Chestnut Drying Is Critical in Determining Aspergillus flavus Growth and Aflatoxin Contamination

 , ,
, , 
Abstract
:1. Introduction
2. Results
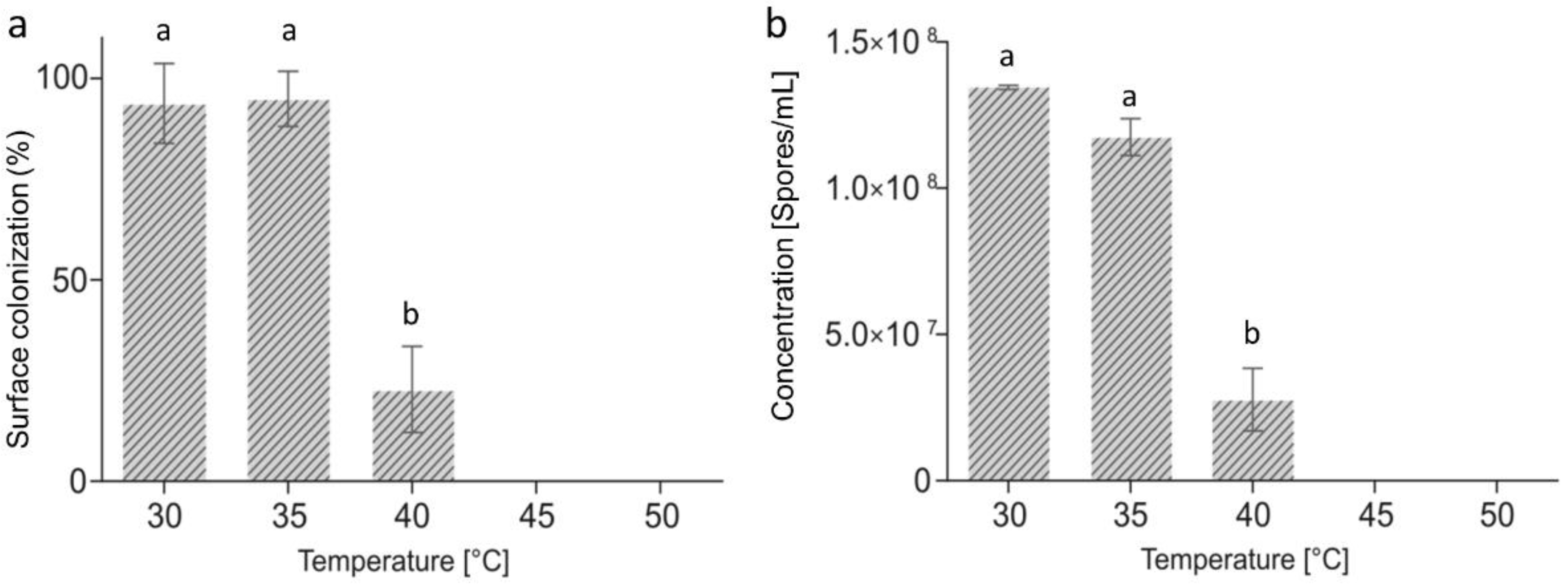
2.1. A. flavus Growth at Different Temperatures
2.2. Aflatoxin analysis
2.3. Composition
2.4. Total Phenol Content (TPC) and Antioxidant Activity
3. Discussion
4. Materials and Methods
4.1. Fungal Strain
4.2. Inoculation of Chestnuts and Fungal Spore Count
4.3. Aflatoxin Extraction and Analysis
4.4. Chestnut Composition
4.5. Total Phenol Content (TPC) and Antioxidant Activity
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations, FAOSTAT. 2014. Available online: http://www.fao.org/faostat/en/#data (accessed on 22 October 2018).
