Sechium edule (Jacq.) Swartz, a New Cultivar with Antiproliferative Potential in a Human Cervical Cancer HeLa Cell Line


 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Flores, M.E. The chayote, Sechium edule Swartz (Cucurbitaceae). Rev. Biol. Trop. 1989, 37, 1–54. Available online: https://www.ncbi.nlm.nih.gov/pubmed/2690200 (accessed on 30 March 2017). [PubMed]
- Rzedowski, J. Aspectos de las plantas ornamentales mexicanas. Chapingo Serie Hortic. 1995, 1, 5–7. [Google Scholar]
- Lira-Saade, R. Chayote. Sechium edule (Jacq.) Sw. In Promoting the Conservation and Use of Underutilized and Neglected Crops. 8; Institute of Plant Genetics and Crop Plant Research; Gatersleben/International Plant Genetic Resources Institute: Rome, Italy, 1996; p. 57. Available online: http://pdf.usaid.gov/pdf_docs/Pnach876.pdf (accessed on 29 March 2017).
- Avendaño-Arrazate, C.H.; Cadena-Iñiguez, J.; Arevalo-Galarza, M.L.C.; Campos-Rojas, E.; Cisneros-Solano, V.M.; Aguirre-Medina, J.F. Las Variedades del Chayote Mexicano, Recurso Ancestral con Potencial de Comercialización; Grupo Interdisciplinario de Investigación en Sechium edule en México AC (GISeM): Estado de México, Mexico, 2010; Available online: http://snics.mx/sinarefi/biblioteca/Las%20variedades%20dek%20chayote%20mexicano.%20recurso%20ancestral%20con%20potencial%20de%20comercializacion.pdf (accessed on 29 March 2017).
- Cambar, P.J.; Portillo, P.; Tabora, F.E.; de Pineda, L.; Tovar, O.; Casco, J.; Alvarado, C.; Díaz, G.; Casco, B.; Cantillo, L. Estudio preliminar sobre las acciones farmacológicas de Sechium edule. Rev. Méd. Hondur. 1980, 48, 97–99. Available online: http://www.bvs.hn/RMH/pdf/1980/pdf/Vol48-4-1980-3.pdf (accessed on 29 March 2017).
- Cambar, P.J. Algunos estudios farmacológicos de las plantas medicinales en Honduras. Rev. Méd. Hondur. 1985, 53, 190–196. Available online: http://www.bvs.hn/RMH/pdf/1985/pdf/Vol53-3-1985-3.pdf (accessed on 29 March 2017).
- Ordoñez, A.A.L.; Gomez, J.D.; Cudmani, N.M.; Vattuone, M.A.; Isla, M.I. Antimicrobial activity of nine extracts of Sechium edule (Jacq.) Swartz. Microb. Ecol. Health Dis. 2003, 15, 33–39. [Google Scholar] [CrossRef]
- Ordoñez, A.A.L.; Gomez, J.D.; Vattuone, M.A.; Isla, M.I. Antioxidant activities of Sechium edule (Jacq.) Swartz extracts. Food Chem. 2006, 97, 452–458. [Google Scholar] [CrossRef]
- Salama, A.M.; Polo, A.E.; Contreras, C.R.; Maldonado, L. Análisis fitoquímico preliminar y determinación de las actividades antiinflamatoria y cardíaca de los frutos de Sechium edule. Rev. Colomb. Cienc. Quím. Farm. 1986, 15, 79–82. Available online: http://www.ciencias.unal.edu.co/unciencias/data-file/farmacia/revista/V15P79–82.pdf (accessed on 29 March 2017).
- Salama, A.M.; Achenbach, H.; Sánchez, M.Y.; Gutiérrez, M. Aislamiento e identificación de glicósidos antiinflamatorios de los frutos de Sechium edule. Rev. Colomb. Cienc. Quím. Farm. 1987, 16, 15–16. Available online: http://www.revistas.unal.edu.co/index.php/rccquifa/article/viewFile/56592/55530 (accessed on 29 March 2017).
