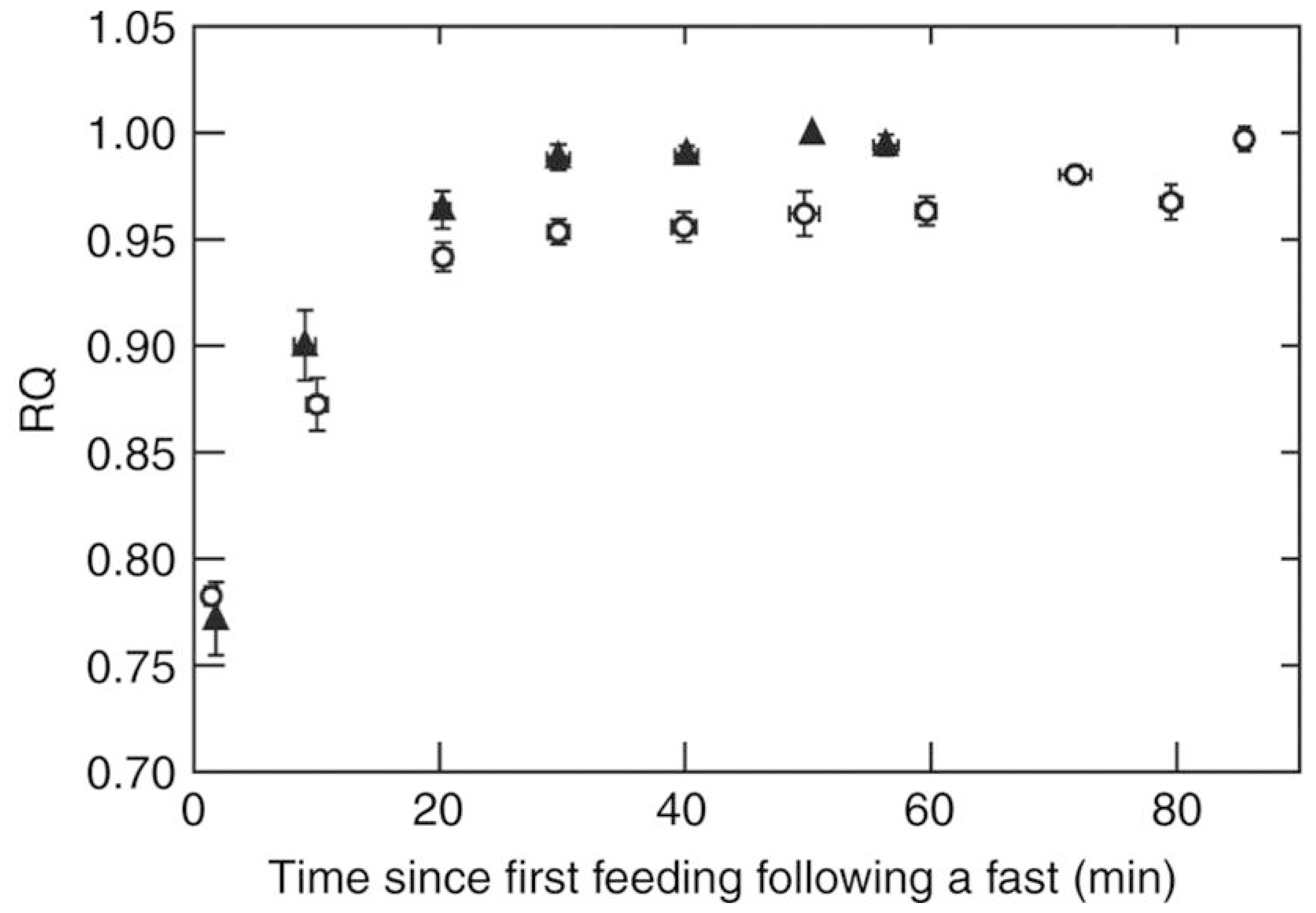
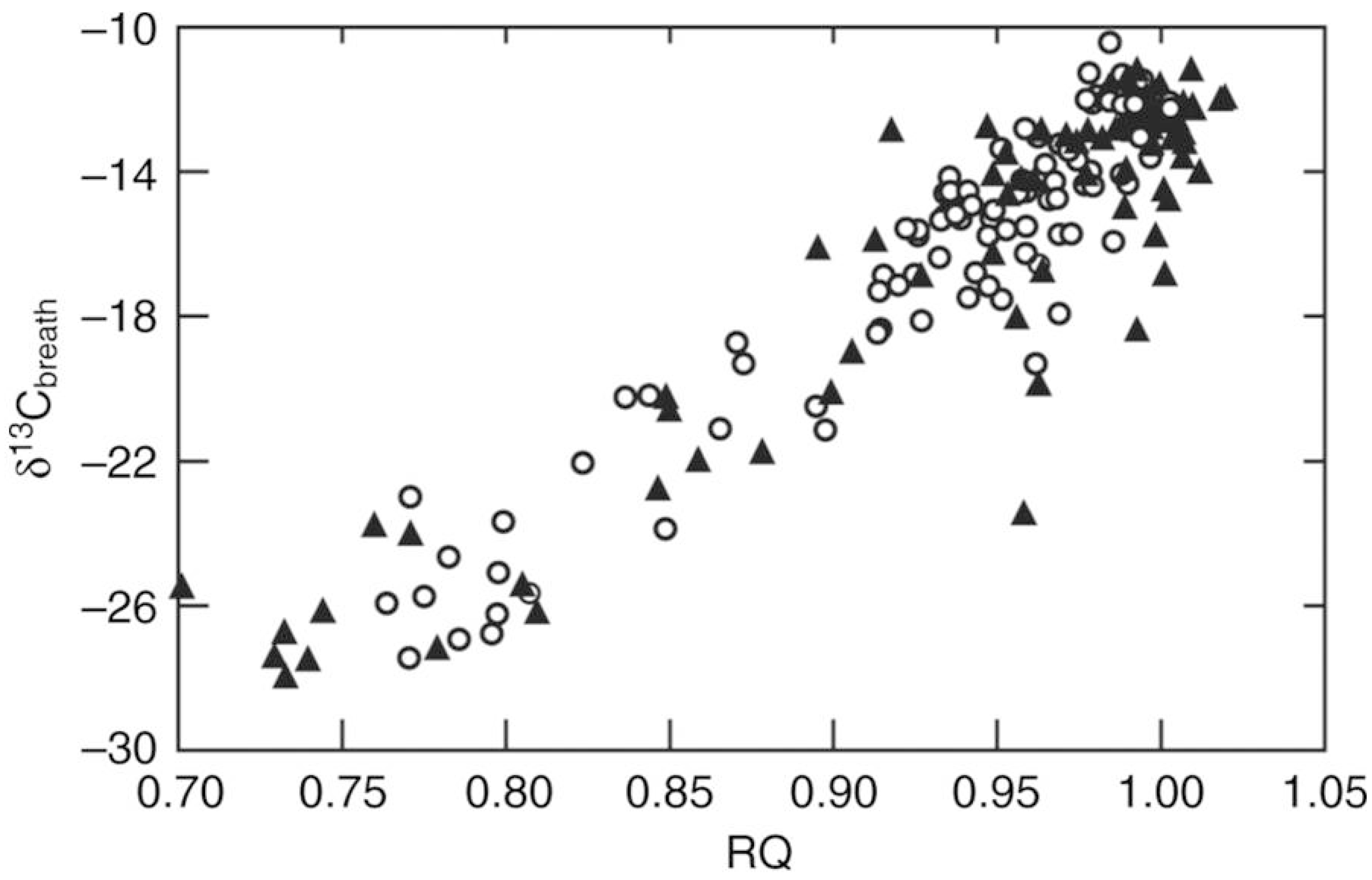
4. Substrate Oxidation during Foraging Flights
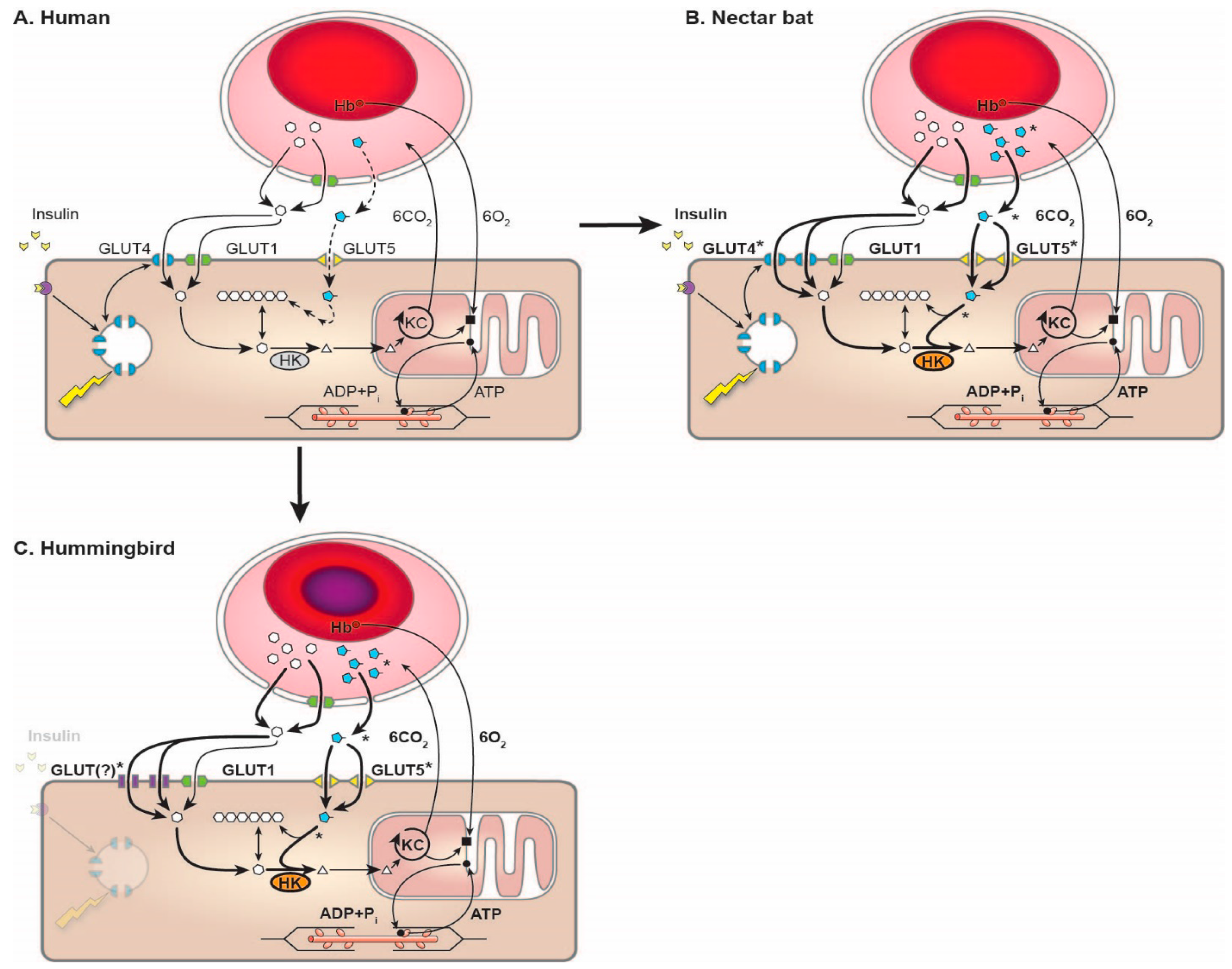
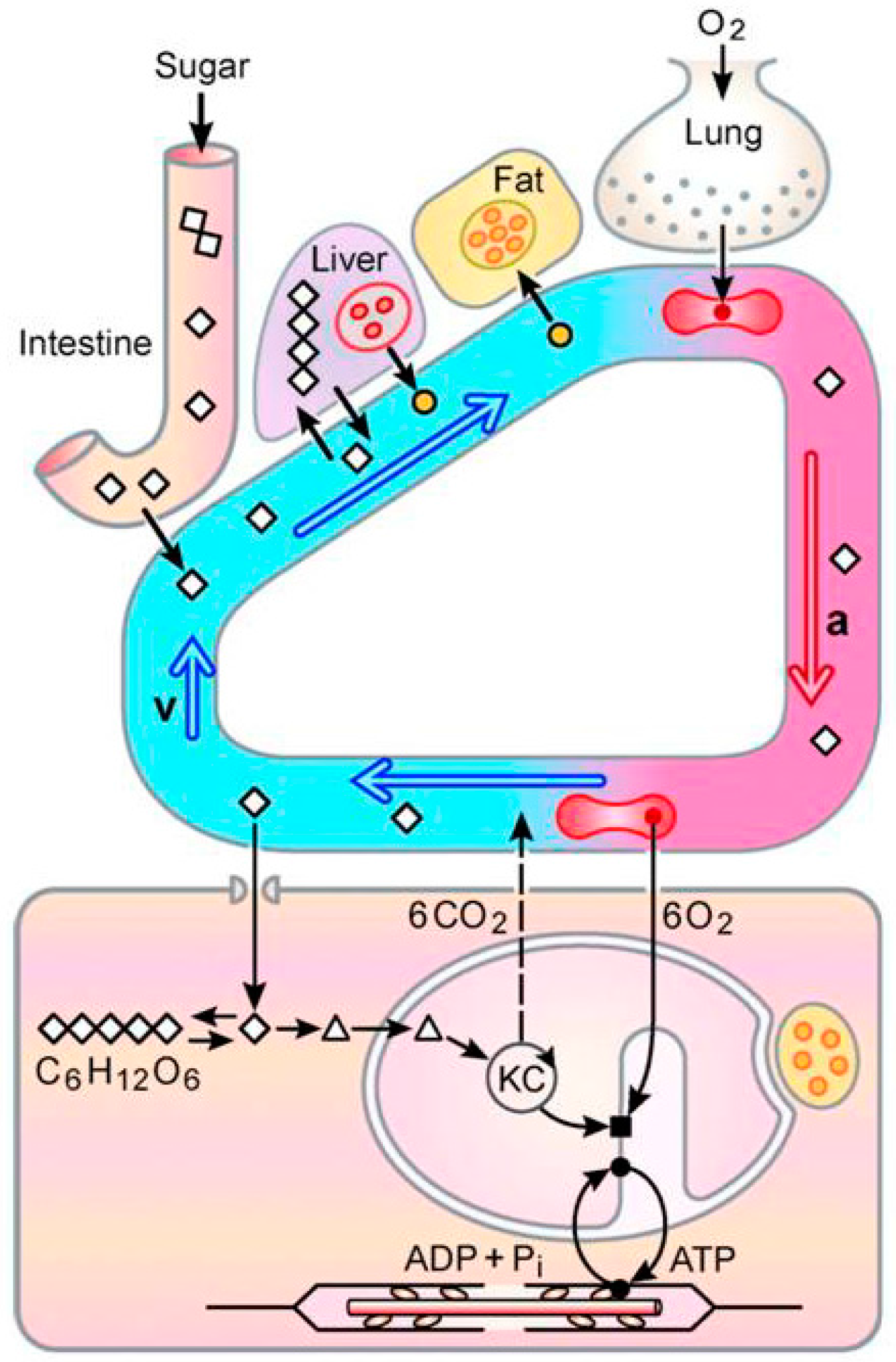
5. A New Concept: The “Sugar Oxidation Cascade”
6. Premigratory Sugar Conversion to Fat in Hummingbirds
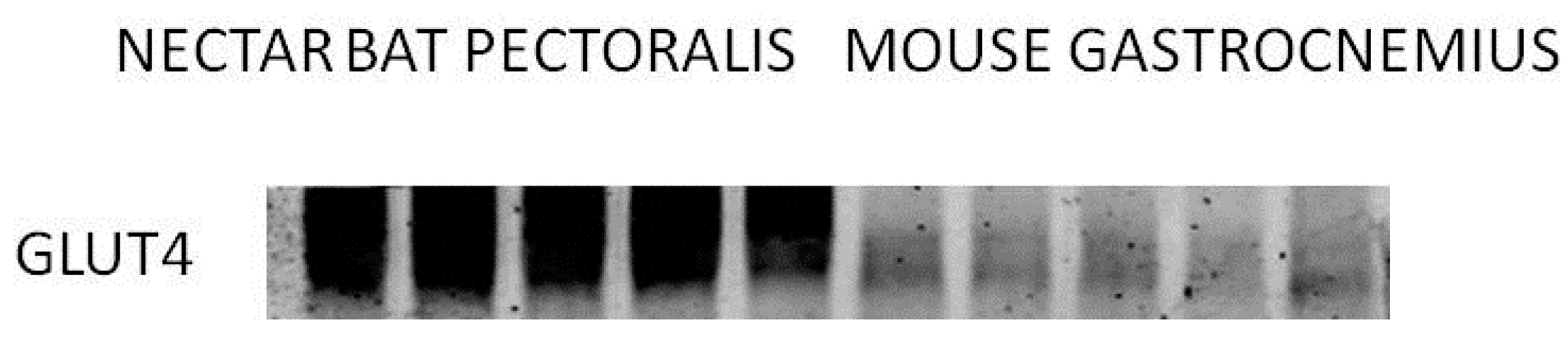
7. Metabolism in Nectarivorous Animals: Implications for Human Health
Acknowledgments
Conflicts of Interest
References
- Heithaus, E.R. Coevolution between bats and plants. In Ecology of Bats; Kunz, T.H., Ed.; Springer: Boston, MA, USA, 1982; pp. 327–367. [Google Scholar]
- Suarez, R.K. Hummingbird flight: Sustaining the highest mass-specific metabolic rates among vertebrates. Experientia 1992, 48, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Suarez, R.K. The sugar oxidation cascade: Aerial refueling in hummingbirds and nectar bats. J. Exp. Biol. 2011, 214, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Von Helversen, O.; Winter, Y. Glossophagine bats and their flowers: Costs and benefits for plants and pollinators. In Bat Ecology; Kunz, T.H., Fenton, B., Eds.; University of Chicago: Chicago, IL, USA, 2003; pp. 346–397. [Google Scholar]
- Powers, D.R.; Nagy, K.A. Field metabolic rate and food consumption by free-living Anna’s hummingbirds (Calypte anna). Physiol. Zool. 1988, 61, 500–506. [Google Scholar] [CrossRef]
- Winter, Y.; Voigt, C.; von Helversen, O. Gas exchange during hovering flight in a nectar-feeding bat Glossophaga soricina. J. Exp. Biol. 1998, 201, 237–244. [Google Scholar] [PubMed]
- Heinrich, B. Energetics of pollination. Ann. Rev. Ecol. Syst. 1975, 6, 139–170. [Google Scholar] [CrossRef]
- Martinez del Rio, C.; Baker, H.G.; Baker, I. Ecological and evolutionary implications of digestive processes: Bird preferences and the sugar constituents of floral nectar and fruit pulp. Experientia 1992, 48, 544–551. [Google Scholar] [CrossRef]
- Nicolson, S.W.; Fleming, P.A. Nectar as food for birds: The physiological consequences of drinking dilute sugar solutions. Plant Syst. Evol. 2003, 238, 139–153. [Google Scholar] [CrossRef]
- Datzmann, T.; von Helversen, O.; Mayer, F. Evolution of nectarivory in phyllostomid bats (Phyllostomidae Gray, 1825, Chiroptera: Mammalia). BMC Evol. Biol. 2010, 10, 165. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.G.; Baker, I.; Hodges, S.A. Sugar composition of nectars and fruits consumed by birds and bats in the tropics and subtropics. Biotropica 1998, 30, 559–586. [Google Scholar] [CrossRef]
