Deoxynivalenol Impairs Porcine Intestinal Host Defense Peptide Expression in Weaned Piglets and IPEC-J2 Cells


 , ,
, , 
Abstract
:1. Introduction
2. Results
2.1. Piglet Performance
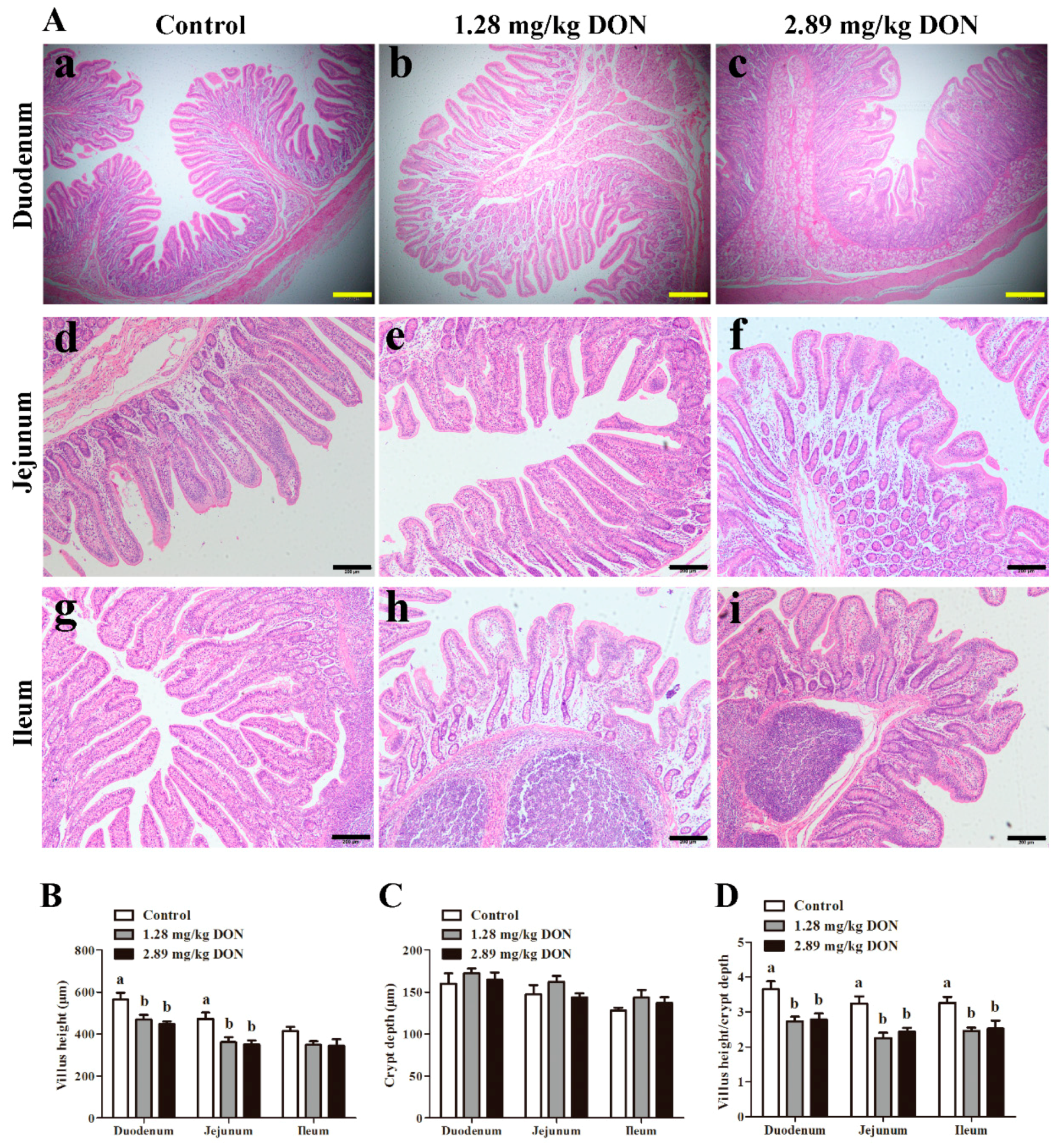
2.2. Intestinal Morphology
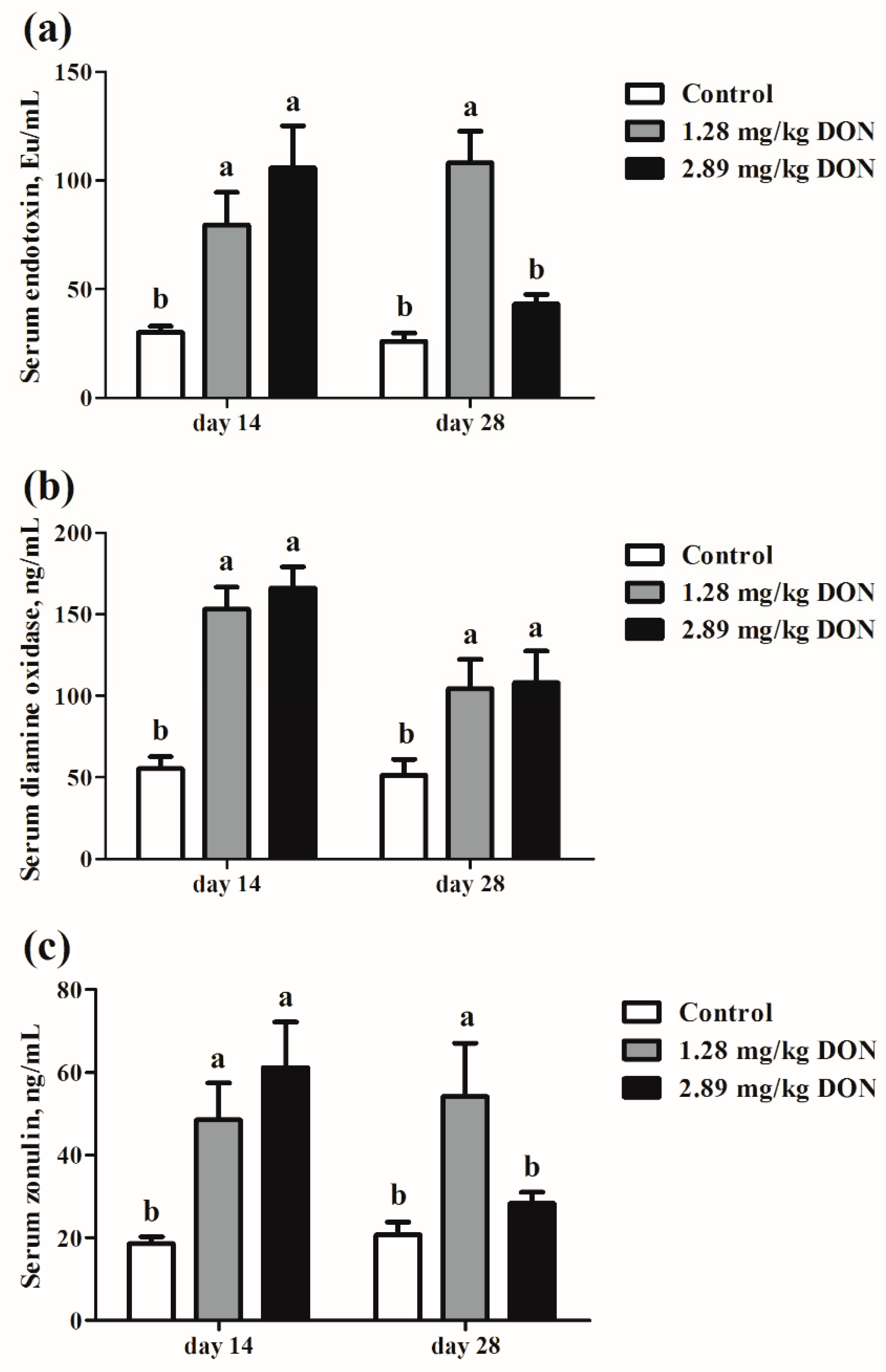
2.3. Intestinal Permeability
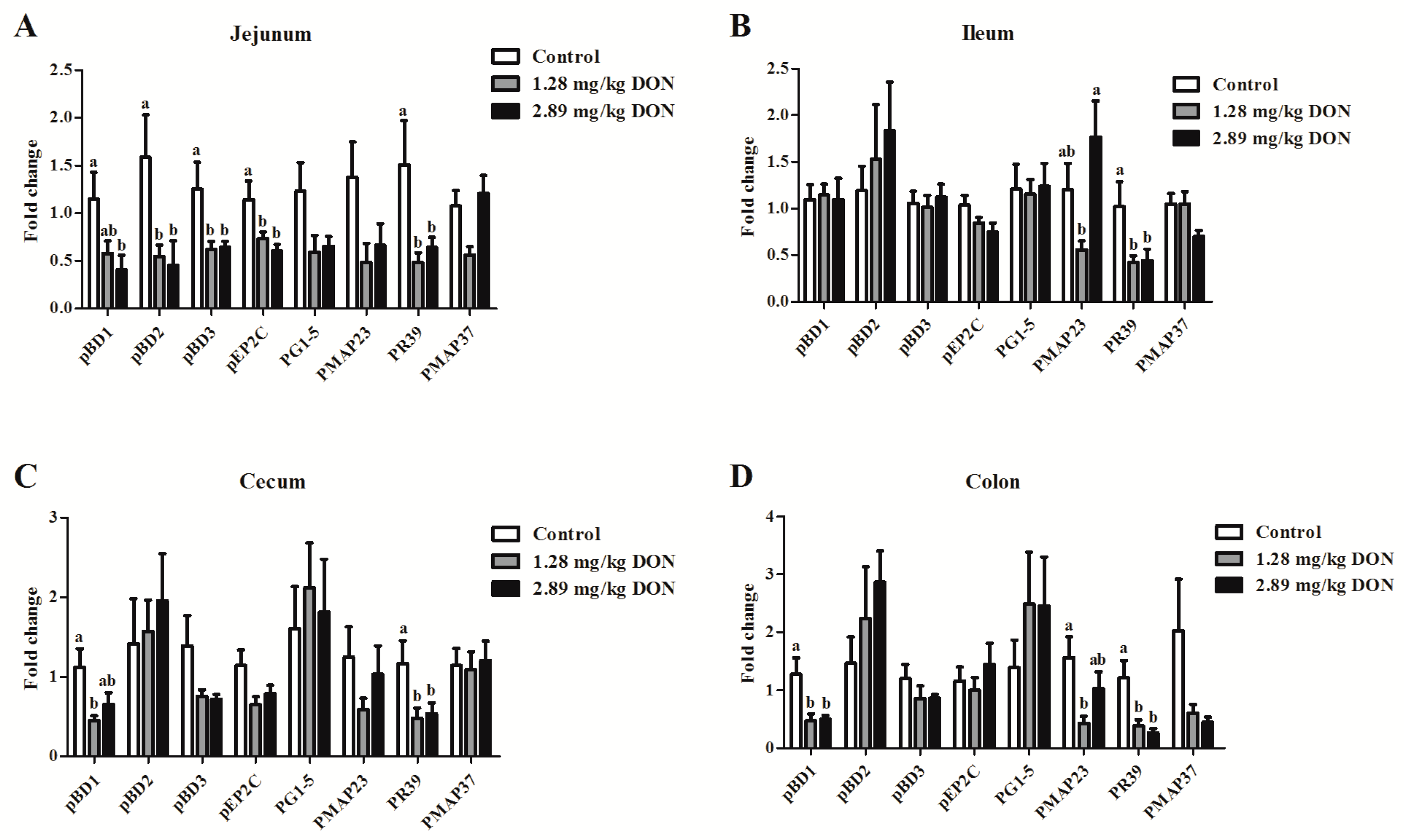
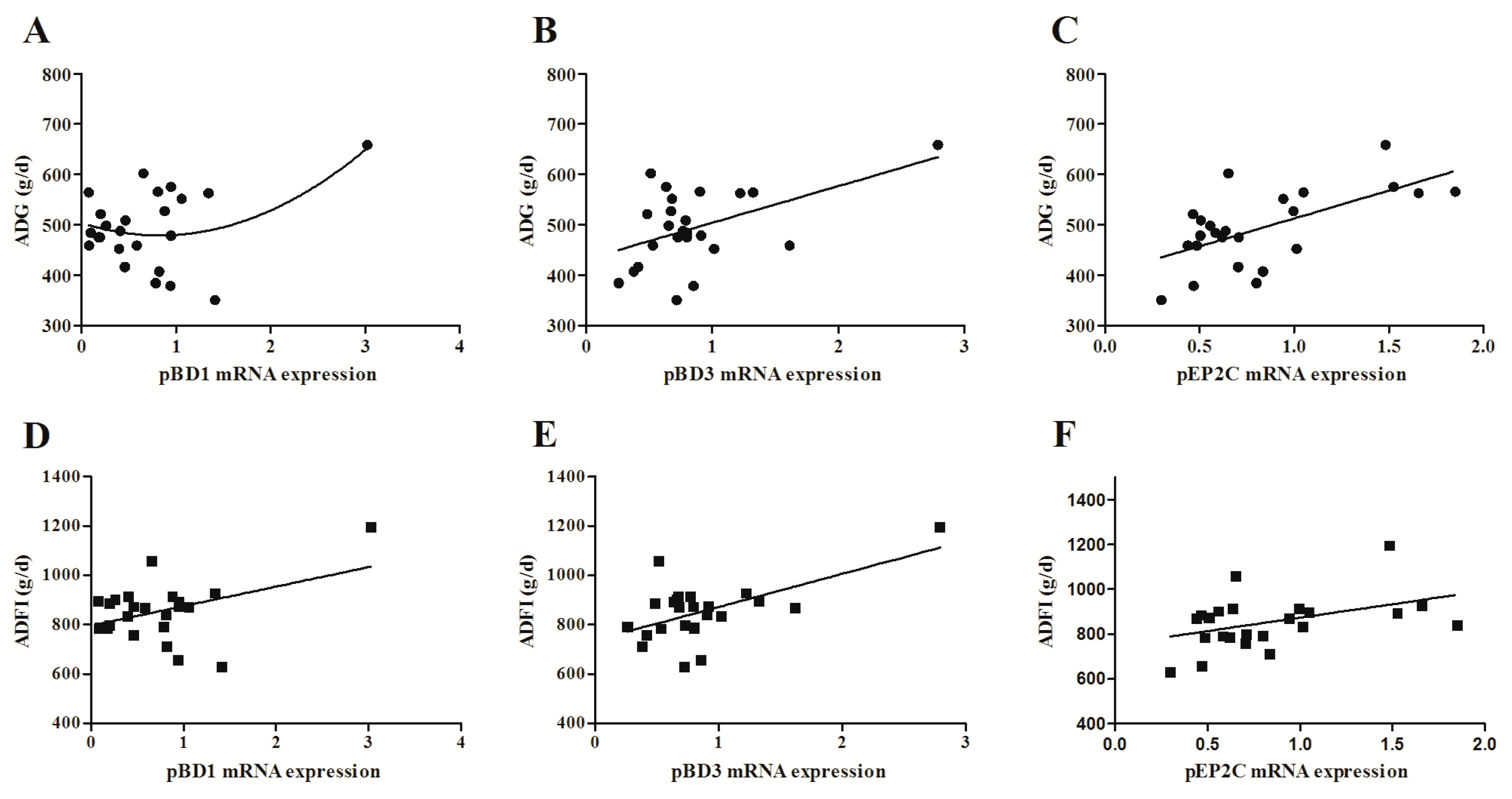
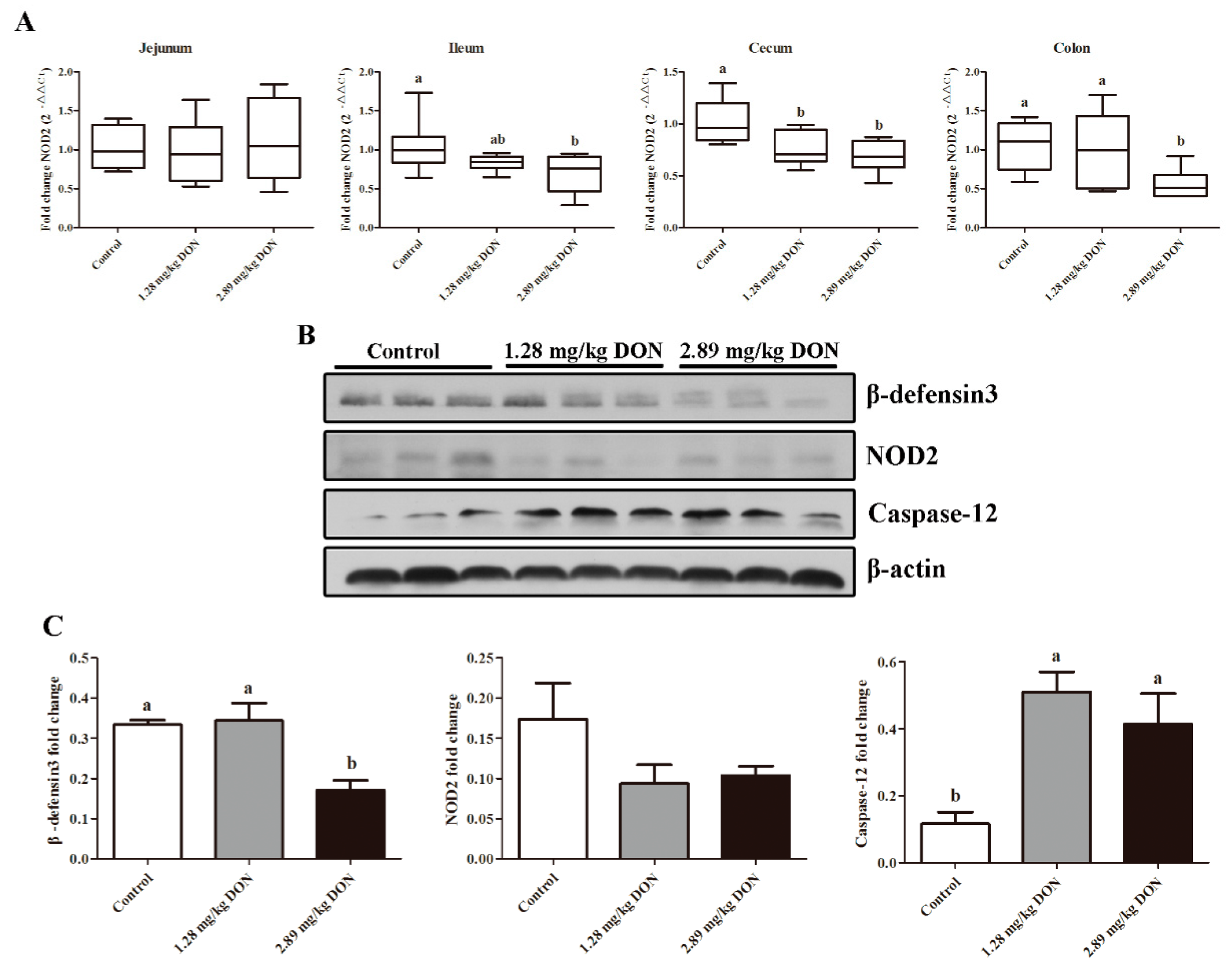
2.4. The Effect of DON on Porcine Intestinal HDPs Expression in Weaned Piglets
2.5. Activation of Caspase-12 in Jejunum
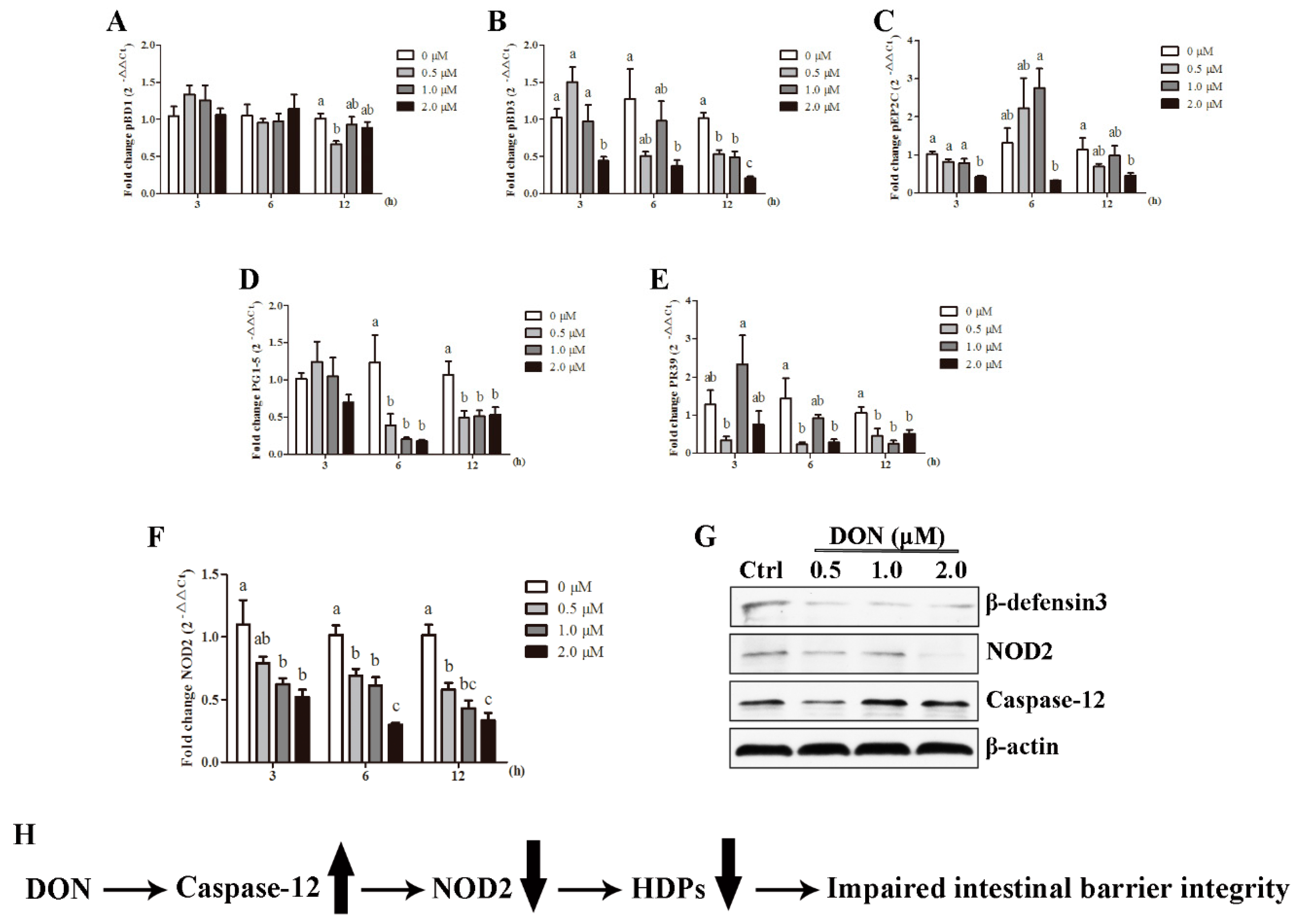
2.6. Caspase-12 Was a Critical Regulator Modulating NOD2-Mediated HDPs Production in Response to DON Exposure
3. Discussion
4. Methods and Methods
4.1. DON Production and Analysis
4.2. Animals and Experimental Design
4.3. Sample Collection
4.4. Chemical Analyses
4.5. Determination of Endotoxin, Diamine Oxidase, and Zonulin in Serum
4.6. Small Intestine Morphology
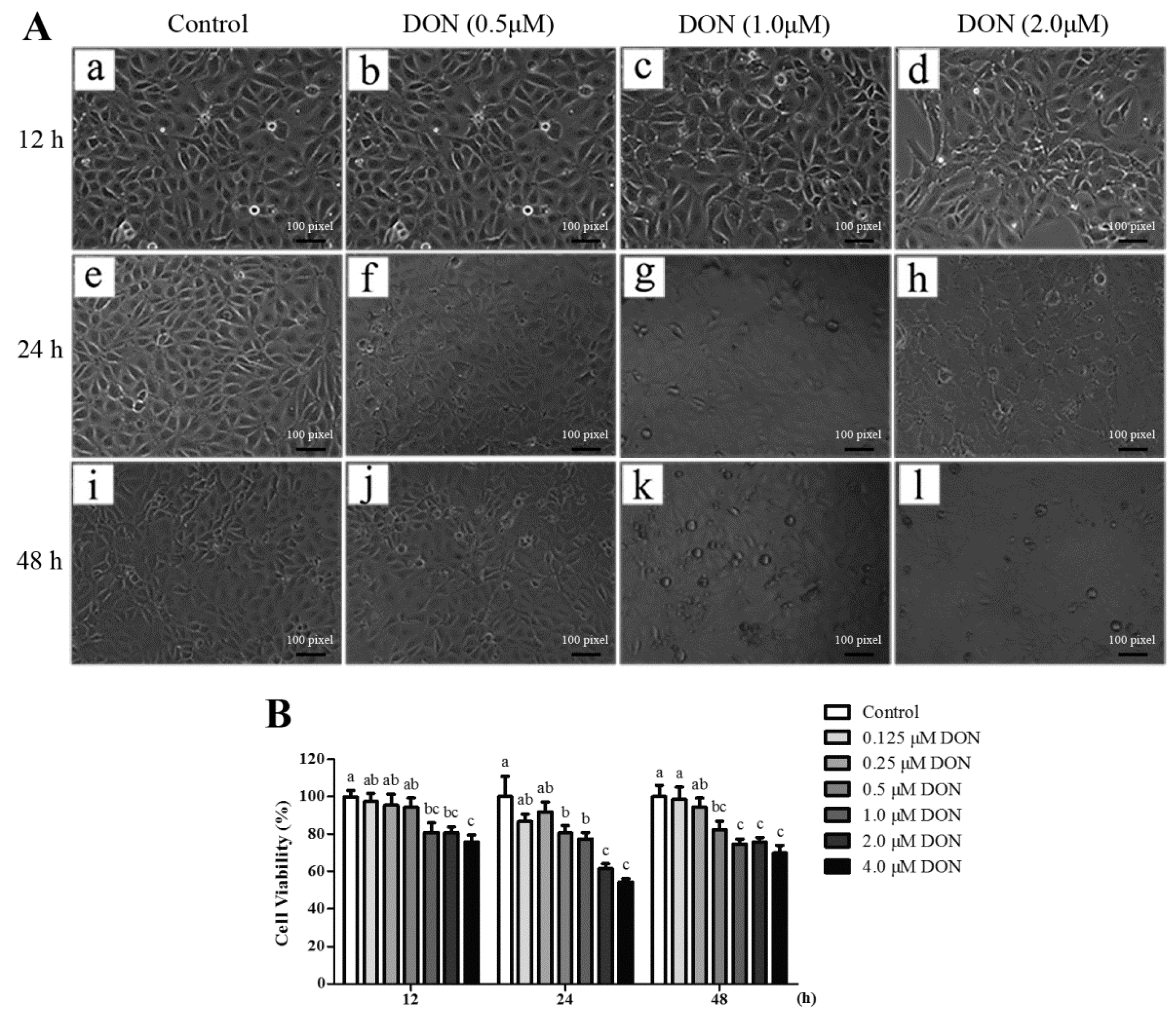
4.7. Cell Culture and Treatment
4.8. Quantitative PCR
4.9. Western Blotting
4.10. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Pinton, P.; Braicu, C.; Nougayrede, J.P.; Laffitte, J.; Taranu, I.; Oswald, I.P. Deoxynivalenol impairs porcine intestinal barrier function and decreases the protein expression of claudin-4 through a mitogen-activated protein kinase-dependent mechanism. J. Nutr. 2010, 140, 1956–1962. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Deoxynivalenol impairs weight gain and affects markers of gut health after low-dose, short-term exposure of growing pigs. Toxins 2015, 7, 2071–2095. [Google Scholar] [CrossRef] [PubMed]