- Gordon, E.A.; Guppy, L.J.; Nelson, M. The antihypertensive effects of the Jamaican Cho-Cho (Sechium edule). West Indian Med. J. 2000, 49, 27–31. Available online: https://www.ncbi.nlm.nih.gov/pubmed/10786447 (accessed on 29 March 2017). [PubMed]
- Mohammed, F.S.; Ahmed, S.; Dey, S. Antiepileptic and central nervous system depressant activity of Sechium edule fruit extract. Bangladesh J. Pharmacol. 2012, 7, 199–202. [Google Scholar] [CrossRef]
- Cadena-Iñiguez, J.; Avendaño-Arrazate, C.H.; Soto-Hernández, M.; Ruiz-Posadas, L.M.; Aguirre-Medina, J.F.; Arévalo-Galarza, L. Infraspecific variation of Sechium edule in the state of Veracruz, Mexico. Genet. Resour. Crop Evol. 2008, 55, 835–847. [Google Scholar] [CrossRef]
- Cadena-Iñiguez, J.; Soto, H.M.; Arévalo, G.M.L.; Avendaño, A.C.H.; Aguirre, M.J.F.; Ruiz, P.L.M. Caracterización bioquímica de variedades domesticadas de Sechium edule (Jacq.) Sw. comparadas con parientes silvestres. Rev. Chapingo Ser. Hortic. Ed. Espec. 2011, 17, 45–55. Available online: http://www.scielo.org.mx/pdf/rcsh/v17nspe2/v17nspe2a5.pdf (accessed on 29 March 2017).
- Cadena-Iñiguez, J.; Arévalo-Galarza, L.; Avendaño-Arrazate, C.H.; Soto-Hernández, M.; Ruiz-Posadas, L.M.; Santiago-Osorio, E.; Acosta-Ramos, M.; Cisneros-Solano, V.M.; Aguirre-Medina, J.F.; Ochoa-Martínez, D. Production, genetics, postharvest management and pharmacological characteristics of Sechium edule (Jacq.) Sw. Fresh Prod. Gl. Sci. Books 2007, 1, 41–53. Available online: http://www.globalsciencebooks.info/Online/GSBOnline/images/0706/FP_1(1)/FP_1(1)41-53o.pdf (accessed on 29 March 2017).
- Cadena-Iñiguez, J.; Soto-Hernández, M.; Torres-Salas, A.; Aguiñiga-Sánchez, I.; Ruiz-Posadas, L.; Rivera-Martínez, A.R.; Avendaño-Arrazate, C.; Santiago-Osorio, E. The antiproliferative effect of chayote varieties (Sechium edule (Jacq.) Sw.) on tumour cell lines. J. Med. Plants Res. 2013, 7, 455–460. [Google Scholar] [CrossRef]
- Monroy-Vázquez, M.E.; Soto-Hernández, M.; Cadena-Iñiguez, J.; Santiago-Osorio, E.; Ruiz-Posadas, L.M.; Rosas-Acevedo, H. Estudio biodirigido de un extracto alcohólico de frutos de Sechium edule (Jacq.) Swartz. Agrociencia 2009, 43, 777–790. Available online: http://www.scielo.org.mx/pdf/agro/v43n8/v43n8a2.pdf (accessed on 29 March 2107).
- Aguiñiga-Sánchez, I. Potencial Antileucémico In Vitro de Extractos de Cuatro Genotipos de Sechium spp. (Cucurbitaceae). Tesis de Maestría, Colegio de Postgraduados: Montecillo Texcoco, Mexico, 2013; p. 109. [Google Scholar]
- Aguiñiga-Sánchez, I.; Soto-Hernández, M.; Cadena-Iñiguez, J.; Ruíz-Posadas, L.M.; Cadena-Zamudio, J.D.; González-Ugarte, A.K.; Weiss-Steiderd, B.; Santiago-Osorio, E. Fruit extract from a Sechium edule hybrid induce apoptosis in leukaemic cell lines but not in normal cells. Nutr. Cancer. 2015, 67, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, T.; Tommasi, N.; de Morelli, I.; Braca, A. Study of Flavonoids of Sechium edule (Jacq) Swartz (Cucurbitaceae) different edible organs by liquid chromatography photodiode array mass spectrometry. J. Agric. Food Chem. 2004, 52, 6510–6515. Available online: http://pubs.acs.org/doi/pdf/10.1021/jf040214q (accessed on 29 March 2017). [CrossRef] [PubMed]
- Palacios-López, K.; Rodríguez-López, N. Plasticidad fenotípica en Lippia alba (Verbenaceae) en respuesta a la disponibilidad hídrica en dos ambientes lumínicos. Acta Biol. Colomb. 2007, 12, 187–198. Available online: http://www.redalyc.org/articulo.oa?id=319027880011 (accessed on 29 March 2017).
