Anti-Diabetic Effects of Phenolic Extract from Rambutan Peels (Nephelium lappaceum) in High-Fat Diet and Streptozotocin-Induced Diabetic Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Experimental Animals
2.3. Experimental Design
2.3.1. Modeling Method
2.3.2. Biochemical Assays
2.3.3. Histopathological and Immunohistochemical Analyses of Tissues
Histopathological Analysis of Tissues
Immunohistochemical Analysis of Tissues
2.4. Statistical Analysis
3. Results and Discussion
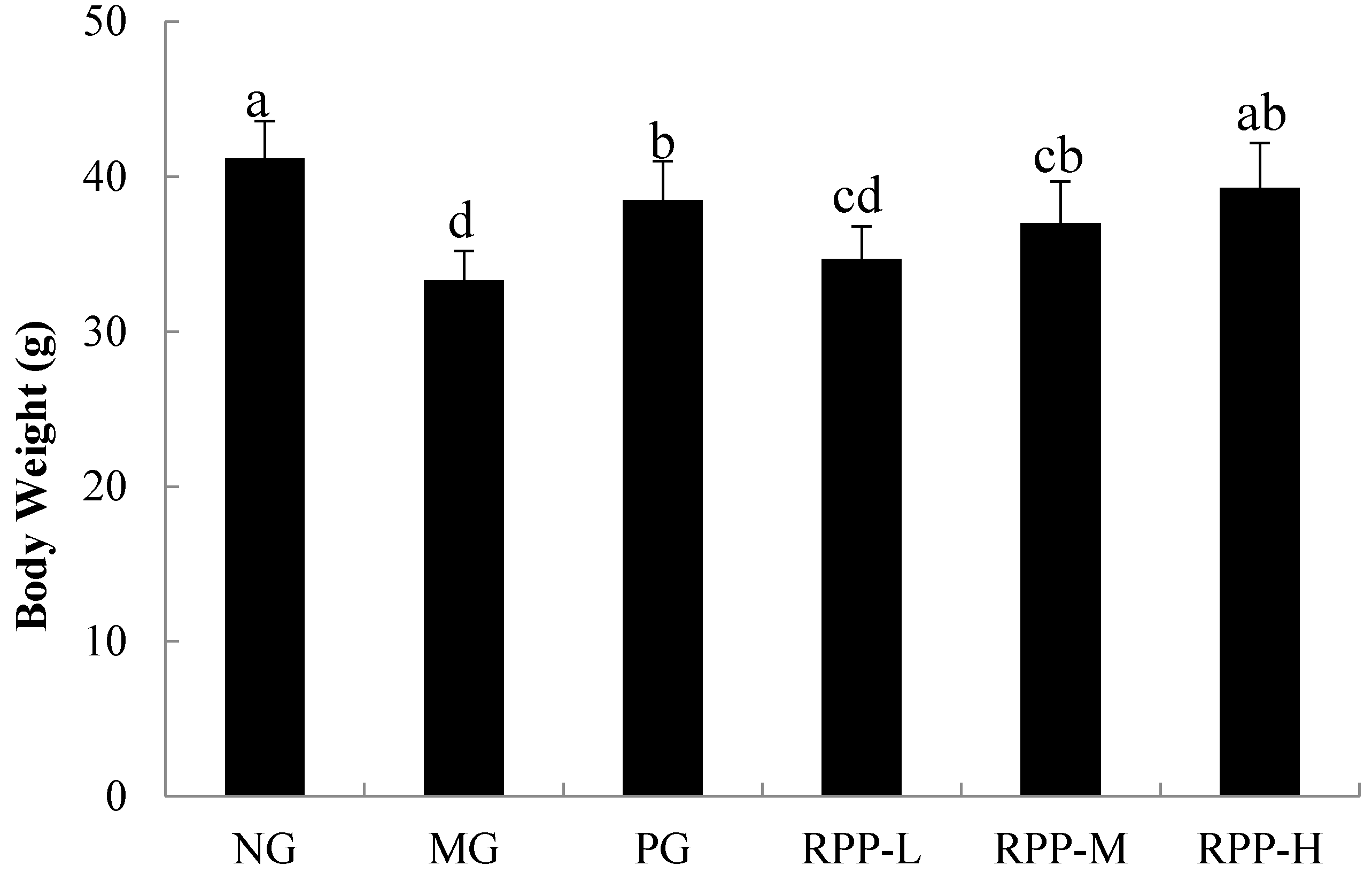
3.1. Body Weight
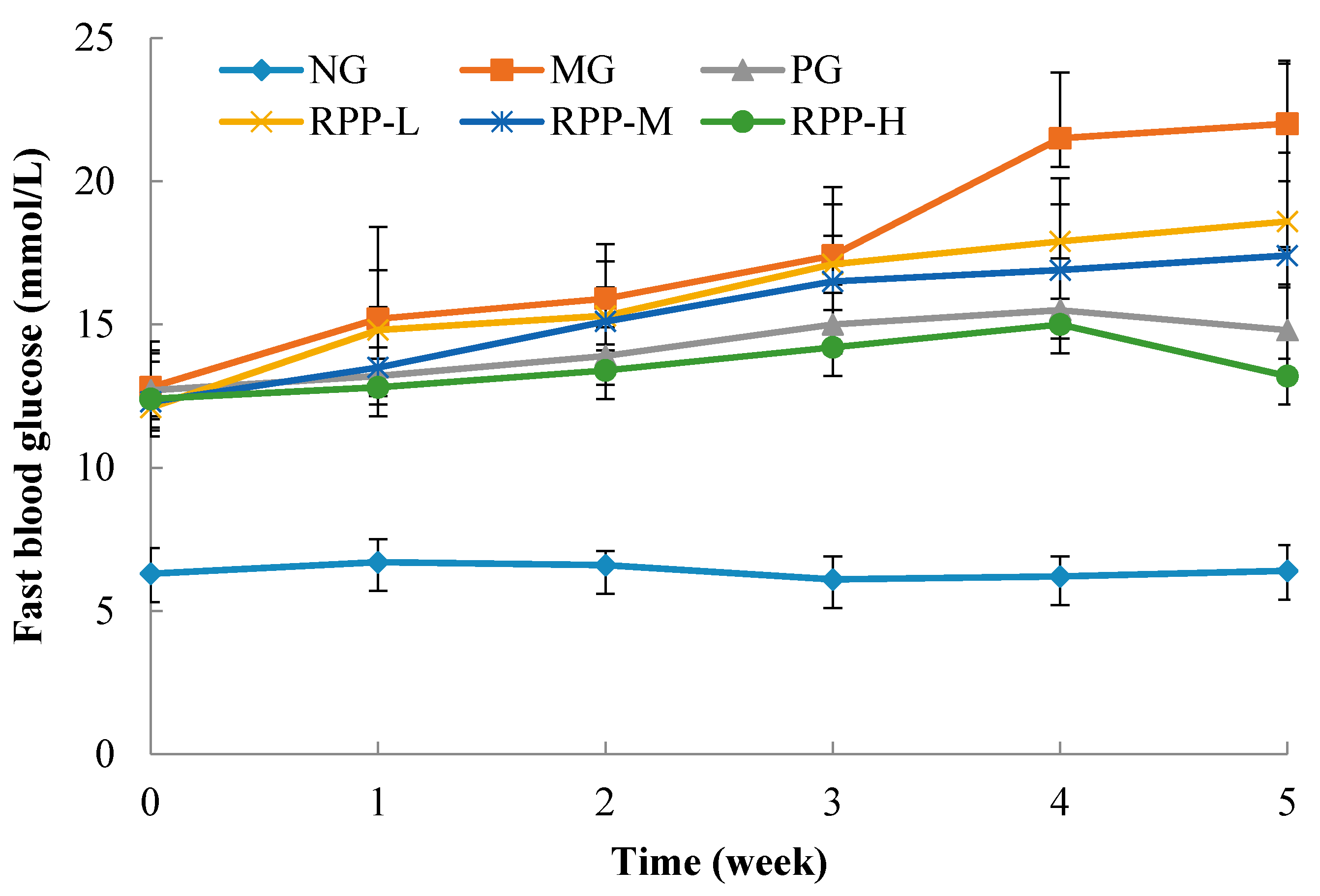
3.2. Fasting Blood Glucose (FBG)
3.3. Serum Biochemical Indicators
3.4. Biochemical Indicators in the Liver
4. Histopathology
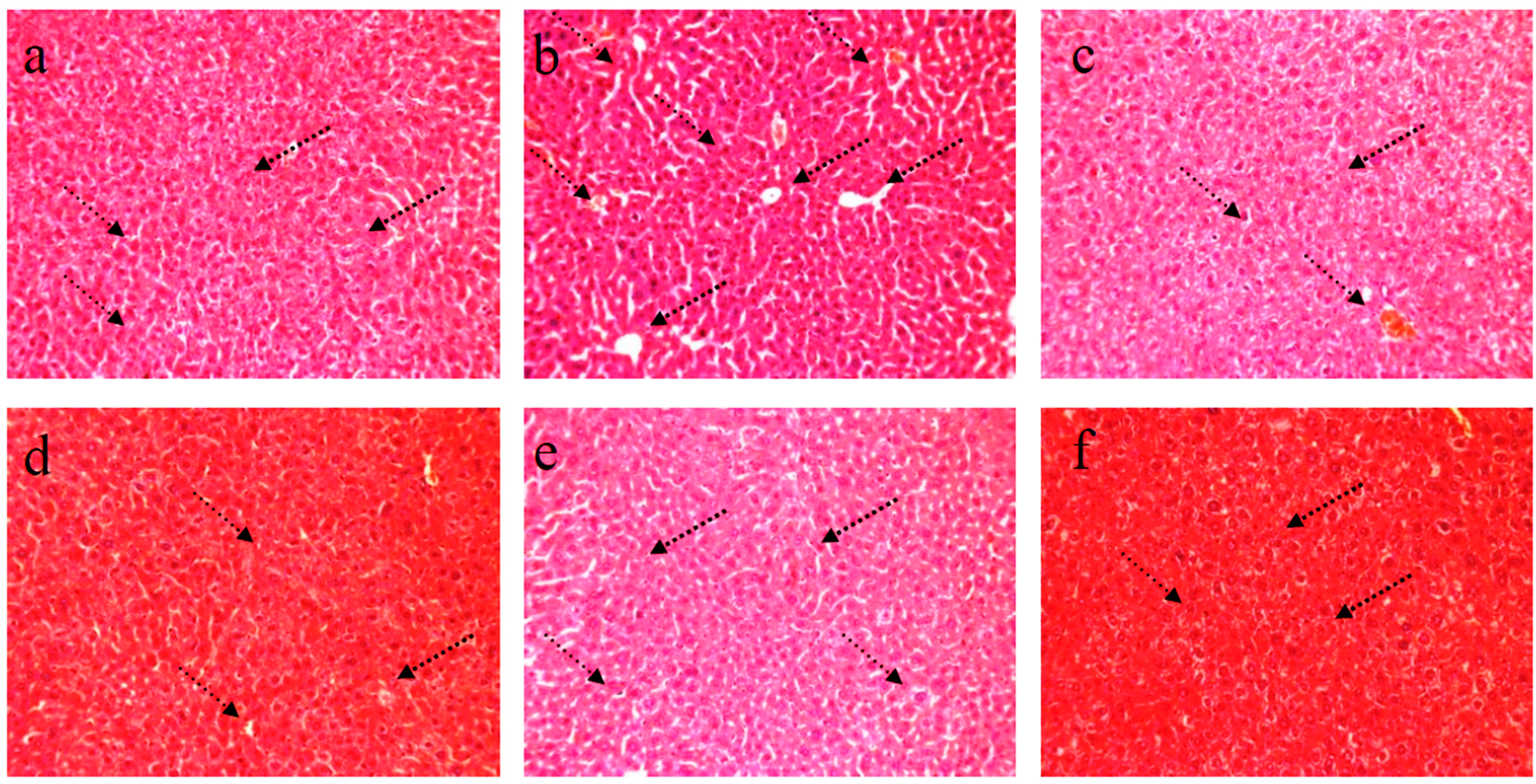
4.1. HE staining of Liver
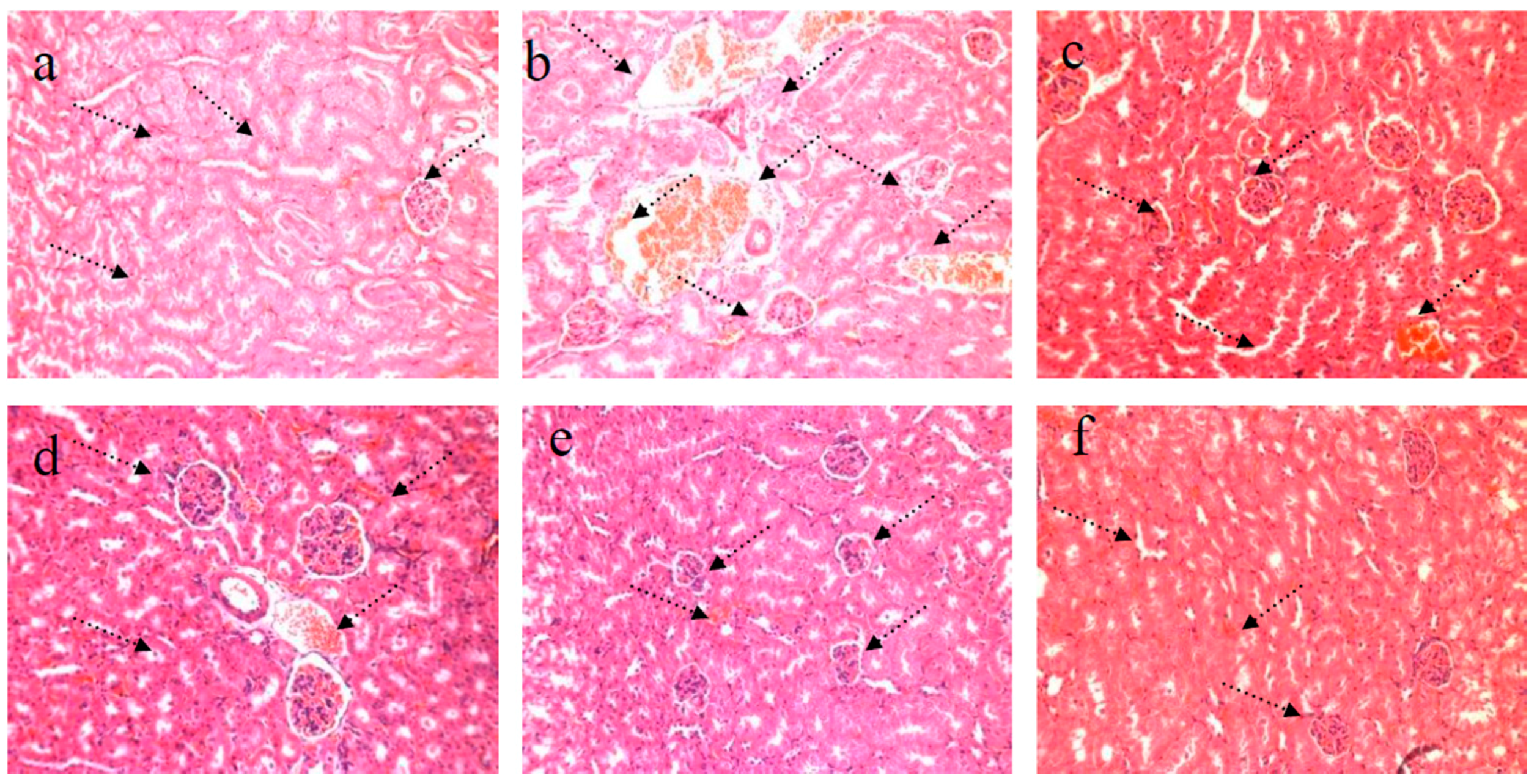
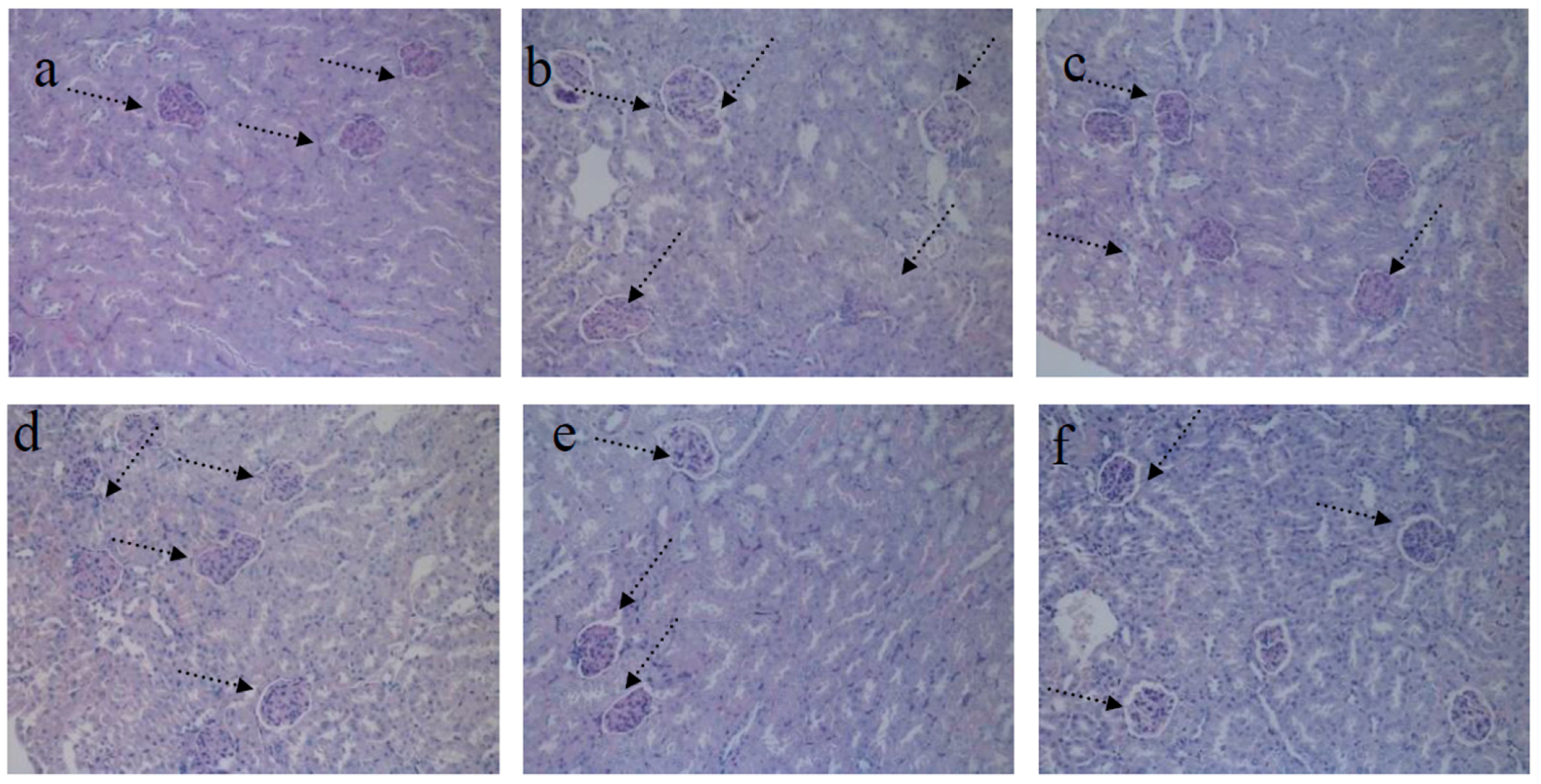
4.2. HE Staining of Kidney
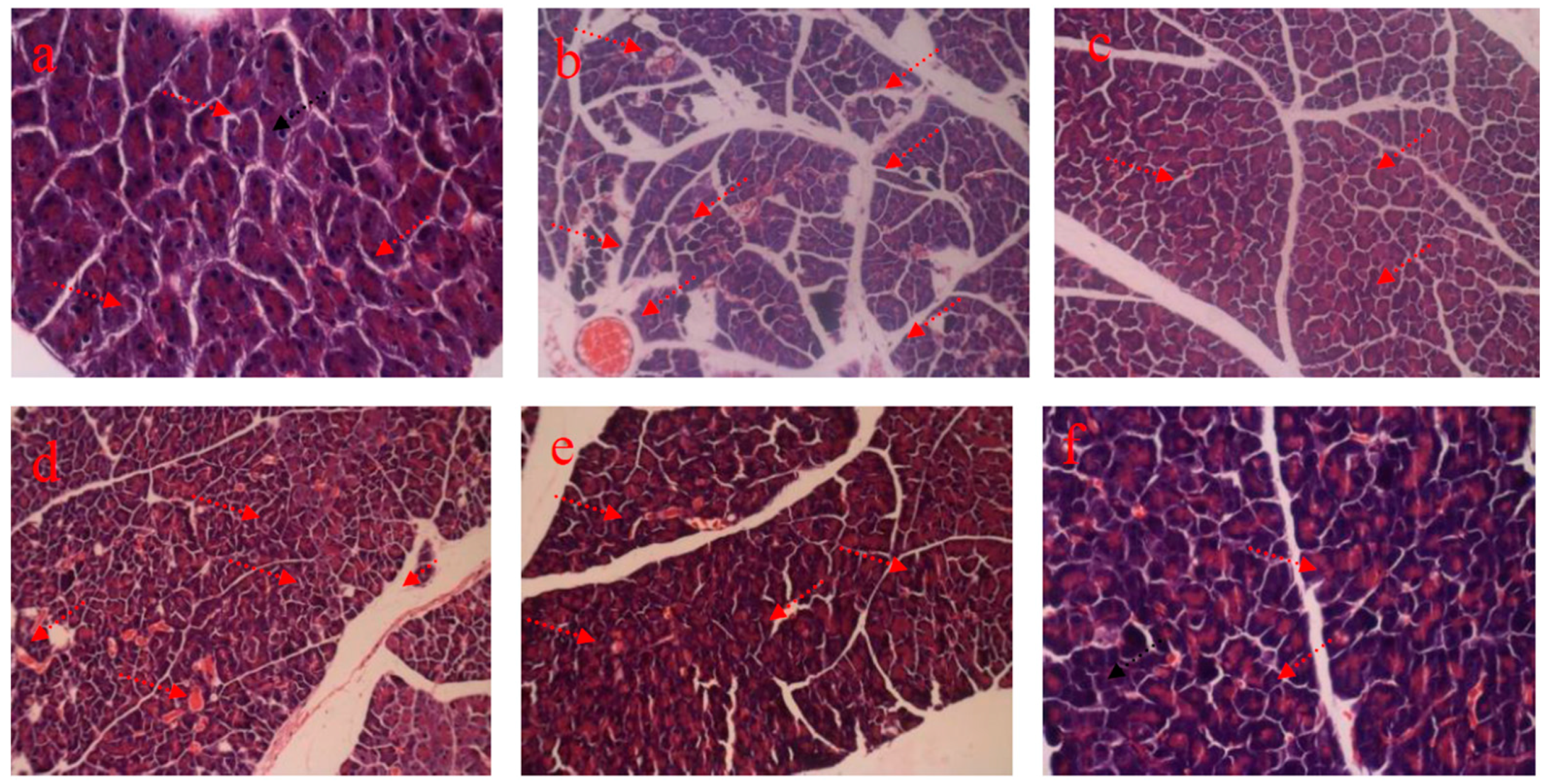
4.3. HE Staining of Pancreas
4.4. PAS Staining of Kidney
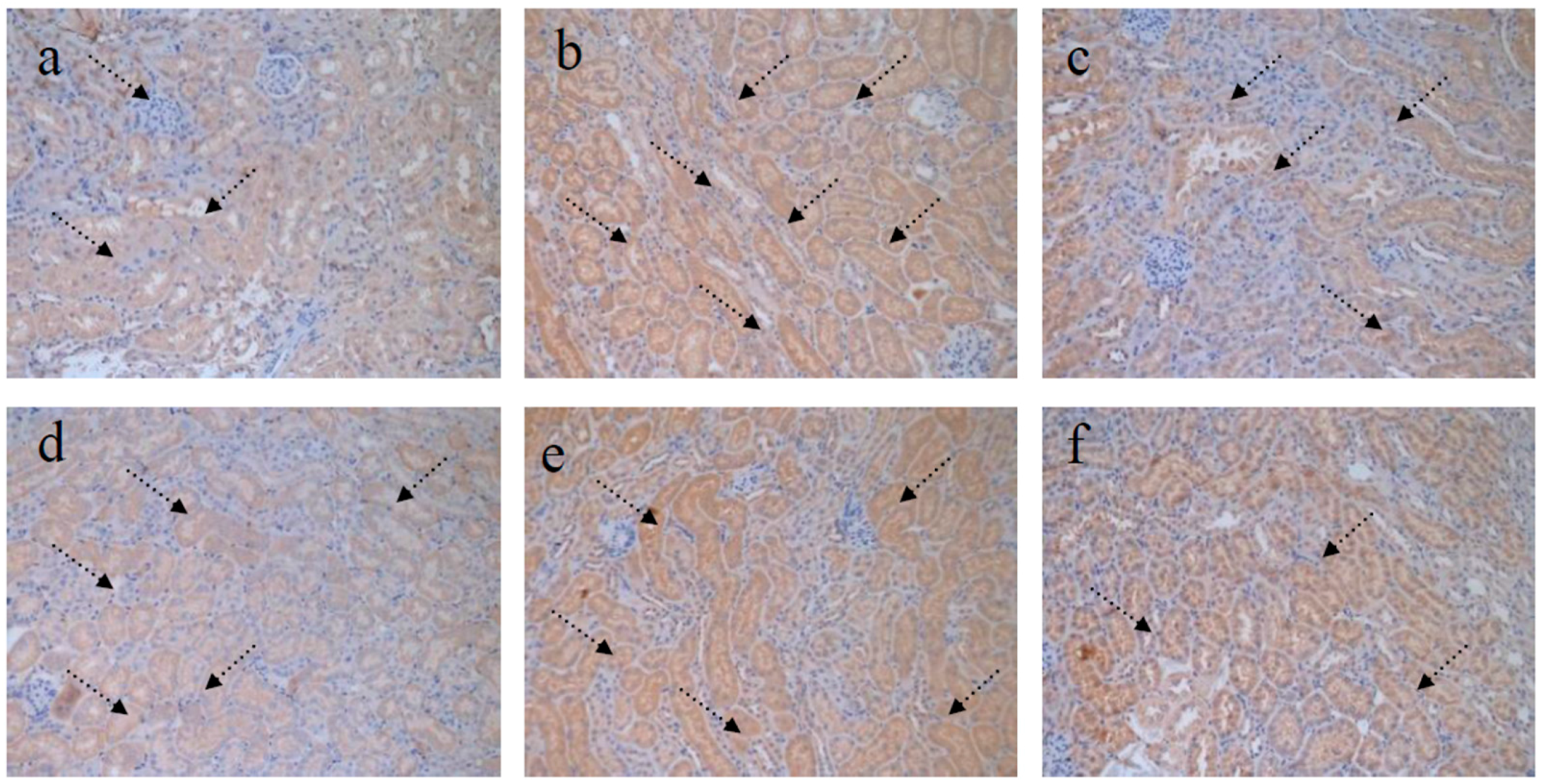
4.5. TGF-β1 Staining of Kidney
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gopal, S.S.; Lakshmi, M.J.; Sharavana, G.; Sathaiah, G.; Sreerama, Y.N.; Baskaran, V. Lactucaxanthin a potential anti-diabetic carotenoid from lettuce (Lactuca sativa) inhibits α-amylase and α-glucosidase activity in vitro and in diabetic rats. Food Funct. 2017, 8, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.J.; Zheng, J.; Wu, J.; Qiao, H.Q.; Chang, Q.; Niu, Y.; Sun, T.; Li, Y.X.; Yu, J.Q. Beneficial effects of Lyciumbarbarum polysaccharide on spermatogenesis by improving antioxidant activity and inhibiting apoptosis in streptozotocin-induced diabetic male mice. Food Funct. 2017, 8, 1215–1226. [Google Scholar] [CrossRef] [PubMed]
