Host and Cropping System Shape the Fusarium Population: 3ADON-Producers Are Ubiquitous in Wheat Whereas NIV-Producers Are More Prevalent in Rice


 ,
, 
Abstract
:1. Introduction
2. Results
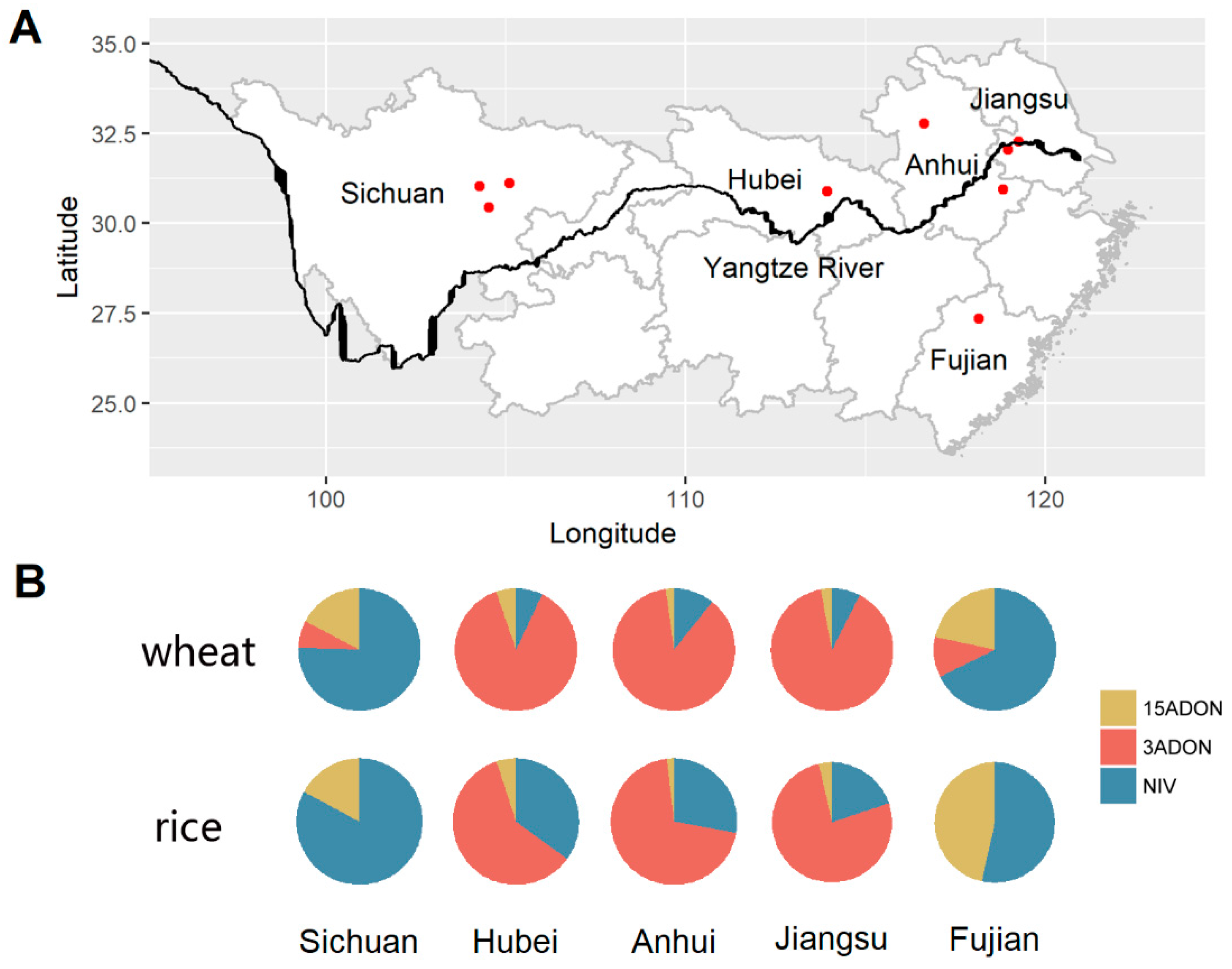
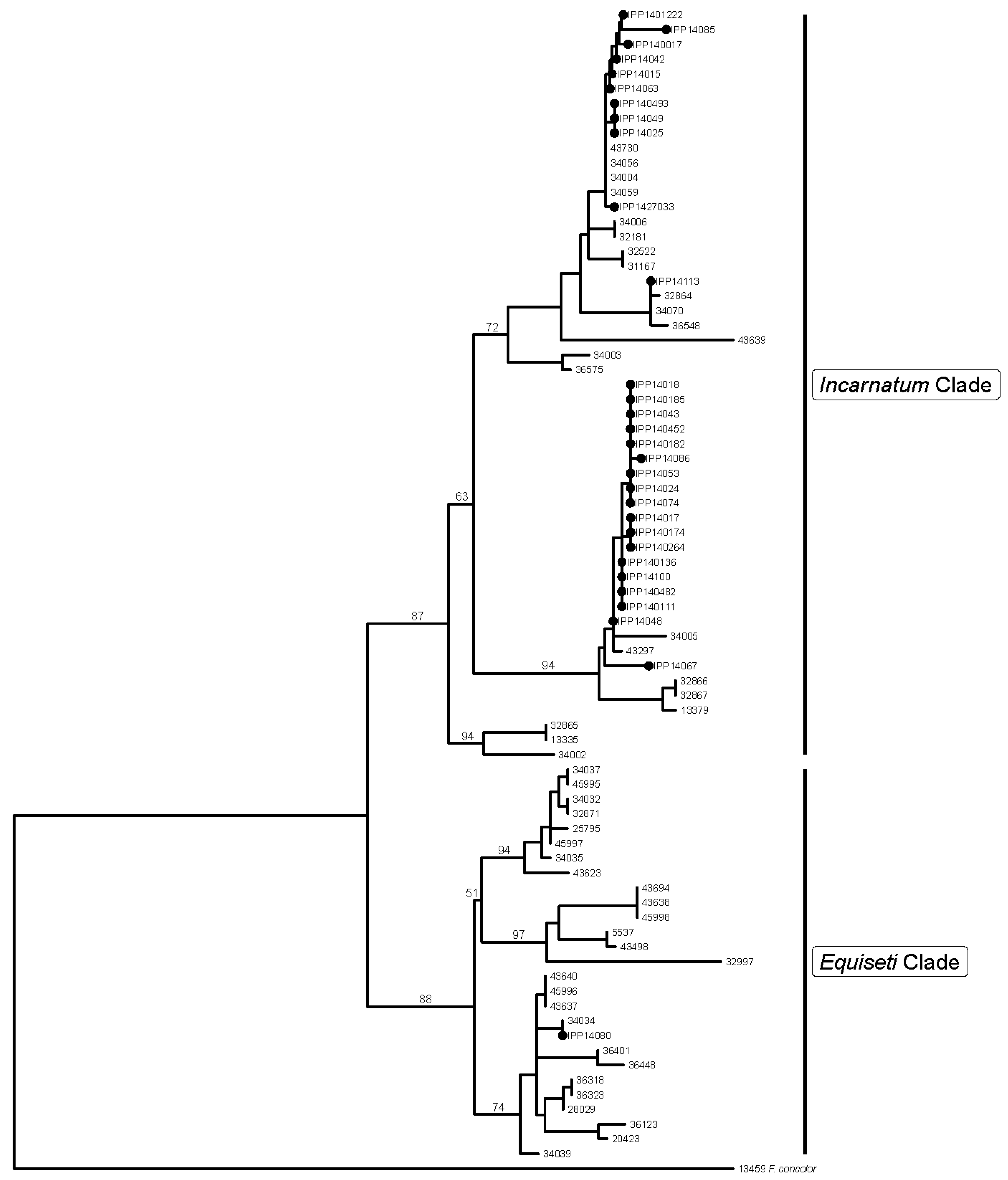
2.1. Fusarium Species Determination
2.2. Trichothecene Chemotype Identification
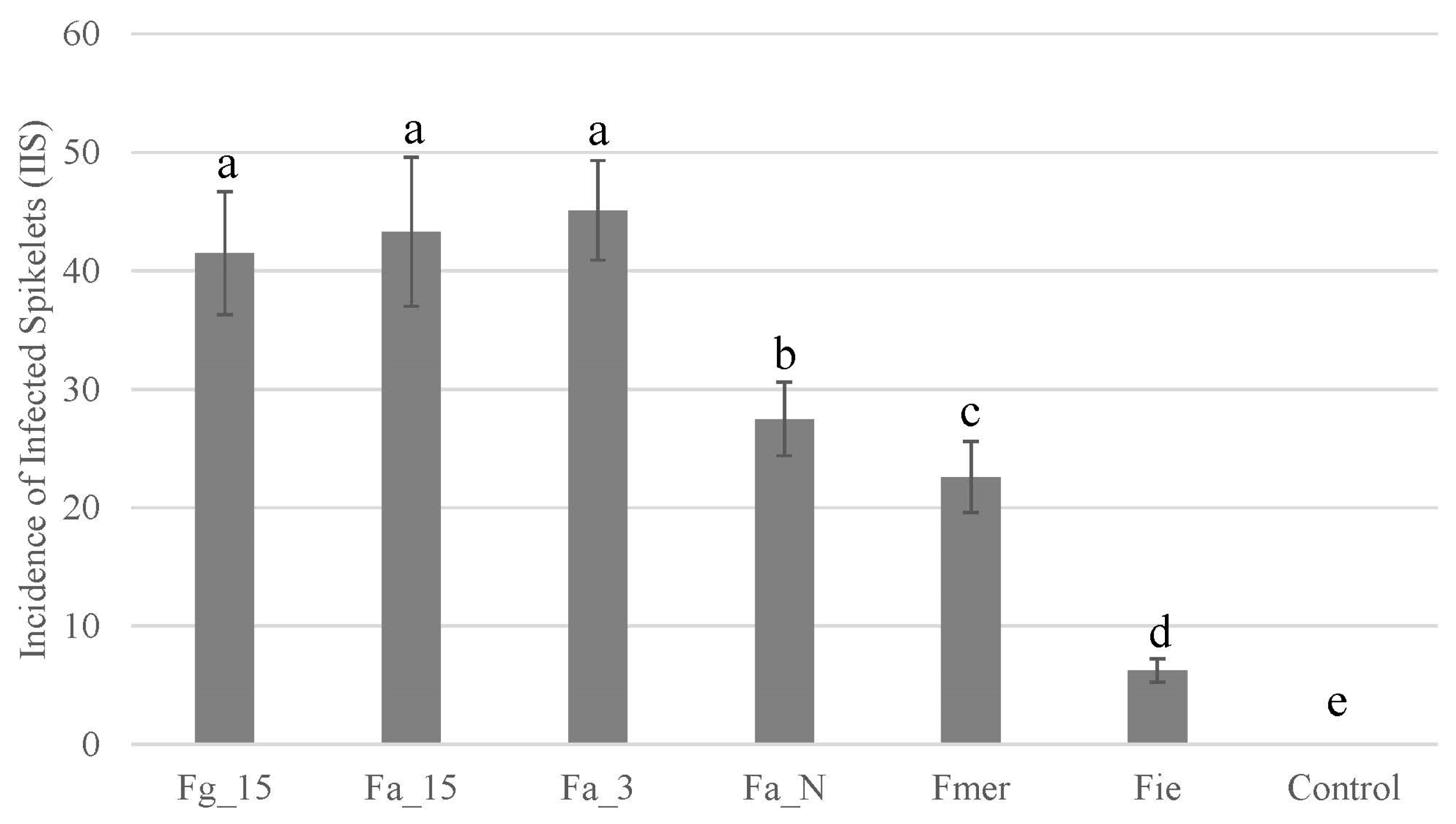
2.3. Pathogenicity Analyses
3. Discussion
4. Materials and Methods
4.1. Fungal Isolates
4.2. Genomic DNA Extraction
4.3. Species and Trichothecene Chemotype Determination
4.4. Pathogenicity Analyses
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kelly, A.C.; Clear, R.M.; O’Donnell, K.; McCormick, S.; Turkington, T.K.; Tekauz, A.; Gilbert, J.; Kistler, H.C.; Busman, M.; Ward, T.J. Diversity of Fusarium head blight populations and trichothecene toxin types reveals regional differences in pathogen composition and temporal dynamics. Fungal Genet. Biol. 2015, 82, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Van der Lee, T.; Waalwijk, C.; Chen, W.Q.; Xu, J.; Xu, J.S.; Zhang, Y.; Feng, J. Population analysis of the Fusarium graminearum species complex from wheat in China show a shift to more aggressive isolates. PLoS ONE 2012, 7, e31722. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Kistler, H.C. Heading for disaster: Fusarium graminearum on cereal crops. Mol. Plant Pathol. 2004, 5, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Sarver, B.A.; Ward, T.J.; Gale, L.R.; Broz, K.; Corby Kistler, H.; Aoki, T.; Nicholson, P.; Carter, J.; O’Donnell, K. Novel Fusarium head blight pathogens from Nepal and Louisiana revealed by multilocus genealogical concordance. Fungal Genet. Biol. 2011, 48, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Ward, T.J.; Aberra, D.; Kistler, H.C.; Aoki, T.; Orwig, N.; Kimura, M.; Bjornstad, S.; Klemsdal, S.S. Multilocus genotyping and molecular phylogenetics resolve a novel head blight pathogen within the Fusarium graminearum species complex from Ethiopia. Fungal Genet. Biol. 2008, 45, 1514–1522. [Google Scholar] [CrossRef] [PubMed]
- Suga, H.; Karugia, G.W.; Ward, T.; Gale, L.R.; Tomimura, K.; Nakajima, T.; Miyasaka, A.; Koizumi, S.; Kageyama, K.; Hyakumachi, M. Molecular characterization of the Fusarium graminearum species complex in Japan. Phytopathology 2008, 98, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, H.; Jeon, J.J.; Kim, H.S.; Zeller, K.A.; Carter, L.L.; Leslie, J.F.; Lee, Y.W. Population structure of and mycotoxin production by Fusarium graminearum from maize in South Korea. Appl. Environ. Microbiol. 2012, 78, 2161–2167. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D.; Greenhalgh, R.; Wang, Y.Z.; Lu, M. Trichothecene chemotypes of three Fusarium species. Mycologia 1991, 83, 121–130. [Google Scholar] [CrossRef]
- Kimura, M.; Kaneko, I.; Komiyama, M.; Takatsuki, A.; Koshino, H.; Yoneyama, K.; Yamaguchi, I. Trichothecene 3-O-acetyltransferase protects both the producing organism and transformed yeast from related mycotoxins. Cloning and characterization of Tri101. J. Biol. Chem. 1998, 273, 1654–1661. [Google Scholar] [CrossRef] [PubMed]
- Ward, T.J.; Clear, R.M.; Rooney, A.P.; O’Donnell, K.; Gaba, D.; Patrick, S.; Starkey, D.E.; Gilbert, J.; Geiser, D.M.; Nowicki, T.W. An adaptive evolutionary shift in Fusarium head blight pathogen populations is driving the rapid spread of more toxigenic Fusarium graminearum in North America. Fungal Genet. Biol. 2008, 45, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Umpierrez-Failache, M.; Garmendia, G.; Pereyra, S.; Rodriguez-Haralambides, A.; Ward, T.J.; Vero, S. Regional