- Villamizar, C.J.M.; Rodríguez, L.N.F.; Tezara, F.W. Plasticidad fenotípica en plantas de Lippia dulcis (Verbenaceae) sometidas a déficit hídrico. Acta Biol. Colomb. 2012, 17, 363–378. Available online: http://www.revistas.unal.edu.co/index.php/actabiol/article/view/27841/32947 (accessed on 27 March 2017).
- Molina, G.J.D. Selección masal para resistencia a sequía de maíz. Agrociencia 1980, 42, 69–77. [Google Scholar]
- Avendaño Arrazate, C.H.; Molina Galán, J.D.; Moreno Pérez, E.D.C.; Cadena Iñiguez, J.; Aguirre Medina, J.F.; Rincón Enríquez, G. Respuesta a la selección para resistencia a sequía en maíz (Zea mays, L.). Interciencia 2009, 34, 801–807. Available online: http://www.scielo.org.ve/scielo.php?script=sci_arttext&pid=S0378-18442009001100010 (accessed on 29 March 2017).
- Avendaño-Arrazate, C.M.; Cadena-Iñiguez, J.; Arévalo-Galarza, M.L.C.; Cisneros-Solano, V.M.; Morales-Flores, F.J.; Ruiz-Posadas, L.M. Mejoramiento genético participativo en chayote. Agroproductividad 2014, 7, 30–39. Available online: http://www.colpos.mx/wb_pdf/Agroproductividad/2014/AGROPRODUCTIVIDAD_VI_2014.pdf (accessed on 29 March 2017).
- Che, C.; Fang, X.; Phoebe, C.H.; Kiunghorn, A.D.; Farnswworth, N.R. High-field 1H-NRM spectral analysis of some cucurbitacins. J. Nat. Prod. 1985, 48, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Afifi, M.S.; Ross, S.A.; Sohly, M.A.; Naeem, Z.E.; Halaweish, F.T. Cucurbitacins of Cucumis prophetarum and Cucumis prophetarum. J. Chem. Ecol. 1999, 25, 847–859. [Google Scholar] [CrossRef]
- Cadena-Iñiguez, J. Caracterización Morfoestructural, Fisiológica, Química y Genética de Diferentes Tipos de Chayote [Sechium edule (Jacq.) Sw.]. Ph.D. Thesis, Colegio de Postgraduados, Montecillo, Texcoco, Estado de México, México, 2005; p. 156. [Google Scholar]
- Wagner, H.; Bland, S. Plant Drug Analysis: A Thin Layer Chromatography Atlas, 2nd ed.; Springer: Munuch, Germany, 2001; p. 371. [Google Scholar]
- Gillies, R.; Didier, N.; Dentosn, M. Determination of cell number in monolayer cultures. Anal. Biochem. 1986, 159, 109–113. [Google Scholar] [CrossRef]
- Kueng, W.; Silver, E.; Eppnberg, V. Quantification of cell cultured on 96-wels plates. Anal. Biochem. 1989, 186, 16–19. [Google Scholar] [CrossRef]
- Gershenzon, J.; Croteau, R. Terpenoids. In Hervibores Their Interactions with Secondary Plant Metabolites, 2nd ed.; Rosenthal, G.A.Y., Berenbaum, M.R., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1991; Chapter 5; Volume 1, p. 468. [Google Scholar]
- Blasiak, J.; Kowalik, J.; Malecka-Panas, E.; Drzewoski, J.; Wojewódzka, M. DNA Damage and repair in human lymphocytes exposed to three anticancer platinum drugs. Teratog. Carcinnog. Mutagen. 2000, 20, 119–131. Available online: http://onlinelibrary.wiley.com/doi/10.1002/(SICI)1520-6866(2000)20:3%3C119::AID-TCM3%3E3.0.CO;2-Z/epdf (accessed on 26 April 2017). [CrossRef]
- Xiong, L.Z.; Xu, C.G.; Saghai Maroof, M.A.; Zhang, Q.F. Patterns of cytosine methylation in an elite rice hybrid and its parental lines by a methylation-sensitive amplification polymorphism technique. Mol. Gen. Genet. 1999, 261, 439–446. Available online: http://www.ncpgr.cn/papers/xiong_mgg.pdf (accessed on 29 March 2017). [CrossRef] [PubMed]
- Bossdorf, O.; Richards, C.L.; Pigliucci, M. Epigenetics for ecologists. Ecol. Lett. 2007, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Richards, E.J. Inherited epigenetic variation—Revisiting soft inheritance. Nat. Rev. Genet. 2006, 7, 395–401. Available online: http://www.nature.com/nrg/journal/v7/n5/pdf/nrg1834.pdf (accessed on 29 March 2017). [CrossRef] [PubMed]