- Bertuzzi, T.; Rastelli, S.; Pietri, A. Aspergillus and Penicillium toxins in chestnuts and derived products produced in Italy. Food Control 2015, 50, 876–880. [Google Scholar] [CrossRef]
- Míguelez, J.D.L.M.; Bernardez, M.M.; Queijeiro, J.G. Composition of varieties of chestnuts from Galicia (Spain). Food Chem. 2004, 84, 401–404. [Google Scholar] [CrossRef]
- Borges, O.; Gonçalves, B.; De Carvalho, J.L.S.; Correia, P.; Silva, A.P. Nutritional quality of chestnut (Castanea sativa Mill.) cultivars from Portugal. Food Chem. 2008, 106, 976–984. [Google Scholar] [CrossRef]
- Peña-Méndez, E.M.; Hernández-Suárez, M.; Díaz-Romero, C.; Rodríguez-Rodríguez, E. Characterization of various chestnut cultivars by means of chemometrics approach. Food Chem. 2008, 107, 537–544. [Google Scholar] [CrossRef]
- Üstün, N.; Tosun, Y.; Serdar, U. Technological properties of chestnut varieties grown in Erfelek district of Sinopy city. Acta Hortic. 1999, 494, 107–110. [Google Scholar]
- Barreira, J.C.; Casal, S.; Ferreira, I.C.; Oliveira, M.B.P.; Pereira, J.A. Nutritional, fatty acid and triacylglycerol profiles of Castanea sativa Mill. cultivars: A compositional and chemometric approach. J. Agric. Food Chem. 2009, 57, 2836–2842. [Google Scholar] [CrossRef]
- Attanasio, G.; Cinquanta, L.; Albanese, D.; Di Matteo, M. Effects of drying temperatures on physico-chemical properties of dried and rehydrated chestnuts (Castanea sativa). Food Chem. 2004, 88, 583–590. [Google Scholar] [CrossRef]
- Moreira, R.; Chenlo, F.; Chaguri, L.; Vázquez, G. Air drying and colour characteristics of chestnuts pre-submitted to osmotic dehydration with sodium chloride. Food Bioprod. Process. 2011, 89, 109–115. [Google Scholar] [CrossRef]
- Nazzaro, M.; Barbarisi, C.; La Cara, F.; Volpe, M.G. Chemical and biochemical characterisation of an IGP ecotype chestnut subjected to different treatments. Food Chem. 2011, 128, 930–936. [Google Scholar] [CrossRef]
- Overy, D.P.; Seifert, K.A.; Savard, M.E.; Frisvad, J.C. Spoilage fungi and their mycotoxins in commercially marketed chestnuts. Int. J. Food Microbiol. 2003, 88, 69–77. [Google Scholar] [CrossRef]
- Rodrigues, P.; Venâncio, A.; Lima, N. Mycobiota and mycotoxins of almonds and chestnuts with special reference to aflatoxins. Food Res. Int. 2012, 48, 76–90. [Google Scholar] [CrossRef]
- Prencipe, S.; Siciliano, I.; Gatti, C.; Garibaldi, A.; Gullino, M.L.; Botta, R.; Spadaro, D. Several species of Penicillium isolated from chestnut flour processing are pathogenic on fresh chestnuts and produce mycotoxins. Food Microbiol. 2018, 76, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, M.E.B.; Freire, F.D.C.O.; Maia, F.E.F.; Guedes, M.I.F.; Rondina, D. Mycotoxins and their effects on human and animal health. Food Control 2014, 36, 159–165. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marín, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef]
- Kosegarten, C.E.; Ramírez-Corona, N.; Mani-López, E.; Palou, E.; López-Malo, A. Description of Aspergillus flavus growth under the influence of different factors (water activity, incubation temperature, protein and fat concentration, pH, and cinnamon essential oil concentration) by kinetic, probability of growth, and time-to-detection models. Int. J. Food Microbiol. 2016, 240, 115–123. [Google Scholar] [PubMed]
- Spadaro, D.; Garibaldi, A. Containment of mycotoxins in the food chain by using decontamination and detoxification techniques. In Practical Tools for Plant and Food Biosecurity; Gullino, M.L., Stack, J., Fletcher, J., Mumford, J., Eds.; Springer: Dordrecht, The Netherlands; pp. 163–177.