- Wan, M.L.; Woo, C.S.; Allen, K.J.; Turner, P.C.; El-Nezami, H. Modulation of porcine β-defensins 1 and 2 upon individual and combined Fusarium toxin exposure in a swine jejunal epithelial cell line. Appl. Environ. Microbiol. 2013, 79, 2225–2232. [Google Scholar] [CrossRef] [PubMed]
- Fischer, N.; Sechet, E.; Friedman, R.; Amiot, A.; Sobhani, I.; Nigro, G.; Sansonetti, P.J.; Sperandio, B. Histone deacetylase inhibition enhances antimicrobial peptide but not inflammatory cytokine expression upon bacterial challenge. Proc. Natl. Acad. Sci. USA 2016, 113, E2993–E3001. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zeng, X.; Yang, Q.; Qiao, S. Antimicrobial peptides as potential alternatives to antibiotics in food animal industry. Int. J. Mol. Sci. 2016, 17, 603. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Gallo, R.L. AMPed up immunity: How antimicrobial peptides have multiple roles in immune defense. Trends Immunol. 2009, 30, 131–141. [Google Scholar] [CrossRef]
- Pasupuleti, M.; Schmidtchen, A.; Malmsten, M. Antimicrobial peptides: Key components of the innate immune system. Crit. Rev. Biotechnol. 2012, 32, 143–171. [Google Scholar] [CrossRef]
- Raqib, R.; Sarker, P.; Bergman, P.; Ara, G.; Lindh, M.; Sack, D.A.; Nasirul, I.K.; Gudmundsson, G.H.; Andersson, J.; Agerberth, B. Improved outcome in shigellosis associated with butyrate induction of an endogenous peptide antibiotic. Proc. Natl. Acad. Sci. USA 2006, 103, 9178–9183. [Google Scholar] [CrossRef] [Green Version]
- Aldhous, M.C.; Noble, C.L.; Satsangi, J. Dysregulation of human β-defensin-2 protein in inflammatory bowel disease. PLoS ONE 2009, 4, e6285. [Google Scholar] [CrossRef]
