Acute Toxicity of the Recombinant and Native Phα1β Toxin: New Analgesic from Phoneutria nigriventer Spider Venom

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Male Protocols
2.1.1. Clinical Sings
2.1.2. Relative Organ Weight
2.1.3. Biochemical Parameters
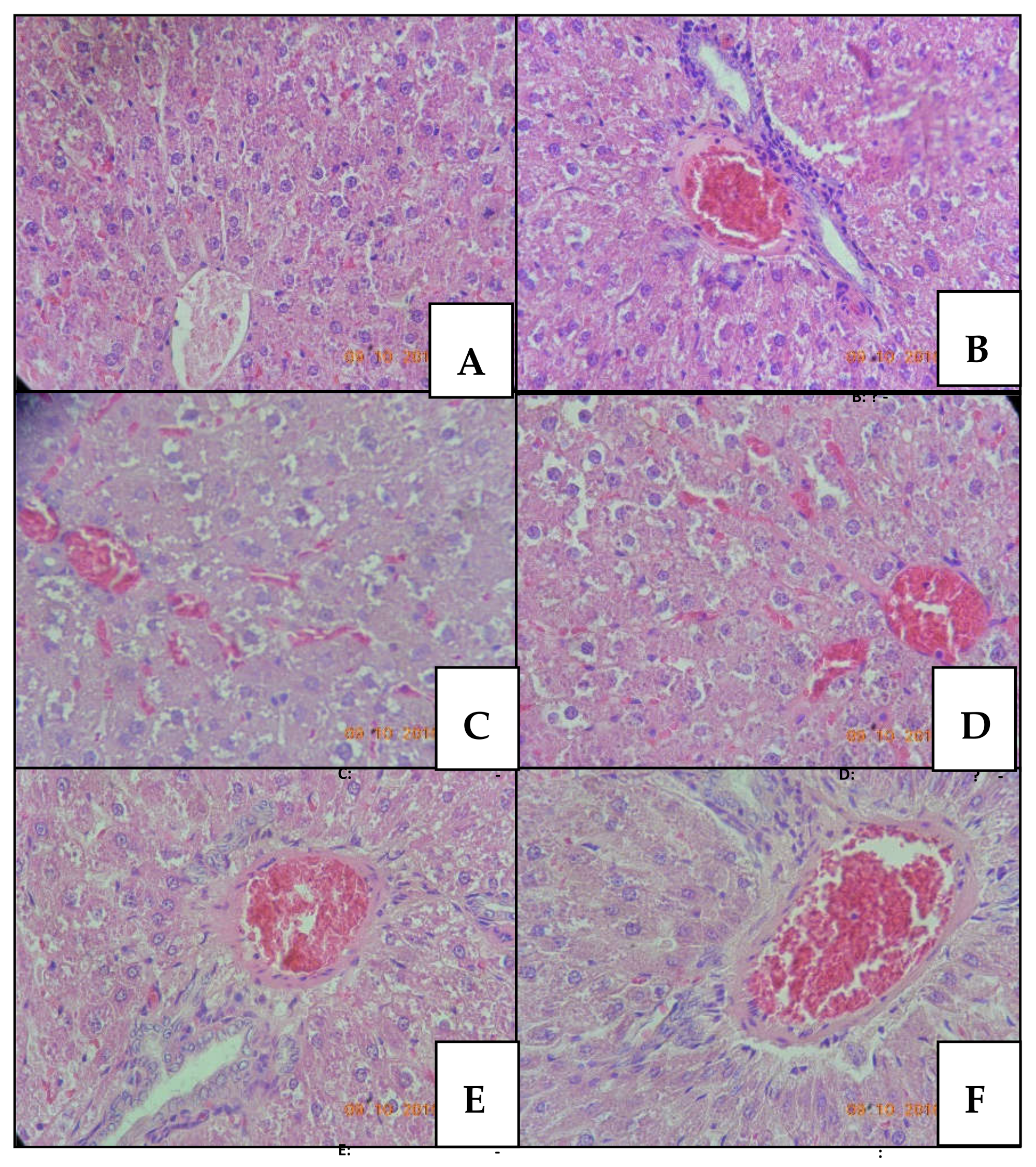
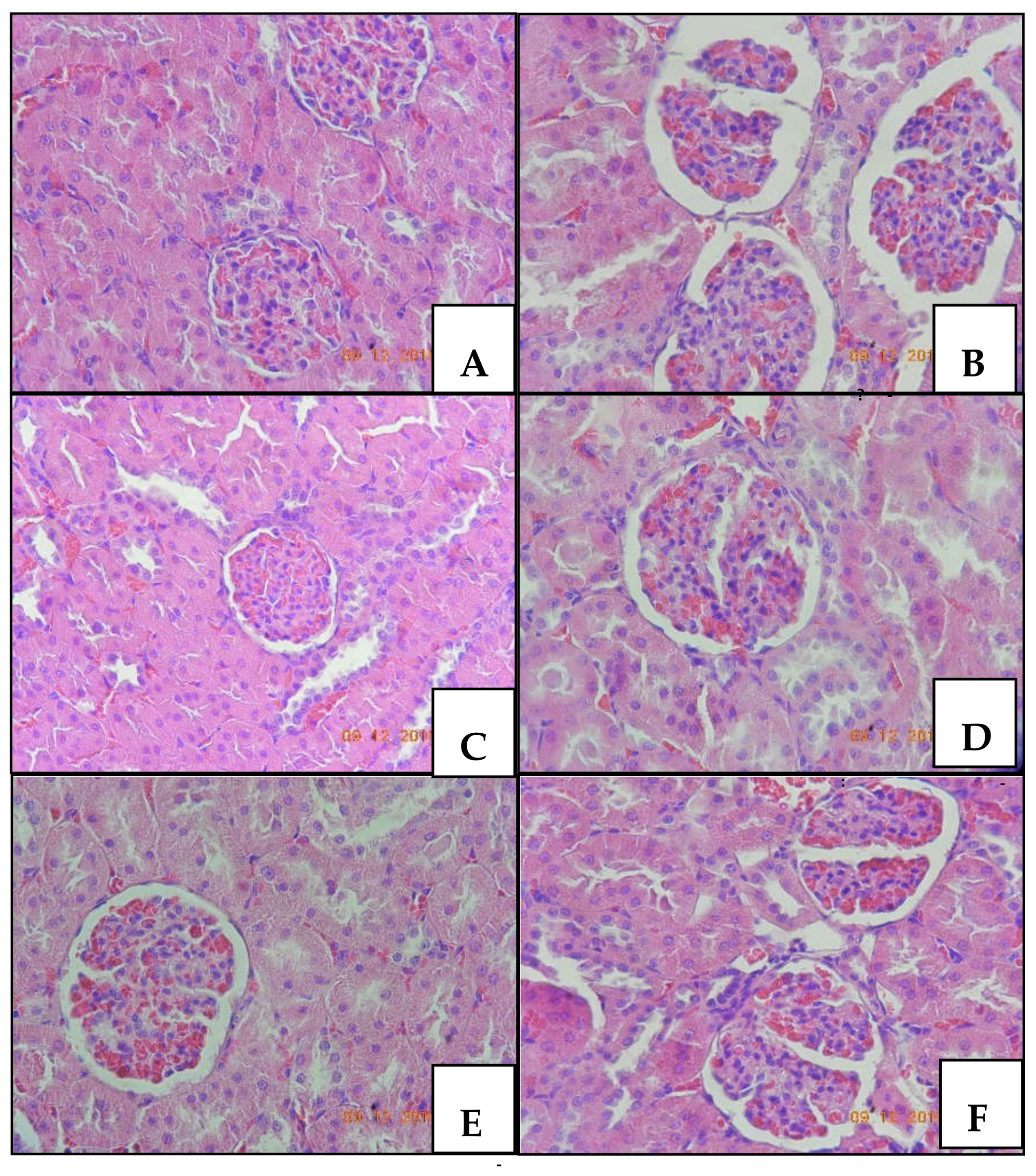
2.1.4. Histopathology
2.2. Female Protocols
2.2.1. Clinical Sings
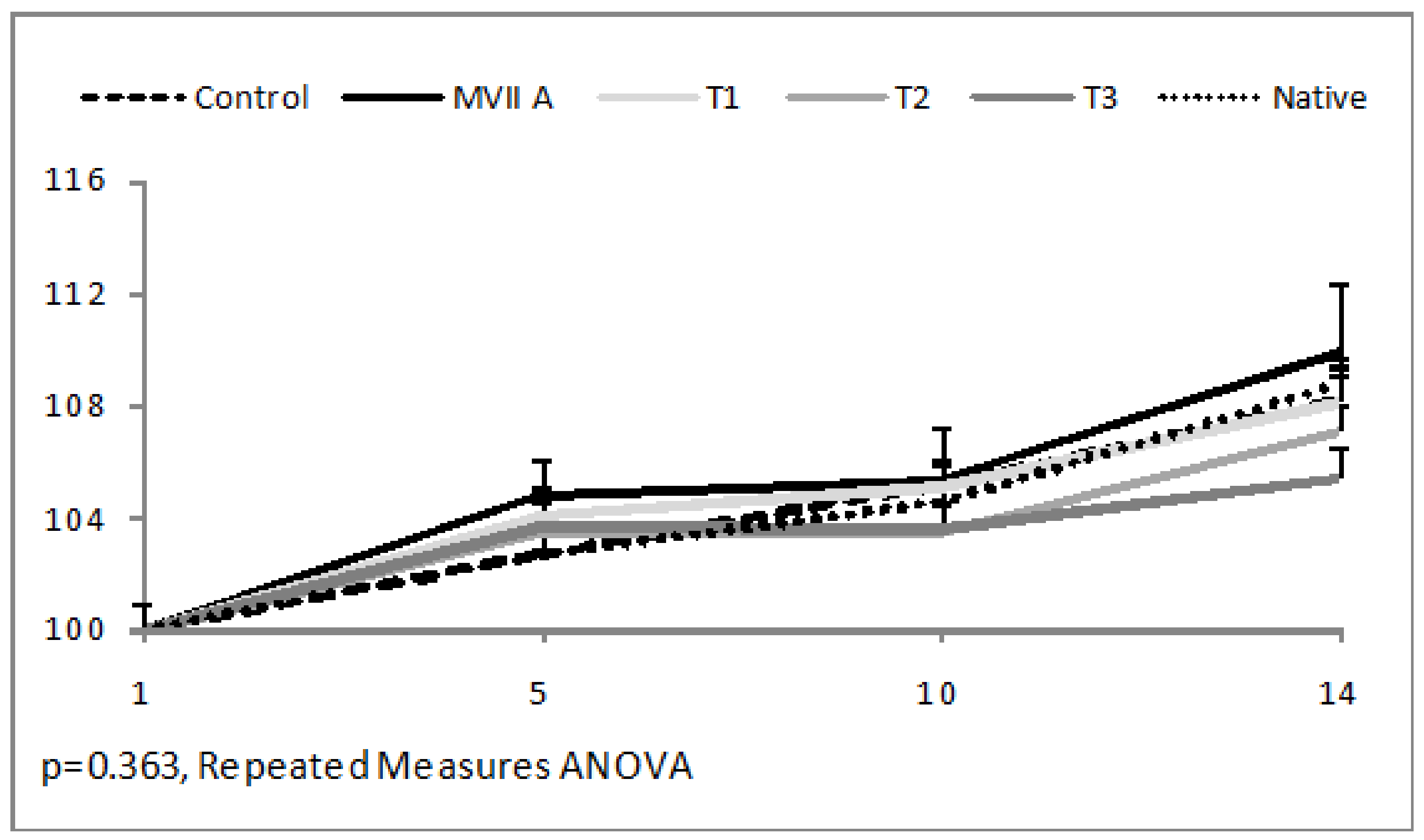
2.2.2. Bodyweight
2.2.3. Relative Organ Weight
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Drugs
5.2. Animals
5.3. Acute Toxicity Protocol
5.4. Biochemical Analysis
5.5. Histopathological Study
5.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Peigneur, S.; de Lima, M.E.; Tytgat, J. Phoneutrianigriventer venom: A pharmacological treasure. Toxicon 2018, 151, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Pineda, S.S.; Chaumeil, P.A.; Kunert, A.; Kaas, Q.; Thang, M.W.C.; Le, L.; Nuhn, M.; Herzig, V.; Saez, N.J.; Cristofori-Armstrong, B.; et al. ArachnoServer 3.0: An online resource for automated discovery, analysis and annotation of spider toxins. Bioinformatics 2018, 34, 1074–1076. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Hofling, M.A.; Tavares, J.C.; Rapôso, C. Phoneutria nigriventer venom: Action in the central nervous system. In Spider Venom; Springer: Dordrecht, The Netherlands, 2016; pp. 176–197. [Google Scholar] [CrossRef]
- Souza, A.H.; Ferreira, J.; Cordeiro, M.d.N.; Vieira, L.B.; De Castro, C.J.; Trevisan, G.; Reis, H.; Souza, I.A.; Richardson, M.; Prado, M.A.M.; et al. Analgesic effect in rodents of native toxin, in rat model of inflammatory and;neuropathic pain. Cell. Mol. Neurobiol. 2013, 33, 59–67. [Google Scholar] [CrossRef] [PubMed]