- Prince, P.S.M.; Kamalakkannan, N.; Menon, V.P. Antidiabetic and antihyperlipidemic effect of alcoholic Syzigiumcumini seeds in alloxan induced diabetic albino rats. J. Ethnopharm. 2004, 91, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wei, Z.; Ma, H.; Cai, A.; Liu, Y.; Sun, J.; DaSilva, N.A.; Johnson, S.L.; Kirschenbaum, L.; Cho, B.P.; et al. Anti-glycation and anti-oxidative effects of a phenolic-enriched maple syrup extract and its protective effects on normal human colon cells. Food Funct. 2017, 8, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Huang, S.; Cai, S.; Cao, J.; Han, P. Digestion property and synergistic effect on biological activity of purple rice (Oryza sativa L.) anthocyanins subjected to a simulated gastrointestinal digestion in vitro. Food Res. Int. 2015, 78, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Alkan, D.; Yemenicioglu, A. Potential application of natural phenolic antimicrobials and edible film technology against bacterial plant pathogens. Food Hydrocol. 2016, 55, 1–10. [Google Scholar] [CrossRef]
- Sun, L.; Guo, Y.; Zhang, Y.; Zhuang, Y. Antioxidant and Anti-tyrosinase Activities of Phenolic Extracts from Rape Bee Pollen and Inhibitory Melanogenesis by cAMP/MITF/TYR Pathway in B16 Mouse Melanoma Cells. Front. Pharmacol. 2017, 8, 104. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Sun, X.; Tian, C.; Zheng, Y.; Zheng, C.; Zhan, J. Chemical composition and hepatoprotective effects of polyphenols extracted from the stems and leaves of Sphallerocarpus gracilis. J. Funct. Foods 2015, 18, 673–683. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Kim, J.-H.; Kim, S.H.; Oh, J.Y.; Seo, W.D.; Kim, K.-M.; Jung, J.-C.; Jung, Y.-S. Barley Sprouts Extract Attenuates Alcoholic Fatty Liver Injury in Mice by Reducing Inflammatory Response. Nutrients 2016, 8, 440. [Google Scholar] [CrossRef] [PubMed]
- Jeszka-Skowron, M.; Flaczyk, E.; Jeszka, J.; Krejpcio, Z.; Król, E.; Buchowski, M.S. Mulberry leaf extract intake reduceshyper glycaemia in streptozotocin (STZ)-induced diabetic rats fed high-fat diet. J. Func. Foods 2014, 8, 9–17. [Google Scholar] [CrossRef]
- Cherbal, A.; Kebieche, M.; Yilmaz, E.M.; Aydoğmuş, Z.; Benzaouia, L.; Benguessoum, M.; Benkedidah, M.; Madani, K. Antidiabetic and hypolipidemic activities of Algerian Pistachia lentiscus L. leaves extract in alloxan-induced diabetic rats. S. Afr. J. Bot. 2017, 108, 157–162. [Google Scholar] [CrossRef]
- Liu, Z.; Zhai, J.; Han, N.; Yin, J. Assessment of anti-diabetic activity of the aqueous extract of leaves of Astilboidestabularis. J. Ethnopharm. 2016, 194, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Boaventura, B.C.B.; Pietro, P.F.D.; Kleina, G.A.; Stefanuto, A.; de Morais, E.C.; de Andrade, F.; Wazlawik, E.; da Silva, E.L. Antioxidant potential of mate tea (Ilex paraguariensis) in type 2 diabetic mellitus and pre-diabetic individuals. J. Func. Foods 2013, 5, 1057–1064. [Google Scholar] [CrossRef]