- Petnicki-Ocwieja, T.; Hrncir, T.; Liu, Y.J.; Biswas, A.; Hudcovic, T.; Tlaskalova-Hogenova, H.; Kobayashi, K.S. Nod2 is required for the regulation of commensal microbiota in the intestine. Proc. Natl. Acad. Sci. USA 2009, 106, 15813–15818. [Google Scholar] [CrossRef] [Green Version]
- Woo, J.I.; Oh, S.; Webster, P.; Lee, Y.J.; Lim, D.J.; Moon, S.K. NOD2/RICK-dependent β-defensin 2 regulation is protective for nontypeable Haemophilus influenzae-induced middle ear infection. PLoS ONE 2014, 9, e90933. [Google Scholar] [CrossRef]
- Tan, G.; Zeng, B.; Zhi, F.C. Regulation of human enteric α-defensins by NOD2 in the Paneth cell lineage. Eur. J. Cell Biol. 2015, 94, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Wehkamp, J.; Harder, J.; Weichenthal, M.; Schwab, M.; Schaffeler, E.; Schlee, M.; Herrlinger, K.R.; Stallmach, A.; Noack, F.; Fritz, P.; et al. NOD2 (CARD15) mutations in Crohn’s disease are associated with diminished mucosal α-defensin expression. Gut 2004, 53, 1658–1664. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, R.S.; Chang, K.C.; Swanson, P.E.; Tinsley, K.W.; Hui, J.J.; Klender, P.; Xanthoudakis, S.; Roy, S.; Black, C.; Grimm, E.; et al. Caspase inhibitors improve survival in sepsis: A critical role of the lymphocyte. Nat. Immunol. 2000, 1, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Zhu, H.; Morishima, N.; Li, E.; Xu, J.; Yankner, B.A.; Yuan, J. Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid-β. Nature 2000, 403, 98–103. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, P.M.; Yeretssian, G.; Rutherford, N.; Doiron, K.; Nadiri, A.; Zhu, L.; Green, D.R.; Gruenheid, S.; Saleh, M. Caspase-12 modulates NOD signaling and regulates antimicrobial peptide production and mucosal immunity. Cell Host Microbe 2008, 3, 146–157. [Google Scholar] [CrossRef]