- Kabak, B.; Dobson, A.D.W.; Var, I. Strategies to prevent mycotoxin contamination of food and animal feed: A review. Crit. Rev. Food Sci. Nutr. 2006, 46, 593–619. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, I.; Spadaro, D.; Prelle, A.; Vallauri, D.; Cavallero, M.C.; Garibaldi, A.; Gullino, M.L. Use of Cold Atmospheric Plasma to Detoxify Hazelnuts from Aflatoxins. Toxins 2016, 8, 125. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, I.; Dal Bello, B.; Zeppa, G.; Spadaro, D.; Gullino, M.L. Static hot air and infrared rays roasting are efficient methods for aflatoxin decontamination on hazelnuts. Toxins 2017, 9, 72. [Google Scholar] [CrossRef]
- Udomkun, P.; Wiredu, A.N.; Nagle, M.; Müller, J.; Vanlauwe, B.; Bandyopadhyay, R. Innovative technologies to manage aflatoxins in foods and feeds and the profitability of application—A review. Food Control 2017, 76, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Pietri, A.; Rastelli, S.; Mulazzi, A.; Bertuzzi, T. Aflatoxins and ochratoxin A in dried chestnuts and chestnut flour produced in Italy. Food Control 2012, 25, 601–606. [Google Scholar] [CrossRef]
- RASFF. 2013. Available online: https://webgate.ec.europa.eu/rasff-window/portal/index.cfm?event¼notificationList (accessed on 22 October 2018).
- Prencipe, S.; Siciliano, I.; Contessa, C.; Botta, R.; Garibaldi, A.; Gullino, M.L.; Spadaro, D. Characterization of Aspergillus section Flavi isolated from fresh chestnuts and the chestnut flour process. Food Microbiol. 2018, 69, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Commission of the European Communities. Commission Regulation (EC) No 165/2010 of 26 February 2010 amending regulation (EC) No 1881/2006 setting maximum levels for certain contaminants in foodstuffs as regards aflatoxins. Off. J. Eur. Union 2010, 50, 8e12. [Google Scholar]
- Jermini, M.; Conedera, M.; Sieber, T.N.; Sassella, A.; Schärer, H. Influence of fruit treatments on perishability during cold storage of sweet chestnuts. J. Sci. Food Agric. 2006, 86, 877–885. [Google Scholar] [CrossRef]
- Marín, S.; Ramos, A.J.; Sanchis, V. Modelling Aspergillus flavus growth and aflatoxins production in pistachio nuts. Food Microbiol. 2012, 32, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Guan, X.; Xing, F.; Lv, C.; Dai, X.; Liu, Y. Effect of water activity and temperature on the growth of Aspergillus flavus, the expression of aflatoxin biosynthetic genes and aflatoxin production in shelled peanuts. Food Control 2017, 82, 325–332. [Google Scholar] [CrossRef]
- Giorni, P.; Battilani, P.; Pietri, A.; Magan, N. Effect of aw and CO2 level of Aspergillus flavus growth and aflatoxin production in high moisture maize post-harvest. Int. J. Food Microbiol. 2008, 122, 109–113. [Google Scholar] [CrossRef]
- Schmidt-Heydt, M.; Rüfer, C.E.; Abdel-Hadi, A.; Magan, N.; Geisen, R. The production of aflatoxin B1 or G1 by Aspergillus parasiticus at various combinations of temperature and water activity is related to the ratio of aflS to aflR expression. Mycotoxin Res. 2010, 26, 241–246. [Google Scholar] [CrossRef]
- Gallo, A.; Solfrizzo, M.; Epifani, F.; Panzarini, G.; Perrone, G. 2016 Effect of temperature and water activity on gene expression and aflatoxin biosynthesis in Aspergillus flavus on almond medium. Int. J. Food Microbiol. 2016, 217, 162–169. [Google Scholar] [CrossRef]
- Neri, L.; Dimitri, G.; Sacchetti, G. Chemical composition and antioxidant activity of cured chestnuts from three sweet chestnut (Castanea sativa Mill.) ecotypes from Italy. J. Food Compos. Anal. 2010, 23, 23–29. [Google Scholar] [CrossRef]