- Chung, I.M.; Kim, E.H.; Yeo, M.A.; Kim, S.J.; Seo, M.C.; Moon, H.I. Antidiabetic effects of three Korean sorghum phenolic extracts in normal and streptozotocin-induced diabetic rats. Food Res. Int. 2011, 44, 127–132. [Google Scholar] [CrossRef]
- Vadivel, V.; Biesalski, H.K. Contribution of phenolic compounds to the antioxidant potential and type II diabetes related enzyme inhibition properties of Pongamia pinnata L. Pierre seeds. Process Biochem. 2011, 46, 1973–1980. [Google Scholar] [CrossRef]
- Taha, H.; Ary, A.; Paydar, M.; Looi, C.Y.; Wong, W.F.; Murthy, C.R.V.; Noordin, M.I.; Ali, H.M.; Mustafa, A.M.; Hadi, A.H.A. Upregulation of insulin secretion and down regulationof pro-inflammatory cytokines, oxidative stress and hyperglycemiain STZ-nicotinamide-induced type 2 diabetic rats by Pseuduvaria monticola bark extract. Food Chem. Toxicol. 2014, 66, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.P.; Zhang, H.L.; Zhuang, Y.L. Preparation of free, soluble conjugate, and insoluble-bound phenolic compounds from peels of rambutan (Nephelium lappaceum) and evaluation of antioxidant activities in vitro. J. Food Sci. 2012, 77, C198–C204. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.; Ma, Q.; Guo, Y.; Sun, L. Purification and identification of rambutan (Nephelium lappaceum) peel phenolics with evaluation of antioxidant and antiglycation activities in vitro. Int. J. Food Sci. Technol. 2017. [Google Scholar] [CrossRef]