- Kelm, D.H.; Schaer, J.; Ortmann, S.; Wibbelt, G.; Speakman, J.R.; Voigt, C.C. Efficiency of facultative frugivory in the nectar-feeding bat Glossophaga commissarisi: The quality of fruits as an alternative food source. J. Comp. Physiol. 2008, 178, 985–996. [Google Scholar] [CrossRef] [PubMed]
- Martinez del Rio, C. Dietary, phylogenetic, and ecological correlates of intestinal sucrase and maltase activity in birds. Physiol. Zool. 1990, 63, 987–1011. [Google Scholar] [CrossRef]
- Diamond, J.M.; Karasov, W.H.; Phan, D.; Carpenter, F.L. Digestive physiology is a determinant of foraging bout frequency in hummingbirds. Nature 1986, 320, 62–63. [Google Scholar] [CrossRef] [PubMed]
- Karasov, W.H.; Phan, D.; Diamond, J.M.; Carpenter, F.L. Food passage and intestinal nutrient absorption in hummingbirds. Auk 1986, 103, 453–464. [Google Scholar]
- McWhorter, T.J.; Bakken, B.H.; Karasov, W.H.; Martinez del Rio, C. Hummingbirds rely on both paracellular and carrier-mediated intestinal glucose absorption to fuel high metabolism. Biol. Lett. 2006, 2, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.; Martinez del Rio, C. Intestinal disaccharidases in five species of phyllostomid bats. Comp. Biochem. Physiol. 1992, 103, 105–111. [Google Scholar]
- Rodriguez-Pena, N.; Price, E.R.; Caviedes-Vidal, E.; Flores-Ortiz, C.M.; Karasov, W.H. Intestinal paracellular absorption is necessary to support the sugar oxidation cascade in nectarivorous bats. J. Exp. Biol. 2016, 219, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Price, E.R.; Brun, A.; Caviedes-Vidal, E.; Karasov, W.H. Digestive adaptations of aerial lifestyles. Physiology 2015, 30, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.C. The power of feeder-mask respirometry as a method for examining hummingbird energetics. Comp. Biochem. Physiol. A 2011, 158, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Altshuler, D.L.; Dudley, R. Kinematics of hovering hummingbird flight along simulated and natural elevational gradients. J. Exp. Biol. 2003, 206, 3139–3147. [Google Scholar] [CrossRef] [PubMed]
- Mahalingan, S.; Welch, K.C., Jr. Neuromuscular control of hovering wingbeat kinematics in response to distinct flight challenges in the ruby-throated hummingbird, Archilochus colubris. J. Exp. Biol. 2013, 216, 4161–4171. [Google Scholar] [CrossRef] [PubMed]
- Norberg, U.M.L.; Winter, Y. Wing beat kinematics of a nectar-feeding bat, Glossophaga soricina, flying at different flight speeds and strouhal numbers. J. Exp. Biol. 2006, 209, 3887–3897. [Google Scholar] [CrossRef] [PubMed]