- Saleh, M.; Mathison, J.C.; Wolinski, M.K.; Bensinger, S.J.; Fitzgerald, P.; Droin, N.; Ulevitch, R.J.; Green, D.R.; Nicholson, D.W. Enhanced bacterial clearance and sepsis resistance in caspase-12-deficient mice. Nature 2006, 440, 1064–1068. [Google Scholar] [CrossRef]
- Shi, Y.; Porter, K.; Parameswaran, N.; Bae, H.K.; Pestka, J.J. Role of GRP78/BiP degradation and ER stress in deoxynivalenol-induced interleukin-6 upregulation in the macrophage. Toxicol. Sci. 2009, 109, 247–255. [Google Scholar] [CrossRef]
- Park, S.H.; Do, K.H.; Choi, H.J.; Kim, J.; Kim, K.H.; Park, J.; Oh, C.G.; Moon, Y. Novel regulatory action of ribosomal inactivation on epithelial Nod2-linked proinflammatory signals in two convergent ATF3-associated pathways. J. Immunol. 2013, 191, 5170–5181. [Google Scholar] [CrossRef]
- Luk, G.D.; Bayless, T.M.; Baylin, S.B. Plasma postheparin diamine oxidase. Sensitive provocative test for quantitating length of acute intestinal mucosal injury in the rat. J. Clin. Investig. 1983, 71, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Navarrete, J.M.; Sabater, M.; Ortega, F.; Ricart, W.; Fernandez-Real, J.M. Circulating zonulin, a marker of intestinal permeability, is increased in association with obesity-associated insulin resistance. PLoS ONE 2012, 7, e37160. [Google Scholar] [CrossRef]