- Zhuang, Y.; Ma, Q.; Guo, Y.; Sun, L. Protective effects of rambutan (Nephelium lappaceum) peel phenolics on H2O2-induced oxidantive damages in HepG2 cells and d-galactose-induced aging mice. Food Chem. Toxicol. 2017. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Oral administration of Lactobacillus rhamnosus CCFM0528 improves glucose tolerance and cytokine secretion on high-fat-fed, streptozotocin-induced type 2 diabetic mice. J. Func. Foods 2014, 10, 318–326. [Google Scholar] [CrossRef]
- Priscilla, D.H.; Jayakumar, M.; Thirumurgan, K. Flavanone naringenin: An effective antihyperglycemic and antihyperlipidemic nutraceutical agent on high fat diet fed streptozotocin induced type 2 diabetic rats. J. Func. Foods 2015, 14, 363–373. [Google Scholar] [CrossRef]
- Mehenni, C.; Atmani-kilani, D.; Dumarcay, S.; Perrin, D.; Gérardin, P.; Atmani, D. Hepatoprorective and antidiabetic effects of Pistacia lentiscus leaf and fruit extracts. J. Food Drug Anal. 2016, 24, 653–669. [Google Scholar] [CrossRef]
- Xu, J.; Wang, Y.; Xu, D.S.; Ruan, K.F.; Feng, Y.; Wang, S. Hypoglycemic effects of MDG-1, a polysaccharide derived from Ophiopogon japonicas, in the ob/ob mouse model of type 2 diabetes mellitus. Int. J. Biol. Macromol. 2011, 49, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ji, D.; Zhong, S.; Lin, T.; Lv, Z. Hypoglycemic effect of deoxynojirimycin-polysaccharide on high fat diet and streptozotocin-induced diabetic mice via regulation of hepatic glucose metabolism. Chemico-Bio. Int. 2015, 225, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.H.; Li, X.F.; Wang, M.N.; Zha, X.Q.; Yang, X.F.; Liu, Z.J.; Luo, Y.B.; Luo, J.P. Comparision of hypoglycemic and antioxidant effects of polysaccharides from four different Dendrobium species. Int. J. Biol. Macromol. 