- Zhu, F. Effect of processing on quality attributes of chestnut. Food Biol. Technol. 2016, 9, 1429–1443. [Google Scholar] [CrossRef]
- Schindler, A.P.; Palmer, J.G.; Eisenberg, W.V. Aflatoxin production by Aspergillus flavus as related to various temperatures. Appl. Microbiol. 1967, 15, 1006–1009. [Google Scholar] [PubMed]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 2nd ed.; Blackie Academic and Professional: London, UK, 1997. [Google Scholar]
- Magan, N.; Aldred, D.; Sanchis, B. The role of spoilage fungi in seed deterioration. In Fungal Biotechnology in Agricultural Food and Environmental Applications; Arora, D.K., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 311–323. [Google Scholar]
- Lahouar, A.; Marin, S.; Crespo-Sempere, A.; Saïd, S.; Sanchis, V. Effects of temperature, water activity and incubation time on fungal growth and aflatoxin B1 production by toxinogenic Aspergillus flavus isolates on sorghum seeds. Rev. Argent Microbiol. 2016, 48, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Trenk, H.L.; Hartman, P.A. Effects of moisture content and temperature on aflatoxin production in corn. Appl. Microbiol. 1970, 19, 781–784. [Google Scholar] [PubMed]
- Pratiwi, C.; Rahayu, W.P.; Lioe, H.N.; Herawati, D.; Broto, W.; Ambarwati, S. The effect of temperature and relative humidity for Aspergillus flavus BIO2237 growth and aflatoxin production on soybeans. Int. Food Res. J. 2015, 22, 82–87. [Google Scholar]
- Gonçalves, B.; Borges, O.; Costa, H.S.; Bennett, R.; Santos, M.; Silva, A.P. Metabolite composition of chestnut (Castanea sativa Mill.) upon cooking: Proximate analysis, fibre, organic acids and phenolics. Food Chem. 2010, 122, 154–160. [Google Scholar] [CrossRef]
- Correia, P.; Leitão, A.; Beirão-da-Costa, M.L. The effect of drying temperatures on morphological and chemical properties of dried chestnuts flours. J. Food Eng. 2009, 90, 325–332. [Google Scholar] [CrossRef]
- Pellegrini, N.; Searfini, M.; Salvadore, S.; Del Rio, D.; Bianchi, M.; Brighenti, F. Total antioxidant capacity of spices, dried fruits, nuts, pulses, cereals and sweets consumed in Italy assessed by three different in vitro assays. Mol. Nutr. Food Res. 2009, 50, 1030–1038. [Google Scholar] [CrossRef]
- Vázquez, G.; Fontenla, E.; Santos, J.; Freire, M.S.; González-Álvarez, J.; Antorrena, G. Antioxidant activity and phenolic content of chestnut (Castanea sativa) shell and eucalyptus (Eucalyptus globulus) bark extracts Industrial. Crops Prod. 2008, 28, 279–285. [Google Scholar] [CrossRef]
- Galiñanes, C.; Freire, M.S.; González-Álvarez, J. Antioxidant activity of phenolic extracts from chestnut fruit and forest industries residues. Eur. J. Wood Prod. 2015, 73, 651–659. [Google Scholar] [CrossRef]
- Barros, A.I.R.N.A.; Nunes, F.M.; Gonçalves, B.; Bennett, R.N.; Silva, A.P. Effect of cooking on total vitamin C contents and antioxidant activity of sweet chestnuts (Castanea sativa Mill.). Food Chem. 2011, 128, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, B.; Rangel, J.; Valentão, P.; Andrade, P.B.; Pereira, J.A.; Bolke, H. Organic acids in two Portuguese chestnut (Castanea sativa Miller) varieties. Food Chem. 2007, 100, 504–508. [Google Scholar] [CrossRef]
- ISTISAN 1996/34. Available online: http://old.iss.it/binary/publ2/cont/Rapporto%2096-34.1140450878.pdf (accessed on 22 October 2018).
- 985.29 AOAC. Available online: http://www.eoma.aoac.org/methods (accessed on 22 October 2018).