- Voigt, C.C.; Winter, Y. Energetic cost of hovering flight in nectar-feeding bats (Phyllostomidae: Glossophaginae) and its scaling in moths, birds and bats. J. Comp. Physiol. 1999, 169, 38–48. [Google Scholar] [CrossRef]
- Fons, R.; Sicart, R. Contribution a la connaissance du metabolisme energetique chez deux Crocidurinae: Suncus etruscus (savi, 1822) et Crocidura russula (Hermann, 1780) (insectivora, Soricidae). Mammalia 1976, 40, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Bartholomew, G.A.; Lighton, J.R.B. Oxygen consumption during hover-feeding in free-ranging Anna hummingbirds. J. Exp. Biol. 1986, 123, 191–199. [Google Scholar] [PubMed]
- Taylor, C.R. Structural and functional limits to oxidative metabolism: Insights from scaling. Ann. Rev. Physiol. 1987, 49, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Blem, C.R. Patterns of lipid storage and utilization in birds. Am. Zool. 1976, 16, 671–684. [Google Scholar] [CrossRef]
- Grinyer, I.; George, J.C. Some observations on the ultrastructure of the hummingbird pectoral muscles. Can. J. Zool. 1969, 47, 771–774. [Google Scholar] [CrossRef] [PubMed]
- Suarez, R.K.; Lighton, J.R.B.; Brown, G.S.; Mathieu-Costello, O. Mitochondrial respiration in hummingbird flight muscles. Proc. Natl. Acad. Sci. USA 1991, 88, 4870–4873. [Google Scholar] [CrossRef] [PubMed]
- Hermanson, J.W.; Ryan, J.M.; Cobb, M.A.; Bentley, J.; Schutt, W.A. Histochemical and electrophoretic analysis of the primary flight muscle of several Phyllostomid bats. Can. J. Zool. 1998, 76, 1983–1992. [Google Scholar] [CrossRef]
- Suarez, R.K.; Welch, K.C., Jr.; Hanna, S.K.; Herrera, M.L.G. Flight muscle enzymes and metabolic flux rates during hovering flight of the nectar bat, Glossophaga soricina: Further evidence of convergence with hummingbirds. Comp. Biochem. Physiol. 2009, 153, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Dubach, M. Quantitative analysis of the respiratory system of the house sparrow, budgerigar and violet-eared hummingbird. Respir. Physiol. 1981, 46, 43–60. [Google Scholar] [CrossRef]
- Maina, J.N. What it takes to fly: The structural and functional respiratory requirements in birds and bats. J. Exp. Biol. 2000, 203, 3045–3064. [Google Scholar] [PubMed]
- Schmidt-Nielsen, K. Scaling. Why Is Animal Size So Important? Cambridge University Press: Cambridge, UK, 1984; 241p. [Google Scholar]
- Canals, M.; Atala, C.; Rossi, B.G.; Iriarte-Diaz, J. Relative size of hearts and lungs of small bats. Acta Chiropterol. 2005, 7, 65–72. [Google Scholar] [CrossRef]