differences in species composition and toxigenic potential among Fusarium head blight isolates from Uruguay indicate a risk of nivalenol contamination in new wheat production areas. Int. J. Food Microbiol. 2013, 166, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Chang, I.Y.; Kim, H.; Yun, S.H.; Leslie, J.F.; Lee, Y.W. Genetic diversity and fitness of Fusarium graminearum populations from rice in Korea. Appl. Environ. Microbiol. 2009, 75, 3289–3295. [Google Scholar] [CrossRef] [PubMed]
- Gale, L.R.; Harrison, S.A.; Ward, T.J.; O’Donnell, K.; Milus, E.A.; Gale, S.W.; Kistler, H.C. Nivalenol-type populations of Fusarium graminearum and F. asiaticum are prevalent on wheat in southern Louisiana. Phytopathology 2011, 101, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Del Ponte, E.M.; Spolti, P.; Ward, T.J.; Gomes, L.B.; Nicolli, C.P.; Kuhnem, P.R.; Silva, C.N.; Tessmann, D.J. Regional and field-specific factors affect the composition of Fusarium head blight pathogens in subtropicalno-till wheat agroecosystem of Brazil. Phytopathology 2015, 105, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Gomes, L.B.; Ward, T.J.; Badiale-Furlong, E.; Del Ponte, E.M. Species composition, toxigenic potential and pathogenicity of Fusarium graminearum species complex isolates from southern Brazilian rice. Plant Pathol. 2015, 64, 980–987. [Google Scholar] [CrossRef]
- Kuhnem, P.R.; Ward, T.J.; Silva, C.N.; Spolti, P.; Ciliato, M.L.; Tessmann, D.J.; Del Ponte, E.M. Composition and toxigenic potential of the Fusarium graminearum species complex from maize ears, stalks and stubble in Brazil. Plant Pathol. 2016, 65, 1185–1191. [Google Scholar] [CrossRef]
