The Primary Duct of Bothrops jararaca Glandular Apparatus Secretes Toxins

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
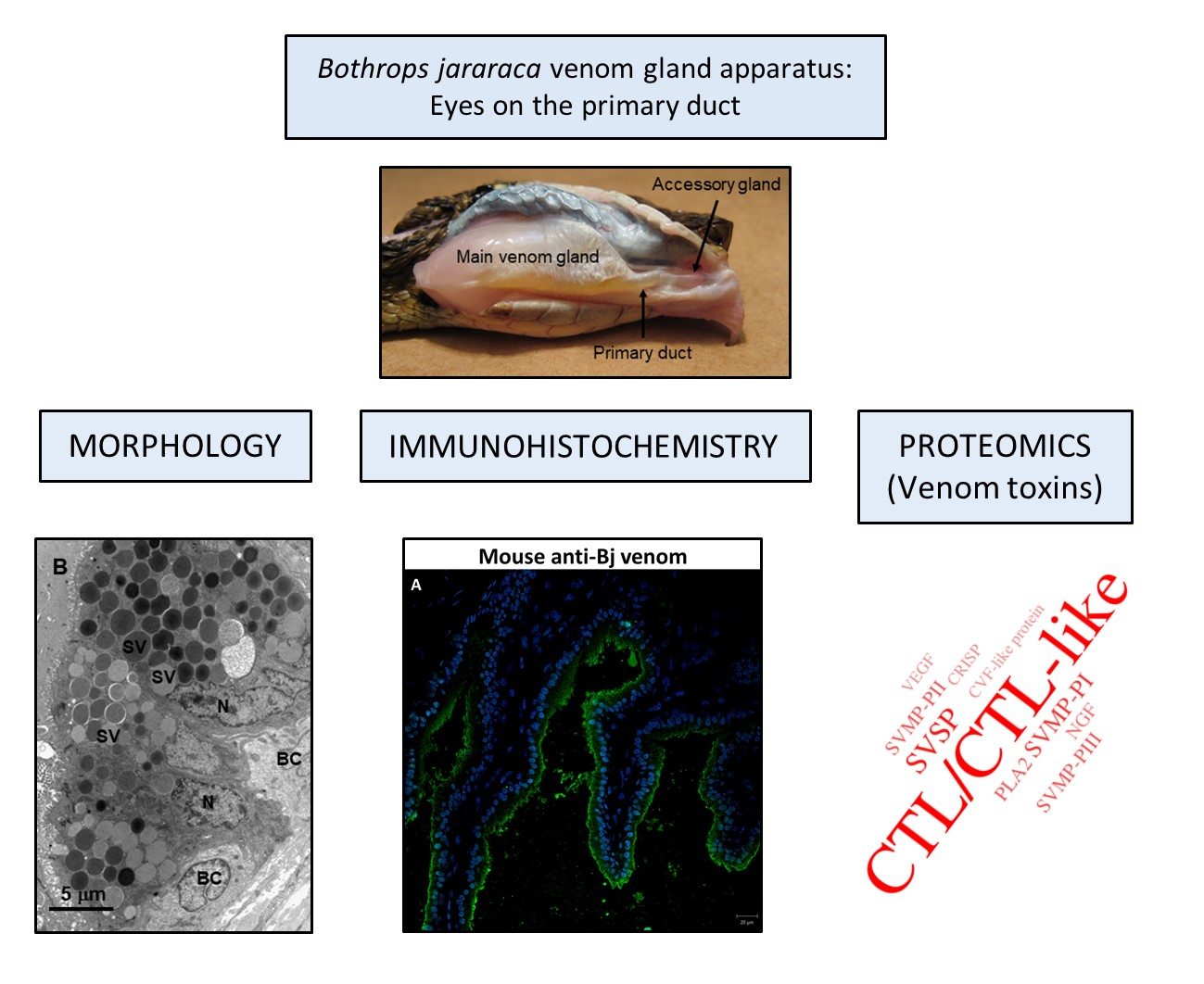
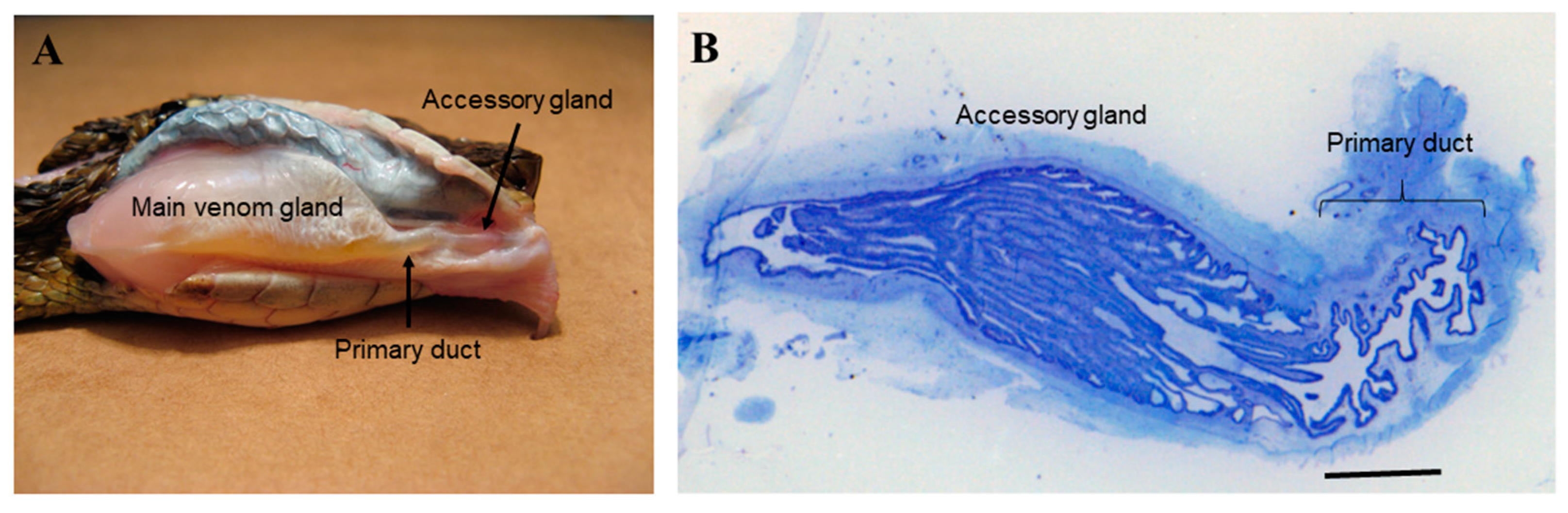
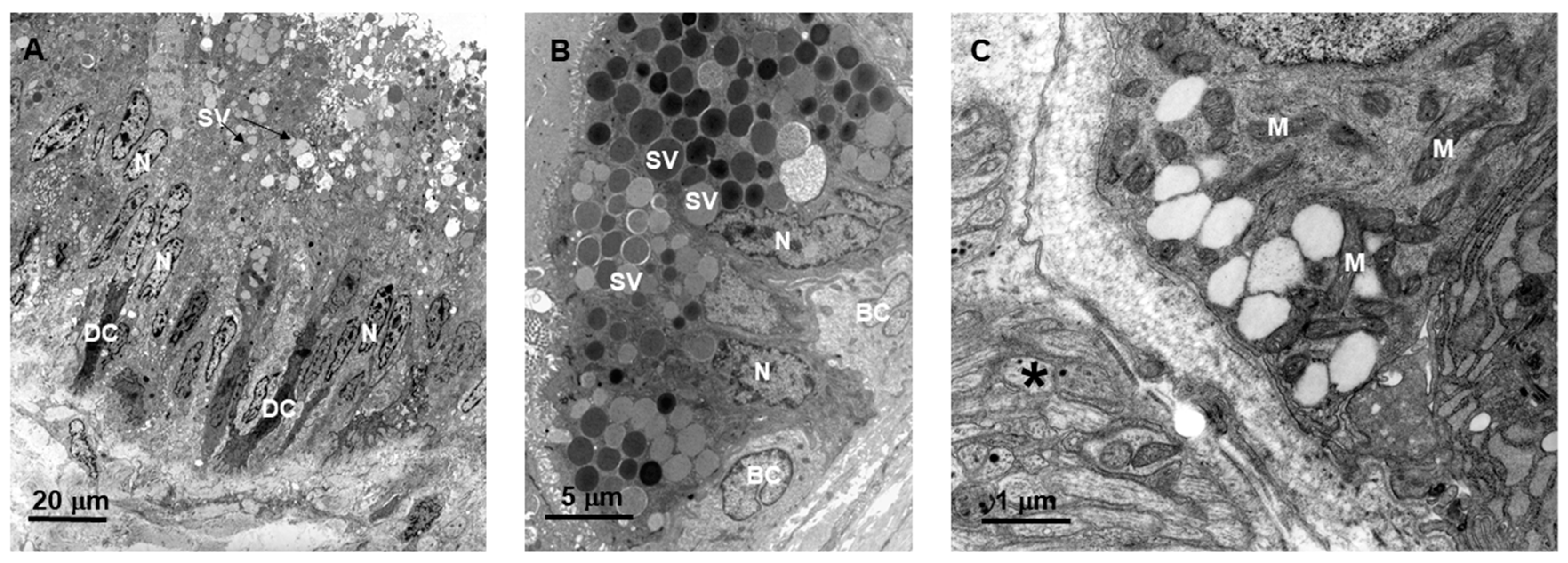
2.1. Bothrops jararaca Snake Primary Duct’s Morphology