- Mathieu-Costello, O.; Suarez, R.K.; Hochachka, P.W. Capillary-to-fiber geometry and mitochondrial density in hummingbird flight muscle. Respir. Physiol. 1992, 89, 113–132. [Google Scholar] [CrossRef]
- Newsholme, E.A.; Crabtree, B. Maximum catalytic activity of some key enzymes in provision of physiologically useful information about metabolic fluxes. J. Exp. Zool. 1986, 239, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Suarez, R.K. Upper limits to mass-specific metabolic rates. Annu. Rev. Physiol. 1996, 58, 583–605. [Google Scholar] [CrossRef] [PubMed]
- Suarez, R.K.; Staples, J.F.; Lighton, J.R.B.; West, T.G. Relationships between enzymatic flux capacities and metabolic flux rates in muscles: Nonequilibrium reactions in muscle glycolysis. Proc. Natl. Acad. Sci. USA 1997, 94, 7065–7069. [Google Scholar] [CrossRef] [PubMed]
- Moyes, C.D. Controlling muscle mitochondrial content. J. Exp. Biol. 2003, 206, 4385–4391. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.M.; Roberts, T.J.; Vock, R.; Weibel, E.R.; Taylor, C.R. Design of the oxygen and substrate pathways. III. Partitioning energy provision from carbohydrates. J. Exp. Biol. 1996, 199, 1659–1666. [Google Scholar] [PubMed]
- Crabtree, B.; Newsholme, E.A. The activities of phosphorylase, hexokinase, phosphofructokinase, lactate dehydrogenase and the glycerol 3-phosphate dehydrogenases in muscles from vertebrates and invertebrates. Biochem. J. 1972, 126, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Wasserman, D.H.; Kang, L.; Ayala, J.E.; Fueger, P.T.; Lee-Young, R.S. The physiological regulation of glucose flux into muscle in vivo. J. Exp. Biol. 2011, 214, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Fell, D.A. Signal transduction and the control of expression of enzyme activity. Adv. Enzyme Regul. 2000, 40, 35–46. [Google Scholar] [CrossRef]
- Brooks, G.A. Mammalian fuel utilization during sustained exercise. Comp. Biochem. Physiol. 1998, 120, 89–107. [Google Scholar] [CrossRef]
- Weber, J.-M.; Haman, F. Oxidative fuel selection: Adjusting mix and flux to stay alive. Int. Congr. Ser. 2004, 1275, 22–31. [Google Scholar] [CrossRef]
- Johansen, K.; Berger, M.; Bicudo, J.E.P.W.; Ruschi, A.; De Almeida, P.J. Respiratory properties of blood and myoglobin in hummingbirds. Physiol. Zool. 1987, 60, 269–278. [Google Scholar] [CrossRef]
- Suarez, R.K.; Brown, G.S.; Hochachka, P.W. Metabolic sources of energy for hummingbird flight. Am. J. Physiol. 1986, 251, R537–R542. [Google Scholar] [PubMed]