- De Souza, A.H.; Ferreira, J.; Cordeiro, M.d.N.; Vieira, L.B.; De Castro, C.J.; Trevisan, G.; Reis, H.; Souza, I.A.; Richardson, M.; Prado, M.A.M.; et al. Analgesic effect in rodents of native and recombinant phα1β toxin, a high-voltage-activated calcium channel blocker isolated from armed spider venom. Pain 2008, 140, 115–126. [Google Scholar] [CrossRef]
- De Souza, A.H.; Castro, C.J., Jr.; Rigo, F.K.; de Oliveira, S.M.; Gomez, R.S.; Diniz, D.M.; Borges, M.H.; Cordeiro, M.N.; Silva, M.A.; et al. An evaluation of the antinociceptive effects of phα1β, a neurotoxin from the spider phoneutria nigriventer, and ω-conotoxin mviia, a cone snail conus magus toxin, in rat model of inflammatory and neuropathic painic and adverse effects caused by morphine in mice. Cell Mol. Neurobiol. 2013, 33, 59–67. [Google Scholar]
- Palhares, M.R.; Silva, J.F.; Mjs, R.; Santos, D.C.; Silva-Junior, C.A.; Borges, M.H.; Ferreira, J.; Gomez, M.V.; Castro Junior, C.J. Synergistic antinociceptive effect of a calcium channel blocker and a trpv1 blocker in an acute pain model in mice. Life Sci. 2017, 182, 122. [Google Scholar] [CrossRef]
- Tonello, R.; Fusi, C.; Materazzi, S.; Marone, I.M.; De, L.F.; Benemei, S.; Gonçalves, M.C.; Coppi, E.; Castro-Junior, C.J.; Gomez, M.V. The peptide phα1β, from spider venom, acts as a trpa1 channel antagonist with antinociceptive effects in mice. Br. J. Pharmacol. 2017, 174, 57. [Google Scholar] [CrossRef]
- Rigo, F.K.; Trevisan, G.; De Pra, S.D.; Cordeiro, M.N.; Borges, M.H.; Silva, J.F.; Santa Cecilia, F.V.; de Souza, A.H.; de Oliveira Adamante, G.; Milioli, A.M.; et al. The spider toxin phalpha1beta recombinant possesses strong analgesic activity. Toxicon 2017, 133, 145–152. [Google Scholar] [CrossRef]