2014, 64, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Irudayaraj, S.S.; Sunil, C.; Duraipandiyan, V.; Ignacimuthu, S. Antidiabetic and antioxidant acticities of Toddalia asiatica (L.) Lam. leaves in streptozotoncin induced daibeitc rats. J. Ethnopharm. 2012, 143, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Yan, J.; Shen, Y.; Tang, K.; Yin, J.; Zhang, Y. Berberine improves glucose metabolism in diabetic rats by inhibition of hepatic gluconeogenesis. PLoS ONE 2011, 6, e16556. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Tang, Z.; Wang, J.; Cao, P.; Li, Q.; Shui, W.; Wang, H.; Zheng, Z.; Zhang, Y. Polysaccharide from Angelica sinensis ameliorates high-fat diet and STZ-induced hepatic oxidative stress and inflammation in diabetic mice by activating the Sirt1-AMPK pathway. J. Nutr. Biochem. 2017, 43, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, Y.; Zhuang, Y. Antiphotoaging effect and purification of an antioxidant peptide from tilapia (Oreochromis niloticus) gelatin peptides. J. Func. Foods 2013, 5, 154–162. [Google Scholar] [CrossRef]
- Arya, A.; Looi, C.Y.; Cheah, S.C.; Mustafa, M.R.; Mohd, M.A. Anti-diabetic effects of Centratherum anthelminticum seeds methanolic fraction on pancreatic cells, β-TC6 and its alleviating role in type 2 diabetic rats. J. Ethnopharmacol. 2012, 144, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Chinchansure, A.A.; Korwar, A.M.; Kulkarni, M.J.; Joshi, S.P. Recent development of plant products with anti-glycation acitvity: A review. RSC Adv. 2015, 5, 31113–31138. [Google Scholar] [CrossRef]