- Beukes, I.; Rose, L.J.; van Coller, G.J.; Viljoen, A. Disease development and mycotoxin production by the Fusarium graminearum species complex associated with South African maize and wheat. Eur. J. Plant Pathol. 2018, 150, 893–910. [Google Scholar] [CrossRef]
- Boutigny, A.-L.; Ward, T.J.; Coller, G.J.V.; Flett, B.; Lamprecht, S.C.; O’Donnell, K.; Viljoen, A. Analysis of the Fusarium graminearum species complex from wheat, barley and maize in South Africa provides evidence of species-specific differences in host preference. Fungal Genet. Biol. 2011, 48, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Brankovics, B.; Luo, W.; Xu, J.; Xu, J.S.; Guo, C.; Guo, J.G.; Jin, S.L.; Chen, W.Q.; Feng, J. Crops are a main driver for species diversity and the toxigenic potential of Fusarium isolates in maize ears in China. World Mycotoxin J. 2016, 9, 701–715. [Google Scholar] [CrossRef]
- Zhang, J.B.; Wang, J.H.; Gong, A.D.; Chen, F.F.; Song, B.; Li, X.; Li, H.P.; Peng, C.H.; Liao, Y.C. Natural occurrence of fusarium head blight, mycotoxins and mycotoxin-producing isolates of Fusarium in commercial fields of wheat in Hubei. Plant Pathol. 2013, 62, 92–102. [Google Scholar] [CrossRef]
- Zhang, J.B.; Li, H.P.; Dang, F.J.; Qu, B.; Xu, Y.B.; Zhao, C.S.; Liao, Y.C. Determination of the trichothecene mycotoxin chemotypes and associated geographical distribution and phylogenetic species of the Fusarium graminearum clade from China. Mycol. Res. 2007, 111, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.B.; Xu, J.H.; Shi, J.R. Molecular characterization of the Fusarium graminearum species complex in Eastern China. Eur. J. Plant Pathol. 2014, 139, 811–823. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, Z.; van der Lee, T.; Chen, W.Q.; Xu, J.; Xu, J.S.; Yang, L.; Yu, D.; Waalwijk, C.; Feng, J. Population genetic analyses of Fusarium asiaticum populations from barley suggest a recent shift favoring 3ADON producers in southern China. Phytopathology 2010, 100, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Puri, K.D.; Saucedo, E.S.; Zhong, S. Molecular characterization of Fusarium head blight pathogens sampled from a naturally infected disease nursery used for wheat breeding programs in China. Plant Dis. 2012, 96, 1280–1285. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Proctor, R.H. Genetic diversity and trichothecene chemotypes of the Fusarium graminearum clade isolated from maize in Nepal and identification of a putative new lineage. Fungal Genet. Biol. 2011, 115, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Sampietro, D.A.; Ficoseco, M.E.; Jimenez, C.M.; Vattuone, M.A.; Catalan, C.A. Trichothecene genotypes and chemotypes in Fusarium graminearum complex strains isolated from maize fields of northwest Argentina. Int. J. Food Microbiol. 2012, 153, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.; Summerell, B. The Fusarium Laboratory Manual; Blackwell Publishing Ltd.: Oxford, UK, 2006. [Google Scholar]
- Van der Lee, T.; Zhang, H.; van Diepeningen, A.; Waalwijk, C. Biogeography of Fusarium graminearum species complex and chemotypes: A review. Food Addit. Contam. 2015, 32, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Geiser, D.M.; del Mar Jiménez-Gasco, M.; Kang, S.; Makalowska, I.; Veeraraghavan, N.; Ward, T.J.; Zhang, N.; Kuldau, A.G.; O’Donnell, K. FUSARIUM-ID v. 1.0: A DNA sequence database for identifying Fusarium. Eur. J. Plant Pathol. 2004, 110, 473–479. [Google Scholar] [CrossRef]
- Fusarium MLST Database. Available online: http://www.westerdijkinstitute.nl/fusarium/ (accessed on 7 March 2018).