- Silva, R.B.; Sperotto, N.D.; Andrade, E.L.; Pereira, T.C.; Leite, C.E.; de Souza, A.H.; Bogo, M.R.; Morrone, F.B.; Gomez, M.V.; Campos, M.M. Spinal blockage of p/q- or n-type voltage-gated calcium channels modulates functional and symptomatic changes related to haemorrhagic cystitis in mice. Br. J. Pharmacol. 2015, 172, 924–939. [Google Scholar] [CrossRef]
- Silva, R.B.M.; Greggio, S.; Venturin, G.T.; da Costa, J.C.; Gomez, M.V.; Campos, M.M. Beneficial effects of the calcium channel blocker ctk 01512-2 in a mouse model of multiple sclerosis. Mol. Neurobiol. 2018, 55, 9307–9327. [Google Scholar] [CrossRef]
- Nicoletti, N.F.; Erig, T.C.; Zanin, R.F.; Roxo, M.R.; Ferreira, N.P.; Gomez, M.V.; Morrone, F.B.; Campos, M.M. Pre-clinical evaluation of voltage-gated calcium channel blockers derived from the spider p. Nigriventer in glioma progression. Toxicon 2017, 129, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Vieira, L.B.; Kushmerick, C.; Hildebrand, M.E.; Garcia, E.; Stea, A.; Cordeiro, M.N.; Richardson, M.; Gomez, M.V.; Snutch, T.P. Inhibition of high voltage-activated calcium channels by spider toxin pntx3-6. J. Pharmacol. Exp. Ther. 2005, 314, 1370–1377. [Google Scholar] [CrossRef] [PubMed]
- Herring, E.Z.; Frizon, L.A.; Hogue, O.; Mejia, J.U.; Rosenquist, R.; Bolash, R.B.; Machado, A.G.; Nagel, S.J. Long-term outcomes using intrathecal drug delivery systems in complex regional pain syndrome. Pain Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Safavi-Hemami, H.; Brogan, S.E.; Olivera, B.M. Pain therapeutics from cone snail venoms: From ziconotide to novel non-opioid pathways. J. Proteomics 2019, 190, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Goga, J.K.; Keshishian, A.; Kutzer, D.; Walters, J.K. Chronic pain management with ziconotide induces suicidal and homicidal ideations. Ann. Pharmacother. 2018, 52, 704–705. [Google Scholar] [CrossRef]