- Guangyuan, L.; Xiaoming, W.; Biyun, C.; Gao, G.; Kun, X. Evaluation of Genetic and Epigenetic Modification in Rapeseed (Brassica napus) Induced by Salt Stress. J. Integr. Plant Biol. 2007, 49, 1599–1607. [Google Scholar] [CrossRef]
- Jirtle, R.L.; Skinner, M.K. Environmental epigenomics and disease susceptibility. Nat. Rev. Genet. 2007, 8, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Masuelli, R.W.; Marfil, C.F. Variabilidad epigenética en plantas y evolución. J. Basic Appl. Genet. 2011, 22, 1–8. Available online: http://www.scielo.org.ar/pdf/bag/v22n1/v22n1a01.pdf (accessed on 29 March 2017).
- Brahmkshatriya, P.P.; Brahmkshatriya, P.S. Terpenes: Chemistry, biological role, and therapeutic applications. In Natural Products; Ramawat, K.G., Mérillon, J.M., Eds.; Springer: Berlin, Germany, 2013; Chapter 86; pp. 2665–2691. Available online: https://link.springer.com/referenceworkentry/10.1007%2F978-3-642-22144-6_120 (accessed on 29 March 2017).
- Takahashi, N.; Yoshida, Y.; Sugiura, T.; Matsuno, K.; Fujino, A.; Yamashita, U. Cucurbitacin D isolated from Trichosanthes kirilowii induces apoptosis in human hepatocellular carcinoma cells in vitro. Int. Immunopharmacol. 2009, 9, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Kira, N.; Yoshida, T.; Narahara, H. Cucurbitacin D induces growth inhibition, cell cycle arrest, and apoptosis in human endometrial and ovarian cancer cells. Tumor Biol. 2013, 34, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Chao-Chen, J.; Hua-Chiu, M.; Lin-Nie, R.; Cordell, A.G.; Qiu, S.X. Cucurbitacins and cucurbitane glycosides: Structures and biological activities. Nat. Prod. Rep. 2005, 22, 386–399. Available online: http://pubs.rsc.org/en/content/articlepdf/2005/np/b418841c (accessed on 29 March 2017). [CrossRef] [PubMed]
- Ding, N.; Yamashita, U.; Matsuoka, H.; Sugiura, T.; Tsukada, J.; Noguchi, J.; Yoshida, Y. Apoptosis induction through proteasome inhibitory activity of Cucurbitacin D in human T-cell leukemia. Cancer 2011, 117, 2735–2746. [Google Scholar] [CrossRef] [PubMed]
- Lui, V.; Yau, D.; Wong, E.; Ng, Y.-K.; Lau, C.; Ho, Y.; Chan, J.; Hong, B.; Ho, K.; Cheung, C.; et al. Cucurbitacin I elicits anoikis sensitization, inhibits cellular invasion and in vivo tumor formation ability of nasopharyngeal carcinoma cells. Carcinogenesis 2009, 30, 2085–2094. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, S. Cucurbitacin B and its rapidly emerging role in the management of systemic malignancies besides lung carcinomas. Cancer Biother. Radiopharm. 2013, 28, 359. [Google Scholar] [CrossRef] [PubMed]
- Soto-Hernández, M.; Cadena-Iñiguez, J.; Arévalo-Galarza, L.; Santiago-Osorio, E.; Aguiñiga-Sánchez, I.; Ruíz-Posadas, L.; del Mar Ruíz-Posadas, L. Lead compounds from Cucurbitaceae for the treatment of cáncer. In Phytochemicals—Isolation, Characterization and Role in Human Health; Rao, A.V., Rao, L.G., Eds.; Intech: Rijeka, Croatia, 2015; Chapter 12; pp. 289–303. [Google Scholar] [CrossRef]
- Alghasham, A.A. Cucurbitacins—A promising target for cancer therapy. Int. J. Health Sci. 2013, 7, 67–79. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3612419/ (accessed on 29 March 2017). [CrossRef]
- Cartaya, O.; Reynaldo, I. Flavonoides: Características químicas y aplicaciones. Cultiv. Trop. 2001, 22, 5–14. Available online: http://www.redalyc.org/pdf/1932/193215009001.pdf (accessed on 29 March 2017).