- Hancock, R.E.; Haney, E.F.; Gill, E.E. The immunology of host defence peptides: Beyond antimicrobial activity. Nat. Rev. Immunol. 2016, 16, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Taranu, I.; Marin, D.E.; Manda, G.; Motiu, M.; Neagoe, I.; Tabuc, C.; Stancu, M.; Olteanu, M. Assessment of the potential of a boron-fructose additive in counteracting the toxic effect of Fusarium mycotoxins. Br. J. Nutr. 2011, 106, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Rotter, B.A.; Thompson, B.K.; Lessard, M.; Trenholm, H.L.; Tryphonas, H. Influence of low-level exposure to Fusarium mycotoxins on selected immunological and hematological parameters in young swine. Fundam. Appl. Toxicol. 1994, 23, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Bergsjo, B.; Matre, T.; Nafstad, I. Effects of diets with graded levels of deoxynivalenol on performance in growing pigs. Zentralbl. Veterinarmed. A 1992, 39, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Rotter, B.A.; Prelusky, D.B.; Pestka, J.J. Toxicology of deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health 1996, 48, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Bracarense, A.P.; Lucioli, J.; Grenier, B.; Drociunas, P.G.; Moll, W.D.; Schatzmayr, G.; Oswald, I.P. Chronic ingestion of deoxynivalenol and fumonisin, alone or in interaction, induces morphological and immunological changes in the intestine of piglets. Br. J. Nutr. 2012, 107, 1776–1786. [Google Scholar] [CrossRef]
- Cheng, C.; Wei, H.; Xu, C.; Xie, X.; Jiang, S.; Peng, J. Maternal soluble fiber diet during pregnancy changes the intestinal microbiota, improves growth performance, and reduces intestinal permeability in piglets. Appl. Environ. Microbiol. 2018, 84, e01047. [Google Scholar] [CrossRef]