- McDowell, G.C.; Pope, J.E. IntrathecalZiconotide: Dosing and administration strategies in patients with refractory chronic pain. Neuromodulation 2016, 19, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Bilbey, D.L.; Salem, H.; Grossman, M.H. The anatomical basis of the straub phenomenon. Br. J. Pharmacol. Chemother. 1960, 15, 540–543. [Google Scholar] [CrossRef] [PubMed]
- Schmidtko, A.; Lötsch, J.; Freynhagen, R.; Geisslinger, G. Ziconotide for treatment of severe chronic pain. Lancet 2010, 375, 1569–1577. [Google Scholar] [CrossRef]
- Wang, F.; Yan, Z.; Liu, Z.; Wang, S.; Wu, Q.; Yu, S.; Ding, J.; Dai, Q. Molecular basis of toxicity of n-type calcium channel inhibitor mviia. Neuropharmacology 2016, 101, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Jayamanne, A.; Jeong, H.J.; Schroeder, C.I.; Lewis, R.J.; Christie, M.J.; Vaughan, C.W. Spinal actions of omega-conotoxins, cvid, mviia and related peptides in a rat neuropathic pain model. Br. J. Pharmacol. 2013, 170, 245–254. [Google Scholar] [CrossRef]
- Wormwood, K.L.; Ngounou Wetie, A.G.; Gomez, M.V.; Ju, Y.; Kowalski, P.; Mihasan, M.; Darie, C.C. Structural characterization and disulfide assignment of spider peptide phalpha1beta by mass spectrometry. J. Am. Soc. Mass Spectrom. 2018, 29, 827–841. [Google Scholar] [CrossRef] [PubMed]
- McGivern, J.G. Ziconotide: A review of its pharmacology and use in the treatment of pain. Neuropsychiatr. Dis. Treat. 2007, 3, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Di, L. Strategic Approaches to Optimizing Peptide ADME Properties. AAPS J. 2015, 17, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.W.; Billington, R.; Debryune, E.; Gamer, A.; Lang, B.; Carpanini, F. Recognition of adverse and nonadverse effects in toxicity studies. Toxicol. Pathol. 2002, 30, 66. [Google Scholar] [CrossRef] [PubMed]