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.C.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A.M. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. Available online: http://ajcn.nutrition.org/content/74/4/418.full.pdf+html (accessed on 29 March 2017). [PubMed]
- Martínez-Flórez, S.; González-Gallego, J.; Culebras, J.M.; Tuñón, M.J. Los flavonoides: propiedades y acciones antioxidantes. Nutr. Hosp. 2002, 17, 271–278. Available online: https://www.google.com.mx/search?q=Los+flavonoides%3A+propiedades+y+acciones+antioxidantes+Nutr.+Hosp&rlz=1C1AVNE_enMX678MX687&oq=Los+flavonoides%3A+propiedades+y+acciones+antioxidantes+Nutr.+Hosp&aqs=chrome..69i57.3802j0j8&sourceid=chrome&ie=UTF-8 (accessed on 30 March 2017).
- Álvarez, C.E.; Orallo, C.F. Actividad biológica de los flavonides (I). Acción frente al cáncer. Offarm Bioquími. 2003, 22, 130–140. Available online: http://www.elsevier.es/es-revista-offarm-4-articulo-actividad-biologica-los-flavonoides-i-13054406 (accessed on 29 March 2017).
- Kanadaswami, C.; Lee, L.T.; Lee, P.P.H.; Hwang, J.J.; Ke, F.C.; Huang, Y.T.; Lee, M.T. The Antitumor Activities of Flavonoids. In Vivo 2005, 19, 895–910. Available online: http://iv.iiarjournals.org/content/19/5/895.long (accessed on 29 March 2017).
- Agrawal, A.D. Pharmacological activities of flavonoids: A review. Pharma. Sienc. Nanotech. 2011, 4, 1394–1398. Available online: http://www.ijpsnonline.com/Issues/1394_full.pdf (accessed on 29 March 2017).
- Kaur, S.H.; Kumar, B.; Prasher, S.; Tiwari, P.; Salhan, M.; Sharma, P. A Review of phytochemistry and pharmacology of flavonoids. Int. Pharm. Sci. 2011, 1, 25–41. Available online: http://www.academia.edu/8181572/A_Review_of_Phytochemistry_and_Pharmacology_of_Flavonoids_INTRODUCTION (accessed on 29 March 2017).