- Brand, M.D. The efficiency and plasticity of mitochondrial energy transduction. Biochem. Soc. Trans. 2005, 33, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.C.; Altschuler, D.L.; Suarez, R.K. Oxygen consumption rates in hovering hummingbirds reflect substrate-dependent differences in P/O ratios: Carbohydrate as a ‘premium fuel’. J. Exp. Biol. 2007, 210, 2146–2153. [Google Scholar] [CrossRef] [PubMed]
- Suarez, R.K.; Lighton, J.R.B.; Moyes, C.D.; Brown, G.S.; Gass, C.L.; Hochachka, P.W. Fuel selection in rufous hummingbirds: Ecological implications of metabolic biochemistry. Proc. Natl. Acad. Sci. USA 1990, 87, 9207–9210. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.C., Jr.; Herrera, M.L.G.; Suarez, R.K. Dietary sugar as a direct fuel for flight in the nectarivorous bat, Glossophaga soricina. J. Exp. Biol. 2008, 211, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.C.; Bakken, B.H.; Martinez del Rio, C.; Suarez, R.K. Hummingbirds fuel hovering flight with newly-ingested sugar. Physiol. Biochem. Zool. 2006, 79, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- McNevin, D.B.; Badger, M.R.; Whitney, S.M.; von Caemmerer, S.; Tcherkez, G.B.; Farquhar, G.D. Differences in carbon isotope discrimination of three variants of d-ribulose-1,5-bisphosphate carboxylase/oxygenase reflect differences in their catalytic mechanisms. J. Biol. Chem. 2007, 282, 36068–36076. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.C., Jr.; Peronnet, F.; Hatch, K.A.; Voigt, C.C.; McCue, M.D. Carbon stable-isotope tracking in breath for comparative studies of fuel use. Ann. N. Y. Acad. Sci. 2016, 1365, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Jentjens, R.L.P.G.; Venables, M.C.; Jeukendrup, A.E. Oxidation of exogenous glucose, sucrose, and maltose during prolonged cycling exercise. J. Appl. Physiol. 2004, 96, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Voigt, C.C.; Speakman, J.R. Nectar-feeding bats fuel their high metabolism directly with exogenous carbohydrates. Funct. Ecol. 2007, 21, 913–921. [Google Scholar] [CrossRef]
- Huang, S.; Czech, M.P. The GLUT4 glucose transporter. Cell Metab. 2007, 5, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Seki, Y.; Sato, K.; Kono, T.; Abe, H.; Akiba, Y. Broiler chickens (Ross strain) lack insulin-responsive glucose transporter GLUT4 and have GLUT8 cDNA. Gen. Comp. Endocrinol. 2003, 133, 80–87. [Google Scholar] [CrossRef]