{kind=link}
{kind=link}
{kind=link}
| Species Complex * | Provinces | Sichuan | Hubei | Anhui | Jiangsu | Fujian | Total | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sampling Sites | Jianyang | Guanghan | Mianyang | Xiaogan | Fengtai | Xuanzhou | Nanjing | Yizheng | Nanping | |||||||||||
| Hosts ** | W | R | W | R | W | R | W | R | W | R | W | R | W | R | W | R | W | R | ||
| FGSC | F. asiaticum | 17 | 56 | 38 | 49 | 28 | 74 | 57 | 60 | 42 | 28 | 50 | 81 | 28 | 75 | 37 | 65 | 211 | 114 | 1110 |
| F. graminearum | 4 | 21 | 1 | 1 | 14 | 14 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 2 | 0 | 6 | 0 | 67 | |
| F. meridionale | 1 | 5 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9 | |
| FIESC | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 21 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 1 | 3 | 2 | 30 | |
| FTSC | F. acuminatum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| F. tricinctum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| FSPSC | F. armeniacum | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 6 |
| FSAMSC | F. kyushuense | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 |
| FFSC | F. fujikuroi | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 6 | 0 | 0 | 7 |
| F. proliferatum | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 6 | |
| F. concentricum | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | |
| F. nygamai | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 3 | |
| F. verticillioides | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| Total | 22 | 87 | 39 | 52 | 43 | 90 | 57 | 84 | 43 | 29 | 51 | 90 | 29 | 78 | 39 | 76 | 221 | 116 | 1246 | |
| Hosts | Provinces | F. asiaticum | F. graminearum | F. meridionale | Total | ||
|---|---|---|---|---|---|---|---|
| NIV | 3ADON | 15ADON | 15ADON | NIV | |||
| Wheat | Sichuan | 74 | 8 | 1 | 19 | 1 | 103 |
| Hubei | 4 | 50 | 3 | 0 | 0 | 57 | |
| Anhui | 10 | 81 | 1 | 1 | 0 | 93 | |
| Jiangsu | 5 | 60 | 0 | 2 | 0 | 67 | |
| Fujian | 147 | 23 | 41 | 6 | 0 | 217 | |
| Rice | Sichuan | 179 | 0 | 0 | 36 | 8 | 223 |
| Hubei | 21 | 36 | 3 | 0 | 0 | 60 | |
| Anhui | 31 | 78 | 0 | 2 | 0 | 111 | |
| Jiangsu | 28 | 108 | 4 | 1 | 0 | 141 | |
| Fujian | 61 | 0 | 53 | 0 | 0 | 114 | |
| Total | 560 | 444 | 106 | 67 | 9 | 1186 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Zhang, H.; Kong, X.; Van der Lee, T.; Waalwijk, C.; Van Diepeningen, A.; Xu, J.; Xu, J.; Chen, W.; Feng, J. Host and Cropping System Shape the Fusarium Population: 3ADON-Producers Are Ubiquitous in Wheat Whereas NIV-Producers Are More Prevalent in Rice. Toxins 2018, 10, 115. https://doi.org/10.3390/toxins10030115
Yang M, Zhang H, Kong X, Van der Lee T, Waalwijk C, Van Diepeningen A, Xu J, Xu J, Chen W, Feng J. Host and Cropping System Shape the Fusarium Population: 3ADON-Producers Are Ubiquitous in Wheat Whereas NIV-Producers Are More Prevalent in Rice. Toxins. 2018; 10(3):115. https://doi.org/10.3390/toxins10030115
Chicago/Turabian StyleYang, Meixin, Hao Zhang, Xiangjiu Kong, Theo Van der Lee, Cees Waalwijk, Anne Van Diepeningen, Jin Xu, Jingsheng Xu, Wanquan Chen, and Jie Feng. 2018. "Host and Cropping System Shape the Fusarium Population: 3ADON-Producers Are Ubiquitous in Wheat Whereas NIV-Producers Are More Prevalent in Rice" Toxins 10, no. 3: 115. https://doi.org/10.3390/toxins10030115
APA StyleYang, M., Zhang, H., Kong, X., Van der Lee, T., Waalwijk, C., Van Diepeningen, A., Xu, J., Xu, J., Chen, W., & Feng, J. (2018). Host and Cropping System Shape the Fusarium Population: 3ADON-Producers Are Ubiquitous in Wheat Whereas NIV-Producers Are More Prevalent in Rice. Toxins, 10(3), 115. https://doi.org/10.3390/toxins10030115