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed]
- Caltagirone, S.; Rossi, C.; Poggi, A.; Ranelletti, F.O.; Natali, P.G.; Brunetti, M.; Aiello, F.B.; Piantelli, M. Flavonoids apigenin and quercetin inhibit melanoma growth and metastatic potential. Int. J. Cancer 2000, 87, 595–600. Available online: https://www.researchgate.net/publication/227498089_Flavonoids_Apigenin_and_Quercetin_inhibit_melanoma_growth_and_metastatic_potential (accessed on 29 March 2017). [CrossRef]
- Boik, J. Natural Compounds in Cancer Therapy: Promising Nontoxic Antitumor Agents form Plants and Other Natural Sources; Oregon Medical Press: Princeton, MN, USA, 2001. [Google Scholar]







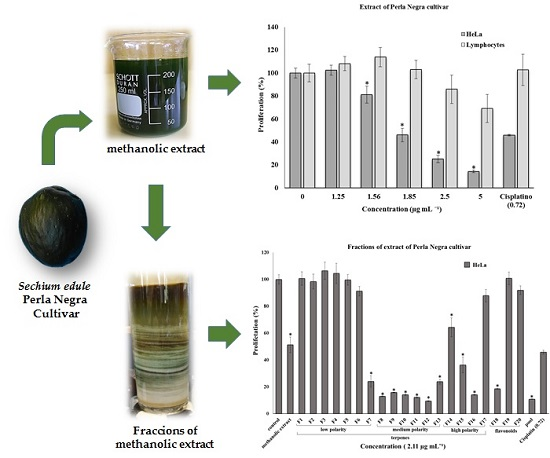{kind=link}
{kind=link}
{kind=link}
{kind=link}
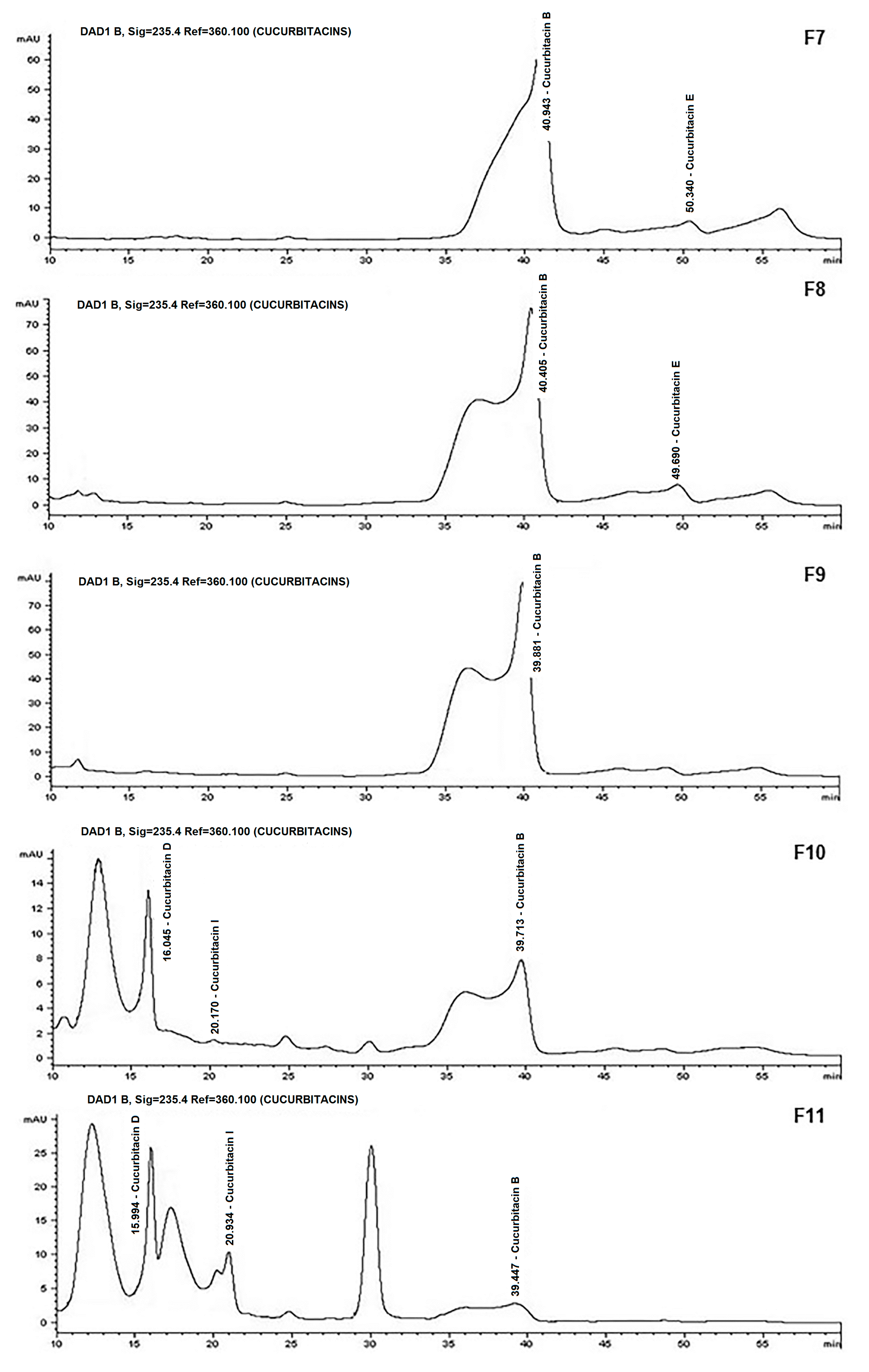{kind=link}
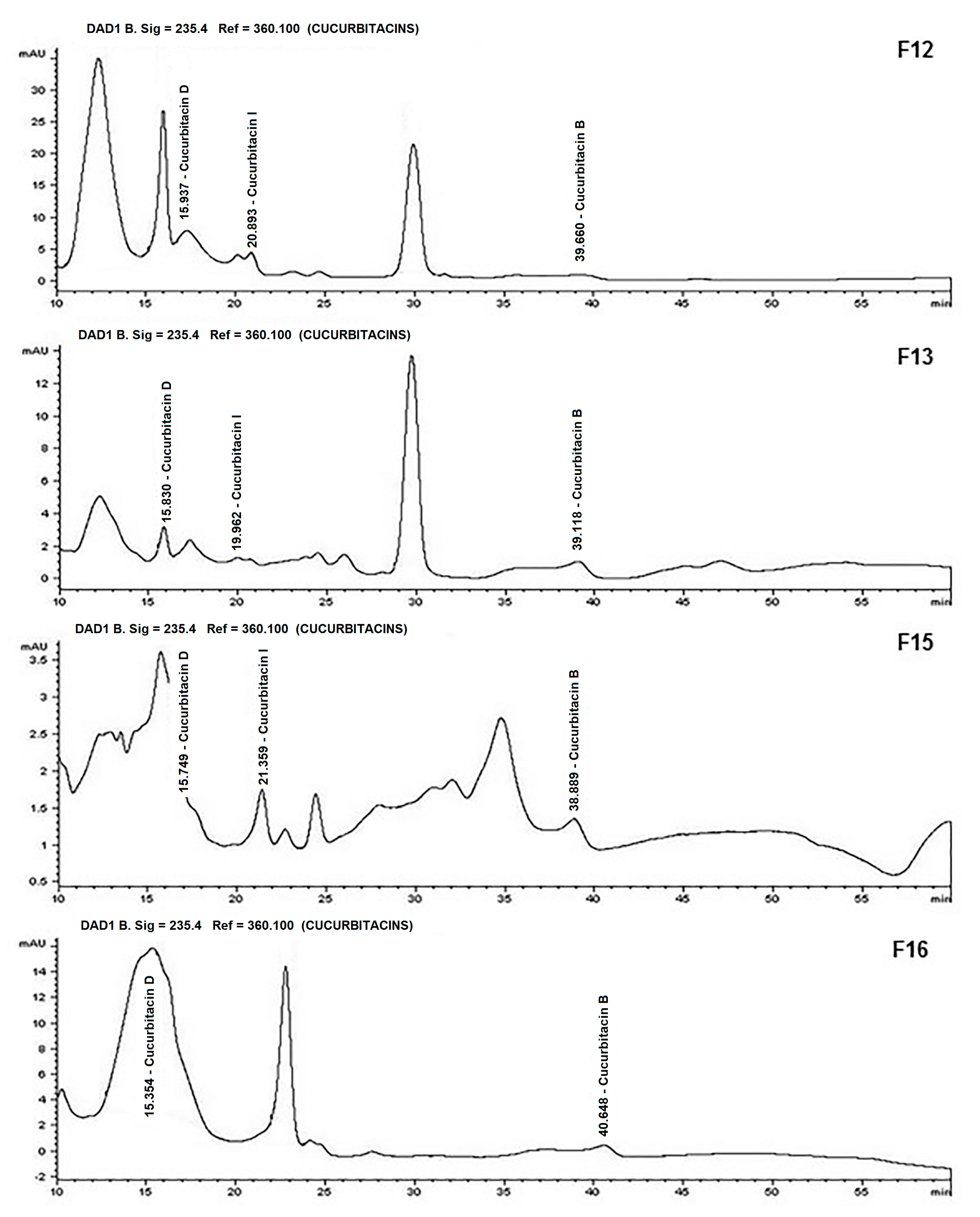{kind=link}
{kind=link}
| FRACTION (mg) | YIELD (%) | Cu B (µg) | Cu D (µg) | Cu E (µg) | Cu I (µg) |
|---|---|---|---|---|---|
| F7 | 1.59 | 29.61 | 0 | 42.39 | 0 |
| F8 | 1.44 | 48.16 | 0 | 53.84 | 0 |