- Sweazea, K.L.; Braun, E.J. Glucose transporter expression in English sparrows (Passer domesticus). Comp. Biochem. Physiol. B 2006, 144, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.C., Jr.; Allalou, A.; Sehgal, P.; Cheng, J.; Ashok, A. Glucose transporter expression in an avian nectarivore: The ruby-throated hummingbird (Archilochus colubris). PLoS ONE 2013, 8, e7703. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.C., Jr.; Chen, C.C.W. Sugar flux through the flight muscles of hovering vertebrate nectarivores: A review. J. Comp. Physiol. 2014, 184, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.W.; Welch, K.C., Jr. Hummingbirds can fuel expensive hovering flight completely with either exogenous glucose or fructose. Funct. Ecol. 2014, 28, 589–600. [Google Scholar] [CrossRef]
- Kristiansen, S.; Darakhshan, F.; Richter, E.A.; Handal, H.S. Fructose transport and GLUT5 protein in human sarcolemmal vesicles. Am. J. Physiol. 1997, 273, E543–E548. [Google Scholar] [PubMed]
- Zierath, J.R.; Nolte, L.A.; Wahlstrom, E.; Galuska, D.; Shepherd, P.R.; Kahn, B.B.; Wallberg-Henriksson, H. Carrier-mediated fructose uptake significantly contributes to carbohydrate metabolism in human skeletal muscle. Biochem. J. 1995, 311, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Jentjens, R.L.P.G.; Achten, J.; Jeukendrup, A.E. High oxidation rates from combined carbohydrates ingested during exercise. Med. Sci. Sports Exerc. 2004, 36, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Beuchat, C.A.; Chong, C.R. Hyperglycemia in hummingbirds and its consequences for hemoglobin glycation. Comp. Biochem. Physiol. 1998, 120, 409–416. [Google Scholar] [CrossRef]
- Kelm, D.H.; Simon, R.; Kuhlow, D.; Voigt, C.C.; Ristow, M. High activity enables life on a high-sugar diet: Blood glucose regulation in nectar-feeding bats. Proc. R. Soc. 2011, 278, 3490–3496. [Google Scholar] [CrossRef] [PubMed]
- Weibel, E.R. The Pathway for Oxygen; Harvard University Press: Cambridge, MA, USA, 1984. [Google Scholar]
- Weibel, E.R.; Taylor, C.R.; Gehr, P.; Hoppeler, H.; Mathieu, O.; Maloiy, G.M.O. Design of the mammalian respiratory system. IX. Functional and structural limits for oxygen flow. Respir. Physiol. 1981, 44, 151–164. [Google Scholar] [CrossRef]
- Goodwin, G.W.; Arteaga, J.R.; Taegtmeyer, H. Glycogen turnover in the isolated working rat heart. J. Biol. Chem. 1995, 270, 9234–9240. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, G.W.; Taylor, C.S.; Taegtmeyer, H. Regulation of energy metabolism of the heart during acute increase in heart work. J. Biol. Chem. 1998, 273, 29530–29539. [Google Scholar] [CrossRef] [PubMed]
- Altshuler, D.L.; Dudley, R.; McGuire, J.A. Resolution of a paradox: Hummingbird flight at high elevation does not come without a cost. Proc. Natl. Acad. Sci. USA 2004, 101, 17731–17736. [Google Scholar] [CrossRef] [PubMed]