- Zhang, G.; Ross, C.R.; Blecha, F. Porcine antimicrobial peptides: New prospects for ancient molecules of host defense. Vet. Res. 2000, 31, 277–296. [Google Scholar] [CrossRef] [Green Version]
- Sang, Y.; Blecha, F. Porcine host defense peptides: Expanding repertoire and functions. Dev. Comp. Immunol. 2009, 33, 334–343. [Google Scholar] [CrossRef]
- Wang, S.; Thacker, P.A.; Watford, M.; Qiao, S. Functions of antimicrobial peptides in gut homeostasis. Curr. Protein Pept. Sci. 2015, 16, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.S.; Chamaillard, M.; Ogura, Y.; Henegariu, O.; Inohara, N.; Nunez, G.; Flavell, R.A. Nod2-dependent regulation of innate and adaptive immunity in the intestinal tract. Science 2005, 307, 731–734. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (NRC). Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Yu, H.T.; Ding, X.L.; Li, N.; Zhang, X.Y.; Zeng, X.F.; Wang, S.; Liu, H.B.; Wang, Y.M.; Jia, H.M.; Qiao, S.Y. Dietary supplemented antimicrobial peptide microcin J25 improves the growth performance, apparent total tract digestibility, fecal microbiota, and intestinal barrier function of weaned pigs. J. Anim. Sci. 2017, 95, 5064–5076. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists. Official Methods of Analysis; AOAC: Arlington, VA, USA, 2003. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Control | 1.28 mg/kg DON | 2.89 mg/kg DON | SEM | p-Value |
|---|---|---|---|---|---|
| Intitial body weight (kg) | 7.89 | 7.24 | 7.72 | 0.24 | 0.15 |
| Body weight at day 14 (kg) | 14.16 a | 12.11 b | 12.74 ab | 0.5 | 0.027 |