| F9 | 0.74 | 53.12 | 0 | 0 | 0 |
| F10 | 1.34 | 5.52 | 75.09 | 0 | 0 |
| F11 | 1.81 | 0 | 108.60 | 1.53 | 30.40 |
| F12 | 1.49 | 0 | 134.67 | 0.46 | 8.92 |
| F13 | 1.44 | 0.29 | 7.06 | 0 | 0 |
| F15 | 1.28 | 0 | 16.13 | 0 | 1.05 |
| F16 | 3.94 | 0.30 | 594.39 | 0 | 0 |
| TOTAL | 15.07 | 137.00 | 935.94 | 98.21 | 40.37 |
| Fraction (mg) | Yield (%) | Rutin (µg) | Miricetin (µg) | Quercetin (µg) | Naringenin (µg) | Phloretin (µg) | Galangin (µg) | Apigenin (µg) |
|---|---|---|---|---|---|---|---|---|
| F18 | 12.07 | 0.34 | 1.85 | 0.25 | 2.26 | 4.72 | 0.43 | 0.18 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salazar-Aguilar, S.; Ruiz-Posadas, L.D.M.; Cadena-Iñiguez, J.; Soto-Hernández, M.; Santiago-Osorio, E.; Aguiñiga-Sánchez, I.; Rivera-Martínez, A.R.; Aguirre-Medina, J.F. Sechium edule (Jacq.) Swartz, a New Cultivar with Antiproliferative Potential in a Human Cervical Cancer HeLa Cell Line. Nutrients 2017, 9, 798. https://doi.org/10.3390/nu9080798