- Farsky, S.H.; Antunes, E.; Mello, S.B. Pro and antiinflammatory properties of toxins from animal venoms. Curr. Drug Targets Inflamm. Allergy 2005, 4, 401–411. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Toxic Signs (Occurrence Interval) | Proportion of Males that Express Signs (%) n = 5/Group | |||||
|---|---|---|---|---|---|---|
| PBS 10 µL/Site | MVIIA 200 pmol/Site | CTK 01512-2 T1 200 pmol/Site | CTK 01512-2 T2 500 pmol/Site | CTK 01512-2 T3 1000 pmol/Site | Phα1β Native 500 pmol/Site | |
| Increased Ambulation (min) | 100 (0–15) | 100 (0–15) | 100 (0–15) | 100 (0–15) | 100 (0–15) | 100 (0–15) |
| Grooming (min) | 60 (0–60) | 60 (0–120) | 80 (0–360) | 100 (0–360) | 100 (0–120) | 100 (0–120) |
| Decreased Ambulation (min) | 100(15–360) | 100 (15–360) | 100 (15–360) | 100 (15–360) | 100 (15–360) | 100 (15–360) |
| Hypnosis * (min) | 80 a (120–360) | 60 a (60–360) | 60 a 120–360) | 100 a (60–360) | 60 a (120–360) | 0 b (-) |
| Decreased Touch Response (min) | 20 (60–120) | 40 (15–360) | 40 (15–360) | 0 (-) | 20 (15–240) | 40 (15–360) |
| Ataxia (min) | 0 (-) | 20 (120–360) | 0 (-) | 0 (-) | 0 (-) | 0 (-) |
| Piloerection (min) | 20 (240–360) | 40 (15–360) | 40 (30–360) | 40 (60–360) | 20 (30–240) | 100 (15–360) |
| Dyspnea * (min) | 0 a (-) | 80 b (15–360) | 80 b (15–360) | 100 b (15–360) | 100 b (15–360) | 80 b (30–360) |
| Tremors (min) | 0 (-) | 20 (15, 360) | 0 (-) | 0 (-) | 0 (-) | 0 (-) |
| Straub tail (min) | 0 (-) | 20 (15–360) | 0 (-) | 0 (-) | 0 (-) | 20 (15–60) |