- Suarez, R.K.; Gass, C.L. Hummingbird foraging and the relation between bioenergetics and behaviour. Comp. Biochem. Physiol. 2002, 133, 335–343. [Google Scholar] [CrossRef]
- Gass, C.L.; Sutherland, G.D. Specialization by territorial hummingbirds on experimentally enriched patches of flowers: Energetic profitability and learning. Can. J. Zool. 1985, 63, 2125–2133. [Google Scholar] [CrossRef]
- Hou, L.; Welch, K.C., Jr. Premigratory ruby-throated hummingbirds, Archilochus colubris, exhibit multiple strategies for fueling migration. Anim. Behav. 2016, 121, 87–99. [Google Scholar] [CrossRef]
- Lasiewski, R.C. The energetics of migrating hummingbirds. Condor 1962, 64, 324. [Google Scholar] [CrossRef]
- Calder, W. Southbound through Colorado: Migration of rufous hummingbirds. Nat. Geogr. Res. 1987, 3, 40–51. [Google Scholar]
- Welch, K.C.; Suarez, R.K. Altitude and temperature effects on the energetic cost of hover-feeding in migratory rufous hummingbirds, Selasphorus Rufus. Can. J. Zool. 2008, 86, 161–169. [Google Scholar] [CrossRef]
- López-Calleja, M.V.; Bozinovic, F. Maximum metabolic rate, thermal insulation and aerobic scope in a small-sized Chilean hummingbird (Sephanoides sephanoides). Auk 1995, 112, 1034–1036. [Google Scholar]
- Carpenter, F.L.; Hixon, M.A.; Beuchat, C.A.; Russell, R.W.; Paton, D.C. Biphasic mass gain in migrant hummingbirds: Body composition changes, torpor, and ecological significance. Ecology 1993, 74, 1173–1182. [Google Scholar] [CrossRef]
- Gass, C.L.; Romich, M.T.; Suarez, R.K. Energetics of hummingbird foraging at low ambient temperature. Can. J. Zool. 1999, 77, 314–320. [Google Scholar] [CrossRef]
- Suarez, R.K. Oxygen and the upper limits to animal design and performance. J. Exp. Biol. 1998, 201, 1065–1072. [Google Scholar] [PubMed]
- Lane, J.S.; Whang, E.E.; Rigberg, D.A.; Hines, O.J.; Kwan, D.; Zinner, M.J.; McFadden, D.W.; Diamond, J.M.; Ashley, S.W. Paracellular glucose transport plays a minor role in the unanesthetized dog. Am. J. Physiol. 1999, 276, G789–G794. [Google Scholar] [PubMed]
- Basu, S.; Yoffe, P.; Hills, N.; Lustig, R.H. The relationship of sugar to population-level diabetes prevalence: An econometric analysis of repeated cross-sectional data. PLoS ONE 2013, 8, e57873. [Google Scholar] [CrossRef] [PubMed]
- Tappy, L.; Mittendorfer, B. Fructose toxicity: Is the science ready for public heath actions? Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Lutmerding, J.A. Longevity Records of North American Birds; USGS: Reston, VA, USA, 2016.