Salazar-Aguilar S, Ruiz-Posadas LDM, Cadena-Iñiguez J, Soto-Hernández M, Santiago-Osorio E, Aguiñiga-Sánchez I, Rivera-Martínez AR, Aguirre-Medina JF. Sechium edule (Jacq.) Swartz, a New Cultivar with Antiproliferative Potential in a Human Cervical Cancer HeLa Cell Line. Nutrients. 2017; 9(8):798. https://doi.org/10.3390/nu9080798
Chicago/Turabian StyleSalazar-Aguilar, Sandra, Lucero Del Mar Ruiz-Posadas, Jorge Cadena-Iñiguez, Marcos Soto-Hernández, Edelmiro Santiago-Osorio, Itzen Aguiñiga-Sánchez, Ana Rocío Rivera-Martínez, and Juan Francisco Aguirre-Medina. 2017. "Sechium edule (Jacq.) Swartz, a New Cultivar with Antiproliferative Potential in a Human Cervical Cancer HeLa Cell Line" Nutrients 9, no. 8: 798. https://doi.org/10.3390/nu9080798
APA StyleSalazar-Aguilar, S., Ruiz-Posadas, L. D. M., Cadena-Iñiguez, J., Soto-Hernández, M., Santiago-Osorio, E., Aguiñiga-Sánchez, I., Rivera-Martínez, A. R., & Aguirre-Medina, J. F. (2017). Sechium edule (Jacq.) Swartz, a New Cultivar with Antiproliferative Potential in a Human Cervical Cancer HeLa Cell Line. Nutrients, 9(8), 798. https://doi.org/10.3390/nu9080798