| Relative Organ Weight (%) | PBS 10 µL/Site | MVIIA 200 pmol/Site | CTK 01512-2 T1 200 pmol/Site | CTK 01512-2 T2 500 pmol/Site | CTK 01512-2 T3 1000 pmol/Site | Phα1β Native 500 pmol/Site |
|---|---|---|---|---|---|---|
| Liver | 4.79 ± 0.18 | 4.70 ± 0.06 | 4.57 ± 0.17 | 4.91 ± 0.11 | 4.76 ± 0.22 | 4.63 ± 0.09 |
| Kidneys | 0.80 ± 0.02 | 0.77 ± 0.03 | 0.79 ± 0.02 | 0.81 ± 0.01 | 0.80 ± 0.02 | 0.74 ± 0.04 |
| Adrenal glands | 0.017 ± 0.001 | 0.015 ± 0.001 | 0.016 ± 0.001 | 0.016 ± 0.001 | 0.017 ± 0.001 | 0.014 ± 0.002 |
| Heart | 0.31 ± 0.01 | 0.32 ± 0.02 | 0.32 ± 0.001 | 0.32 ± 0.01 | 0.32 ± 0.01 | 0.31 ± 0.001 |
| Lung | 0.38 ± 0.01 | 0.41 ± 0.02 | 0.43 ± 0.02 | 0.42 ± 0.02 | 0.43 ± 0.02 | 0.39 ± 0.02 |
| Spleen | 0.24 ± 0.01 | 0.24 ± 0.01 | 0.25 ± 0.01 | 0.24 ± 0.01 | 0.24 ± 0.01 | 0.23 ± 0.1 |
| Biochemical Parameters | PBS 10 µL/Site | MVIIA 200 pmol/Site | CTK1512-2 T1 200 pmol/Site | CTK1512-2 T2 500 pmol/Site | CTK1512-2 T3 1000 pmol/Site | Phα1β Native 500 pmol/Site |
|---|---|---|---|---|---|---|
| Urea * (mg/dL) | 58.0 ± 4.52 a | 61.0 ± 2.38 a | 55.2 ± 2.65 a | 45.2 ± 1.93 b | 47.8 ± 1.77 b | 44.7 ± 5.80 b |
| Creatinine (mg/dL) | 0.59 ± 0.032 | 0.57 ± 0.032 | 0.54 ± 0.02 | 0.58 ± 0.027 | 0.55 ± 0.018 | 0.48 ± 0.025 |
| AST * (U/L) | 192.8 ± 22.55 a | 210.0 ± 35.55 a | 191.2 ± 39.16 a | 145.2 ± 12.54 b | 146.2 ± 20.30 a,b | 109.7 ± 5.89 b |
| ALT* (U/L) | 107.0 ± 15.32 a | 97.1 ± 12.09 a | 94.6 ± 7.73 a | 80.0 ± 3.37 b | 77.2 ± 3.24 b | 66.0 ± 2.51 b |
| ALP * (U/L) | 244.2 ± 14.94 a | 201.3 ± 8.78 b | 292.2 ± 17.33 a | 250.8 ± 32.43 a | 281.0 ± 65.85 a | 177.7 ± 21.12 b |
| Amylase (U/L) | 1381.0 ± 201.42 | 1148.6 ± 83.12 | 1159.8 ± 119.92 | 1029.2 ± 77.81 | 968.4 ± 53.64 | 958.2 ± 69.34 |