- Calder, W.A. Avian longevity and aging. In Genetic Effects on Aging II; Harrison, D.E., Ed.; Telford Press: West Caldwell, NJ, USA, 1990; pp. 185–204. [Google Scholar]
- Pedersen, B.K.; Fischer, C.P. Beneficial health effects of exercise—The role of IL-6 as a myokine. Trends Pharmacol. Sci. 2007, 28, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Bishop-Bailey, D. Mechanisms governing the health and performance benefits of exercise. Br. J. Pharmacol. 2013, 170, 1153–1166. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K. Muscles and their myokines. J. Exp. Biol. 2011, 214, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Febbraio, M.A. Muscles, exercise and obesity: Skeletal muscle as a secretory organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Crittenden, A.N. The importance of honey consumption in human evolution. Food Foodways 2011, 19, 257–273. [Google Scholar] [CrossRef]
- Ball, D.W. The chemical composition of honey. J. Chem. Educ. 2007, 84, 1643–1646. [Google Scholar] [CrossRef]
- Marlowe, F. Male contribution to diet and female reproductive success among foragers. Curr. Anthropol. 2001, 42, 755–760. [Google Scholar] [CrossRef]
- Pontzer, H. Lessons from the Hadza: Poor diets wreck efforts to prevent obesity and diabetes. Diabetes Voice 2012, 57, 26–29. [Google Scholar]
- Pontzer, H.; Raichlen, D.A.; Wood, B.M.; Mabulla, A.Z.P.; Racette, S.B.; Marlowe, F.W. Hunter-gatherer energetics and human obesity. PLoS ONE 2012, 7, e40503. [Google Scholar] [CrossRef] [PubMed]
- Krogh-Madsen, R.; Thyfault, J.P.; Broholm, C.; Mortensen, O.H.; Olsen, R.H.; Mounier, R.; Plomgaard, P.; van Hall, G.; Booth, F.W.; Pedersen, B.K. A 2-wk reduction of ambulatory activity attenuates peripheral insulin sensitivity. J. Appl. Physiol. 2010, 108, 1034–1040. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.H.; Krogh-Madsen, R.; Thomsen, C.; Booth, F.W.; Pedersen, B.K. Metabolic responses to reduced daily steps in healthy nonexercising men. JAMA 2008, 299, 1261–1263. [Google Scholar] [PubMed]
- Tappy, L.; Lê, K.A. Metabolic effects of fructose and the worldwide increase in obesity. Physiol. Rev. 2010, 90, 23–46. [Google Scholar] [CrossRef] [PubMed]
- Bidwell, A.J.; Fairchild, T.J.; Redmond, J.; Wang, L.; Keslacy, L.; Kanaley, J.A. Physical activity offsets the negative effects of a high-fructose diet. Med. Sci. Sports Exerc. 2014, 46, 2091–2098. [Google Scholar] [CrossRef] [PubMed]
- Egli, L.; Lecoultre, V.; Theytaz, F.; Campos, V.; Hodson, L.; Schneiter, P.; Mittendorfer, B.; Patterson, B.W.; Fielding, B.A.; Gerber, P.A.; et al. Exercise prevents fructose-induced hypertriglyceridemia in healthy young subjects. Diabetes 2013, 62, 2259–2265. [Google Scholar] [CrossRef] [PubMed]
- Egli, L.; Lecoultre, V.; Cros, J.; Rosset, R.; Marques, A.-S.; Schneiter, P.; Hodson, L.; Gabert, L.; Laville, M.; Tappy, L. Exercise performed immediately after fructose ingestion enhances fructose oxidation and suppresses fructose storage. Am. J. Clin. Nutr. 2016, 103, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Krebs, H.A. The August Krogh principle: “For many problems there is an animal on which it can be most conveniently studied”. J. Exp. Zool. 1975, 194, 221–226. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Nectar Bat 1 Pectoralis | Hummingbird 2 Pectoralis | Shrew 3 Quadriceps | Rat 4 Soleus |
|---|---|---|---|---|
| Glycogen phosphorylase | 46.0 | 59.0 | n.a. | 10.08 |
| Hexokinase | 15.9 | 18.4 | 1.10 | 2.20 |
| Citrate synthase | 204.7 | 448.4 | 37.0 | 45.1 |
| Carnitine palmitoyl transferase | 6.0 | 7.2 | 2.7 | 0.28 |
| Nectar Bat | Hummingbird | |
|---|---|---|
| Whole-body V (mL O2 g−1 h−1) | 24.5 | 33.3 |
| Flight muscle V (mL O2 g−1 h−1) | 84.7 | 119.8 |
| Glucose oxidation rate (µmol g−1 min−1) | 9.1 | 14.8 |
| Palmitate oxidation rate (µmol g−1 min−1) | 2.0 | 2.8 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suarez, R.K.; Welch, K.C. Sugar Metabolism in Hummingbirds and Nectar Bats. Nutrients 2017, 9, 743. https://doi.org/10.3390/nu9070743
Suarez RK, Welch KC. Sugar Metabolism in Hummingbirds and Nectar Bats. Nutrients. 2017; 9(7):743. https://doi.org/10.3390/nu9070743
Chicago/Turabian StyleSuarez, Raul K., and Kenneth C. Welch. 2017. "Sugar Metabolism in Hummingbirds and Nectar Bats" Nutrients 9, no. 7: 743. https://doi.org/10.3390/nu9070743
APA StyleSuarez, R. K., & Welch, K. C. (2017). Sugar Metabolism in Hummingbirds and Nectar Bats. Nutrients, 9(7), 743. https://doi.org/10.3390/nu9070743