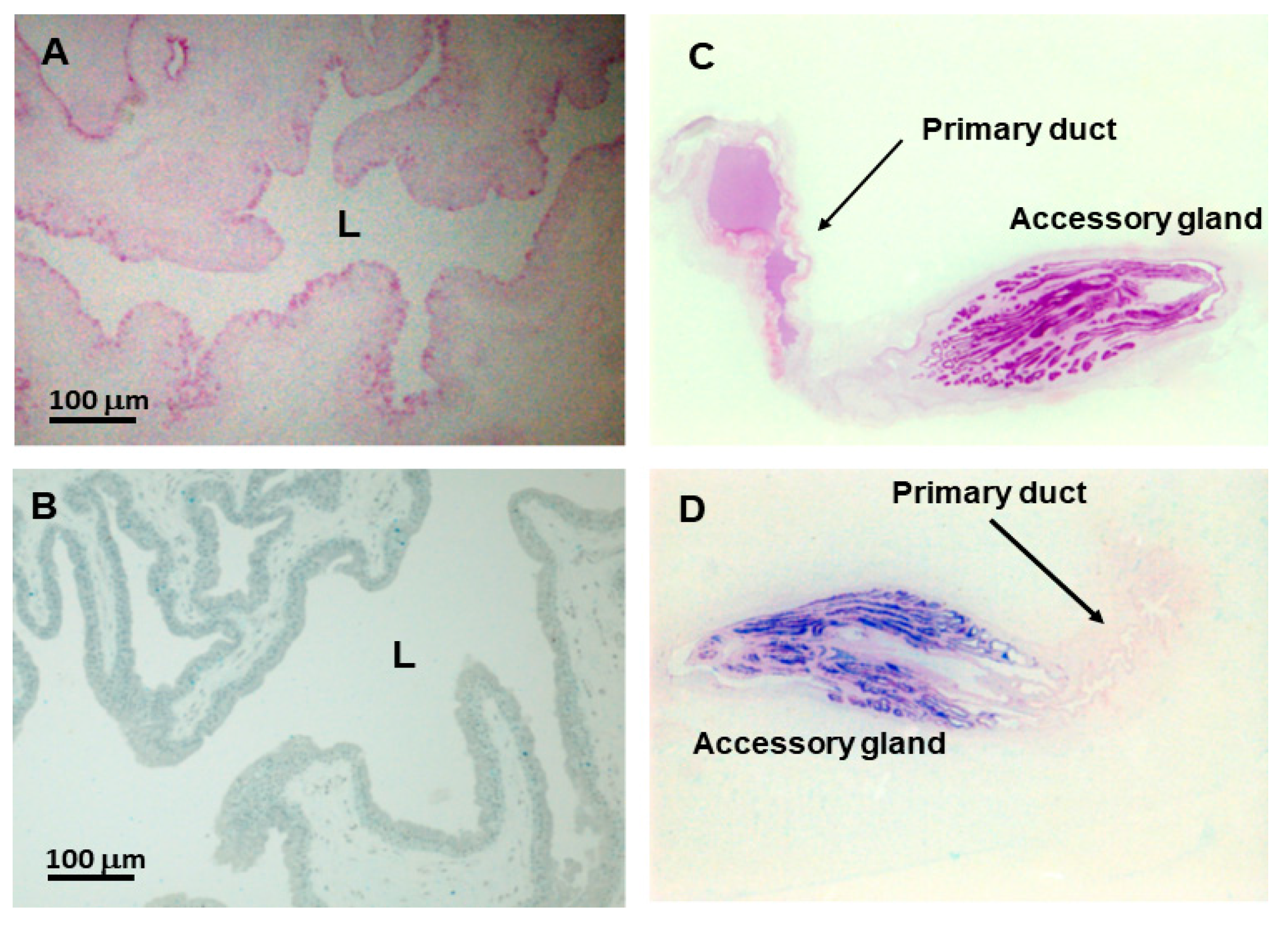
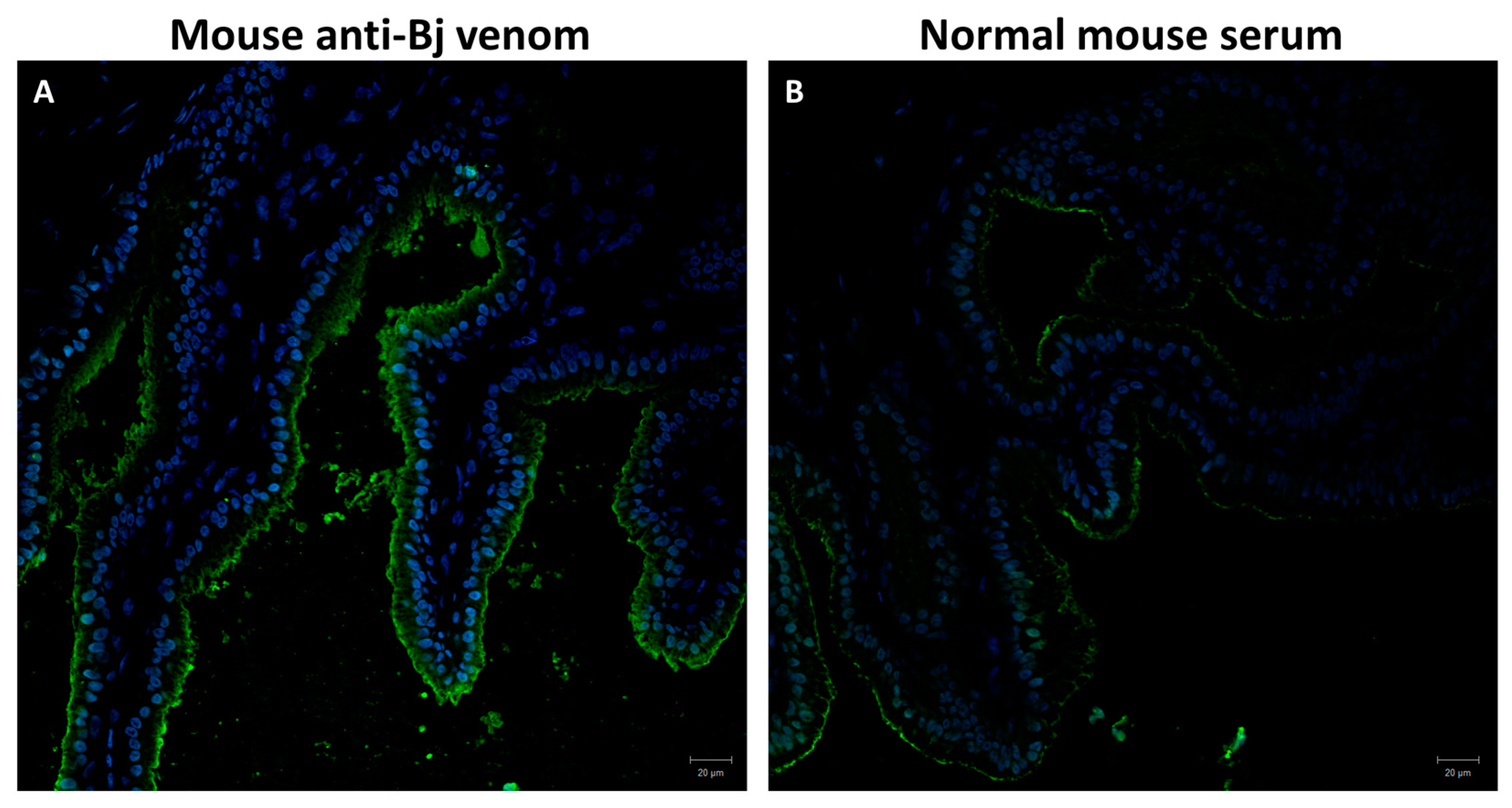
2.2. Detection of Venom in the Primary Duct
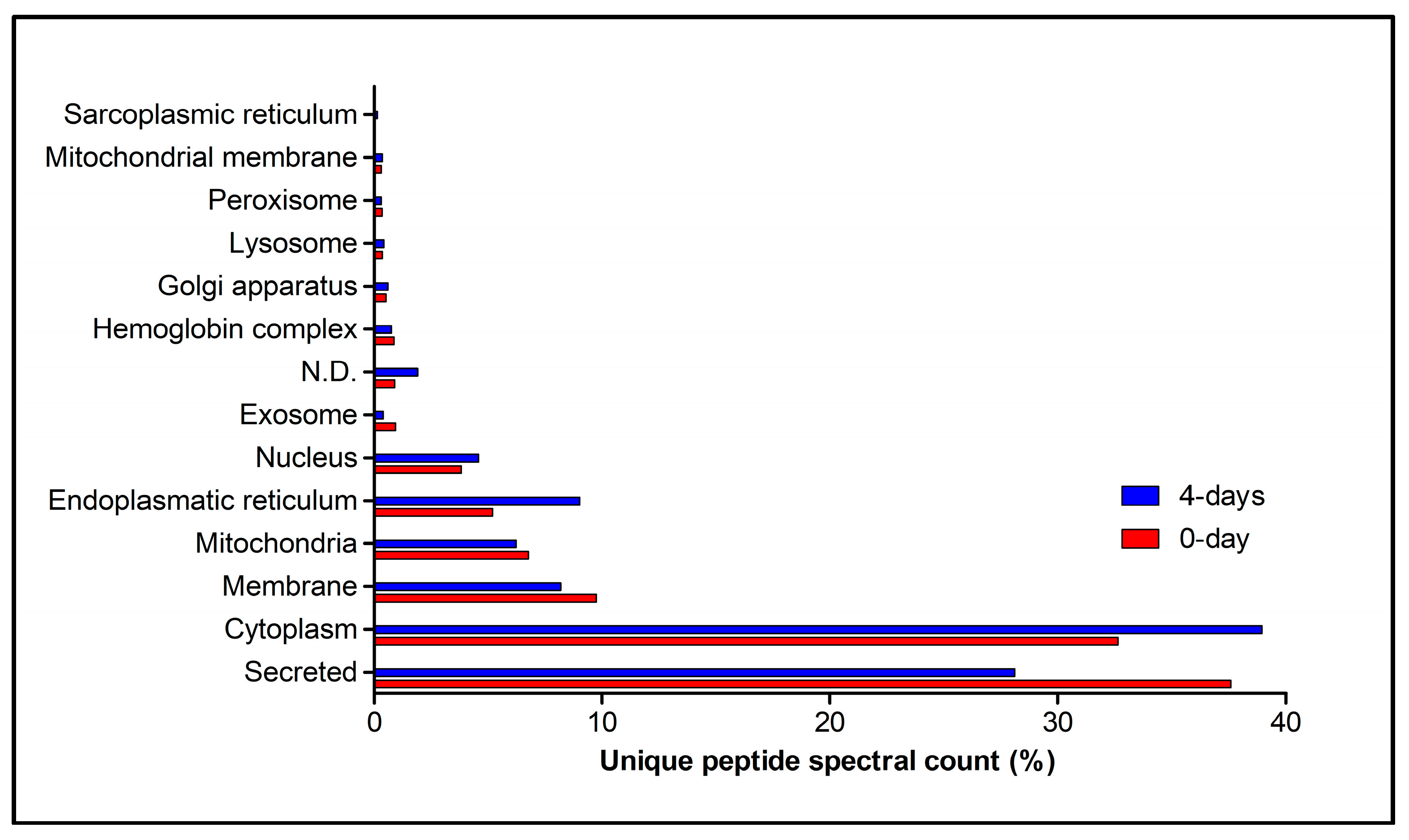
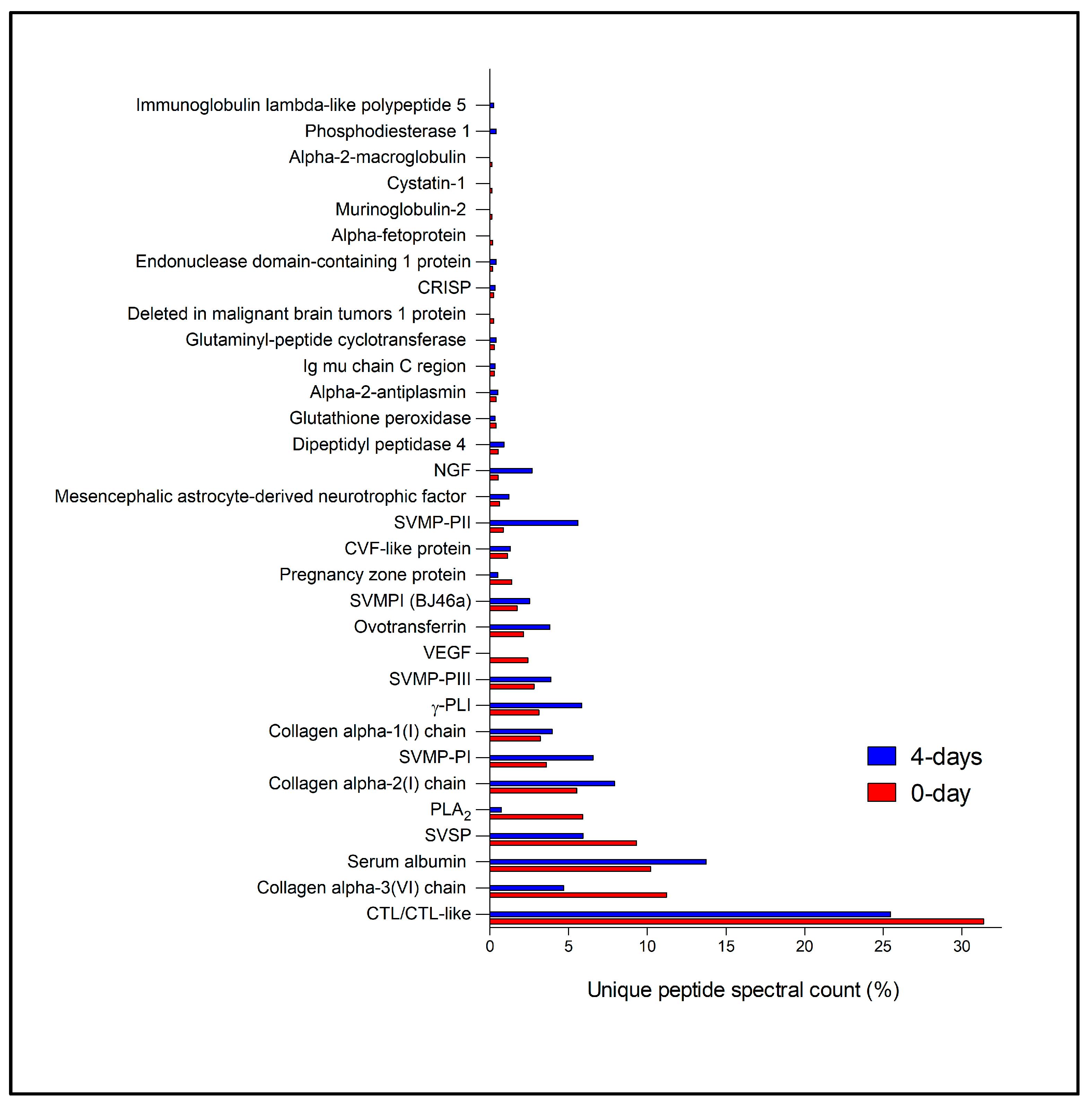
2.3. Primary Duct’s Secretome Proteomic Analysis
3. Conclusions