- Pinent, M.; Blay, M.; Bladé, M.C.; Salvadó, M.J.; Arola, L.; Ardévol, A. Grape seedderived procyanidins have an antihyperglycemic effect in streptozotocin-induced diabetic rats and insulinomimetic acitivity in insulin-sensitive cell lines. Endocrinology 2004, 145, 4985–4990. [Google Scholar] [CrossRef] [PubMed]
- Nankar, R.P.; Doble, M. Ellagic acid potentiates insulin sensitising activity of pioglitazone in L6 myotubes. J. Func. Foods 2015, 15, 1–10. [Google Scholar] [CrossRef]
- Sutariya, B.; Saraf, M. Betanin, isolated from fruits of Opuntiaelatior Mill attenuates renal fibrosis in diabetic rats through regulating oxidative stress and TGF-β pathway. J. Ethnopharmacol. 2017, 198, 432–443. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | TC (mmol/L) | TG (mmol/L) | CRE (μmol/L) | GSP (mmol/L) |
|---|---|---|---|---|
| NG | 5.90 ± 0.31d | 1.43 ± 0.11c | 10.62 ± 1.53d | 7.14 ± 0.43d |
| MG | 8.57 ± 0.43a | 2.27 ± 0.13a | 22.78 ± 1.96a | 14.08 ± 1.12a |
| PG | 6.28 ± 0.26c | 1.53 ± 0.09c | 13.93 ± 0.98c | 7.93 ± 0.62d |
| RPP-L | 7.12 ± 0.11bc | 1.74 ± 0.11b | 20.50 ± 2.04a | 13.14 ± 0.86b |
| RPP-M | 6.37 ± 0.35c | 1.54 ± 0.10c | 16.60 ± 1.14b | 10.65 ± 0.91bc |
| RPP-H | 6.01 ± 0.26cd | 1.46 ± 0.12c | 12.09 ± 0.85c | 8.72 ± 0.78c |
| Group | TP (g/g Liver) | GC (mg/g Liver) | SOD (U/mg) | GSH-Px (U/mg) | MDA (nmol/mg) |
|---|---|---|---|---|---|