| Body weight at day 28 (kg) | 23.39 a | 20.43 b | 20.48 b | 0.67 | 0.007 |
| Average daily gain (g) | |||||
| 1–14 days | 447 | 347 | 358 | 29.51 | 0.051 |
| 15–28 days | 660 a | 594 ab | 553 b | 26.21 | 0.028 |
| 1–28 days | 554 a | 471 b | 456 b | 22.67 | 0.013 |
| Average daily feed intake (g/day) | |||||
| 1–14 days | 676 a | 552 b | 567 b | 34.95 | 0.041 |
| 15–28 days | 1217 a | 1072 b | 1027 b | 44.68 | 0.018 |
| 1–28 days | 946 a | 812 b | 798 b | 35.48 | 0.013 |
| Gain: Feed | |||||
| 1–14 days | 0.66 | 0.62 | 0.63 | 0.03 | 0.527 |
| 15–28 days | 0.55 | 0.55 | 0.54 | 0.02 | 0.8 |
| 1–28 days | 0.59 | 0.58 | 0.57 | 0.02 | 0.774 |
| Diarrhea score | 0.22 | 0.61 | 0.63 | - | - |
| Diarrhea incidence (%) | 9.38 | 25.45 | 29.02 | - | - |
| Item | 1–28 d |
|---|---|
| Ingredient (%) | |
| Maize | 62.64 |
| Soybean meal, dehulled (CP, 46%) | 13.80 |
| Concentrated soyabean protein | 5.00 |
| Fish meal (CP, 64%) | 2.50 |
| Whey powder | 10.00 |
| Soybean oil | 2.00 |
| Dicalcium phosphate | 1.00 |
| Limestone | 0.80 |
| Salt | 0.80 |
| L-Lys (78%) | 0.33 |
| DL-Met (98%) | 0.20 |
| Choline chloride | 0.13 |
| Zinc oxide | 0.30 |
| Mineral and vitamin premix 1 | 0.50 |
| Nutrient composition (% analysed) 2 | |
| Crude protein | 20.71 |
| Dry matter | 89.02 |
| Calcium | 0.85 |
| Phosphorus | 0.65 |
| Lys | 1.48 |
| Thr | 0.89 |
| Met + Cys | 0.92 |
| Calculated nutritional value | |
| Metabolic energy (kcal/kg) | 3250 |
| Gene | Forward Primer (5’–3’) | Reverse Primer (5’–3’) |
|---|---|---|
| β-actin | TACACCGCTACCAGTTCGC | GCTCGATGGGGTACTTGAGG |
| pBD1 | TGCCACAGGTGCCGATCT | TGCCACAGGTGCCGATCT |
| pBD2 | GACTGTCTGCCTCCTCTC | GGTCCCTTCAATCTGTTG |