| CK (U/L) | 388.2 ± 78.51 | 225.5 ± 18.06 | 378.6 ± 70.19 | 265.8 ± 36.82 | 340.6 ± 63.30 | 282.7 ± 40.36 |
| Toxic Signs (Occurrence Interval) | Proportion of females that express the signs (%) n = 5/group | |||||
|---|---|---|---|---|---|---|
| PBS 10 µL/Site | MVIIA 200 pmol/Site | CTK 01512-2 T1 200 pmol/Site | CTK 01512-2 T2 500 pmol/Site | CTK 01512-2 T3 1000 pmol/Site | Phα1β Native 500 pmol/Site | |
| Increased Ambulation (min) | 100 (0–15) | 100 (0–15) | 100 (0–15) | 100 (0–15) | 100 (0–15) | 100 (0–15) |
| Grooming (min) | 60 (0–360) | 60 (0–30) | 100 (0–360) | 60 (0–30) | 100 (0–360) | 60 (0–15) |
| Decreased Ambulation (min) | 100 (15–360) | 100 (15–360) | 100 (15–360) | 100 (15–360) | 100 (15–360) | 100 (15–240) |
| Hypnosis (min) | 60 (240–360) | 40 (240–360) | 40 (240–360) | 40 (30–240) | 40 (60–360) | 0 (-) |
| Decreased Touch Response * (min) | 20 a (120–360) | 100 b (30–360) | 40 a (15–120) | 100 b (30–360) | 60 a (30–360) | 100 b (15–360) |
| Piloerection (min) | 80 (0–15) | 80 (15–360) | 100 (15–360) | 100 (0–360) | 40 (30–360) | 80 (0–360) |
| Dyspnea (min) | 80 (0–15) | 80 (15–360) | 100 (15–360) | 100 (30–360) | 80 (120–360) | 100(30–360) |
| Tremors * (min) | 0 a (-) | 100b (15, 360) | 40 b (15–360) | 80 b (30–360) | 40 b (240–300) | 60 b (30–360) |
| Straub tail (min) | 0 (-) | 40 (15–360) | 0 (-) | 0 (-) | 20 (300–360) | 0 (-) |
| Relative Organ Weight (%) | PBS 10 µL/Site | MVIIA 200 pmol/Site | CTK 01512-2 T1 200 pmol/Site | CTK 01512-2 T2 500 pmol/Site | CTK 01512-2 T3 1000 pmol/Site | Phα1β Native 500 pmol/Site |
|---|---|---|---|---|---|---|
| Liver | 4.29 ± 0.13 | 4.61 ± 0.21 | 4.38 ± 0.14 | 4.23 ± 0.23 | 4.45 ± 0.16 | 4.23 ± 0.13 |