| NG | 0.17 ± 0.01a | 32.37 ± 6.15a | 316.19 ± 24.06a | 105.11 ± 9.02a | 1.18 ± 0.18c |
| MG | 0.13 ± 0.02b | 16.58 ± 4.76c | 214.26 ± 17.10d | 67.13 ± 7.00b | 1.74 ± 0.37a |
| PG | 0.16 ± 0.01a | 27.70 ± 3.92b | 305.60 ± 31.74a | 101.58 ± 12.10a | 1.36 ± 0.07b |
| RPP-L | 0.13 ± 0.01b | 19.62 ± 4.11bc | 256.82 ± 31.63c | 85.18 ± 13.64ab | 1.70 ± 0.16a |
| RPP-M | 0.16 ± 0.01a | 22.22 ± 5.26b | 263.69 ± 25.11bc | 97.76 ± 6.30a | 1.45 ± 0.24b |
| RPP-H | 0.16 ± 0.01a | 25.05 ± 4.71b | 297.65 ± 23.29b | 101.83 ± 10.60a | 1.17 ± 0.16c |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Guo, Y.; Sun, L.; Zhuang, Y. Anti-Diabetic Effects of Phenolic Extract from Rambutan Peels (Nephelium lappaceum) in High-Fat Diet and Streptozotocin-Induced Diabetic Mice. Nutrients 2017, 9, 801. https://doi.org/10.3390/nu9080801
Ma Q, Guo Y, Sun L, Zhuang Y. Anti-Diabetic Effects of Phenolic Extract from Rambutan Peels (Nephelium lappaceum) in High-Fat Diet and Streptozotocin-Induced Diabetic Mice. Nutrients. 2017; 9(8):801. https://doi.org/10.3390/nu9080801
Chicago/Turabian StyleMa, Qingyu, Yan Guo, Liping Sun, and Yongliang Zhuang. 2017. "Anti-Diabetic Effects of Phenolic Extract from Rambutan Peels (Nephelium lappaceum) in High-Fat Diet and Streptozotocin-Induced Diabetic Mice" Nutrients 9, no. 8: 801. https://doi.org/10.3390/nu9080801
APA StyleMa, Q., Guo, Y., Sun, L., & Zhuang, Y. (2017). Anti-Diabetic Effects of Phenolic Extract from Rambutan Peels (Nephelium lappaceum) in High-Fat Diet and Streptozotocin-Induced Diabetic Mice. Nutrients, 9(8), 801. https://doi.org/10.3390/nu9080801