| pBD3 | CCTTCTCTTTGCCTTGCTCTT | GCCACTCACAGAACAGCTACC |
| pEP2C | ACTGCTTGTTCTCCAGAGCC | TGGCACAGATGACAAAGCCT |
| PG1-5 | GTAGGTTCTGCGTCTGTGTCG | CAAATCCTTCACCGTCTACCA |
| PR39 | AGCAGTCCTCGGAAGCTAATC | GTCATTGGATGGGTTCAAGGT |
| PMAP23 | GGATTATAGACCTGCTGTGGA | AGAACTCTTCCCTGTGTCTTG |
| PMAP37 | GCTGTGTGACTTCAAGGAGAA | GAAATCTCCTGACACCCTCATT |
| NOD2 | GAGCGCATCCTCTTAACTTTCG | ACGCTCGTGATCCGTGAAC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Yang, J.; Zhang, B.; Wu, K.; Yang, A.; Li, C.; Zhang, J.; Zhang, C.; Rajput, S.A.; Zhang, N.; et al. Deoxynivalenol Impairs Porcine Intestinal Host Defense Peptide Expression in Weaned Piglets and IPEC-J2 Cells. Toxins 2018, 10, 541. https://doi.org/10.3390/toxins10120541
Wang S, Yang J, Zhang B, Wu K, Yang A, Li C, Zhang J, Zhang C, Rajput SA, Zhang N, et al. Deoxynivalenol Impairs Porcine Intestinal Host Defense Peptide Expression in Weaned Piglets and IPEC-J2 Cells. Toxins. 2018; 10(12):541. https://doi.org/10.3390/toxins10120541
Chicago/Turabian StyleWang, Shuai, Jiacheng Yang, Beiyu Zhang, Kuntan Wu, Ao Yang, Chong Li, Jiacai Zhang, Cong Zhang, Shahid Ali Rajput, Niya Zhang, and et al. 2018. "Deoxynivalenol Impairs Porcine Intestinal Host Defense Peptide Expression in Weaned Piglets and IPEC-J2 Cells" Toxins 10, no. 12: 541. https://doi.org/10.3390/toxins10120541
APA StyleWang, S., Yang, J., Zhang, B., Wu, K., Yang, A., Li, C., Zhang, J., Zhang, C., Rajput, S. A., Zhang, N., Sun, L., & Qi, D. (2018). Deoxynivalenol Impairs Porcine Intestinal Host Defense Peptide Expression in Weaned Piglets and IPEC-J2 Cells. Toxins, 10(12), 541. https://doi.org/10.3390/toxins10120541