| Kidneys | 0.82 ± 0.03 | 0.82 ± 0.02 | 0.81 ± 0.03 | 0.83 ± 0.05 | 0.83 ± 0.02 | 0.83 ± 0.03 |
| Adrenal glans * | 0.033 ± 0.002 a.b | 0.034 ± 0.002 a,b | 0.028 ± 0.002 a | 0.037 ± 0.001 b | 0.035 ± 0.001 a,b | 0.037 ± 0.001 b |
| Heart | 0.34 ± 0.01 | 0.35 ± 0.01 | 0.34 ± 0.01 | 0.32 ± 0.02 | 0.34 ± 0.01 | 0.32 ± 0.02 |
| Lung | 0.51 ± 0.03 | 0.50 ± 0.01 | 0.52 ± 0.05 | 0.50 ± 0.04 | 0.55 ± 0.02 | 0.50 ± 0.02 |
| Spleen * | 0.28 ± 0.01a | 0.27 ± 0.01 a,b | 0.26 ± 0.01 a,b | 0.25 ± 0.02 a,b | 0.27 ± 0.01a,b | 0.25 ± 0.01b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dallegrave, E.; Taschetto, E.; Bainy Leal, M.; Techera Antunes, F.T.; Gomez, M.V.; Hubner de Souza, A. Acute Toxicity of the Recombinant and Native Phα1β Toxin: New Analgesic from Phoneutria nigriventer Spider Venom. Toxins 2018, 10, 531. https://doi.org/10.3390/toxins10120531
Dallegrave E, Taschetto E, Bainy Leal M, Techera Antunes FT, Gomez MV, Hubner de Souza A. Acute Toxicity of the Recombinant and Native Phα1β Toxin: New Analgesic from Phoneutria nigriventer Spider Venom. Toxins. 2018; 10(12):531. https://doi.org/10.3390/toxins10120531
Chicago/Turabian StyleDallegrave, Eliane, Eliane Taschetto, Mirna Bainy Leal, Flavia Tasmim Techera Antunes, Marcus Vinicius Gomez, and Alessandra Hubner de Souza. 2018. "Acute Toxicity of the Recombinant and Native Phα1β Toxin: New Analgesic from Phoneutria nigriventer Spider Venom" Toxins 10, no. 12: 531. https://doi.org/10.3390/toxins10120531
APA StyleDallegrave, E., Taschetto, E., Bainy Leal, M., Techera Antunes, F. T., Gomez, M. V., & Hubner de Souza, A. (2018). Acute Toxicity of the Recombinant and Native Phα1β Toxin: New Analgesic from Phoneutria nigriventer Spider Venom. Toxins, 10(12), 531. https://doi.org/10.3390/toxins10120531