- 920.39 AOAC. Available online: http://www.eoma.aoac.org/methods (accessed on 22 October 2018).
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Szőllősi, R.; Szőllősi Varga, I. Total antioxidant power in some species of Labiatae (Adaptation of FRAP method). Acta Biol. Szeged. 2002, 46, 125–127. [Google Scholar]



{kind=link}
{kind=link}
| Sample | Moisture [%] * | Fiber [g/100 g] * | Fat Content [g/100 g] * | Carbohydrates [g/100 g] * | Protein [g/100 g] * | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Harvest (Raw) | 45.50 | ±0.46 | a | 6.51 | ±0.10 | a | 0.47 | ±0.02 | a | 42.65 | ±0.06 | a | 3.51 | ±0.08 | a |
| Treatment 30 °C | 26.67 | ±1.35 | b | 5.25 | ±0.35 | a | 0.56 | ±0.09 | a | 62.23 | ±2.15 | b | 3.53 | ±0.05 | a |
| Treatment 35 °C | 19.87 | ±1.45 | c | 5.23 | ±1.21 | a | 0.56 | ±0.03 | a | 70.00 | ±1.20 | c | 3.46 | ±0.03 | a |
| Treatment 40 °C | 16.30 | ±1.60 | c | 5.32 | ±0.23 | a | 0.61 | ±0.05 | ab | 72.65 | ±4.25 | c | 3.51 | ±0.08 | a |
| Treatment 45 °C | 11.30 | ±1.40 | d | 5.42 | ±1.15 | a | 0.71 | ±0.03 | b | 77.35 | ±0.55 | d | 3.50 | ±0.08 | a |
| Treatment 50 °C | 9.62 | ±1.48 | d | 5.51 | ±0.32 | a | 0.91 | ±0.03 | c | 77.90 | ±1.39 | d | 3.66 | ±0.18 | a |
| Treatment | Total Phenol Content [mg GAE */100 g FW] * | Antioxidant Activity [µmol AAE **/100 g FW] ** | ||
|---|---|---|---|---|
| Harvest (raw) | 60.8 ± 2.02 | a | 928 ± 1.9 | c |
| Treatment 30 °C | 63.3 ± 3.44 | ab | 884 ± 1.5 | bc |
| Treatment 35 °C | 66.2 ± 6.18 | b | 755 ± 1.3 | ac |
| Treatment 40 °C | 74.3 ± 4.36 | c | 744 ± 1.5 | ac |
| Treatment 45 °C | 92.3 ± 4.72 | d | 665 ± 1.5 | ab |
| Treatment 50 °C | 131.7 ± 3.50 | e | 644 ± 1.2 | a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prencipe, S.; Siciliano, I.; Gatti, C.; Gullino, M.L.; Garibaldi, A.; Spadaro, D. Chestnut Drying Is Critical in Determining Aspergillus flavus Growth and Aflatoxin Contamination. Toxins 2018, 10, 530. https://doi.org/10.3390/toxins10120530
Prencipe S, Siciliano I, Gatti C, Gullino ML, Garibaldi A, Spadaro D. Chestnut Drying Is Critical in Determining Aspergillus flavus Growth and Aflatoxin Contamination. Toxins. 2018; 10(12):530. https://doi.org/10.3390/toxins10120530
Chicago/Turabian StylePrencipe, Simona, Ilenia Siciliano, Carlotta Gatti, Maria Lodovica Gullino, Angelo Garibaldi, and Davide Spadaro. 2018. "Chestnut Drying Is Critical in Determining Aspergillus flavus Growth and Aflatoxin Contamination" Toxins 10, no. 12: 530. https://doi.org/10.3390/toxins10120530
APA StylePrencipe, S., Siciliano, I., Gatti, C., Gullino, M. L., Garibaldi, A., & Spadaro, D. (2018). Chestnut Drying Is Critical in Determining Aspergillus flavus Growth and Aflatoxin Contamination. Toxins, 10(12), 530. https://doi.org/10.3390/toxins10120530