4. Materials and Methods
4.1. Animals and Primary Duct
4.2. Light and Electron Microscopy
4.3. Immunohistochemistry Analysis
4.4. Proteomics
4.4.1. Protein Extraction
4.4.2. SDS-PAGE (Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis)
4.4.3. In-Gel Trypsin Digestion
4.4.4. Mass Spectrometric Analysis
4.4.5. Data Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gomes, N.; Puorto, G. Atlas anatômico de Bothrops jararaca Wied, 1824 (Serpentes: Viperidae). Mem. Inst. Butantan 1993, 55, 69–100. [Google Scholar]
- Kochva, E.; Gans, C. Histology and histochemistry of venom glands of some Crotaline snakes. Copeia 1966, 3, 506–515. [Google Scholar] [CrossRef]
- Kochva, E.; Gans, C. The venom gland of Vipera palestinae with comments on the glands of some other viperines. Acta Anat. 1965, 62, 365–401. [Google Scholar] [CrossRef]
- Mackessy, S.P.; Baxter, L.M. Bioweapons synthesis and storage: The venom gland of front-fanged snakes. Zool. Anz. 2006, 245, 147–159. [Google Scholar] [CrossRef]
- Ben-Shaul, Y.; Lifshitz, S.H.; Kochva, E. Ultrastructural aspects of secretion in the venom glands of Vipera palaestinae. In Toxins of Animal and Plant Origin; de Vries, A., Kochva, E., Eds.; Gordon and Breach: London, UK, 1971; pp. 87–107. [Google Scholar]
- Carneiro, S.M.; Pinto, V.R.; Jared, C.; Lula, L.A.; Faria, F.P.; Sesso, A. Morphometric studies on venom secretory cells from Bothrops jararacussu (Jararacuçu) before and after venom extraction. Toxicon 1991, 29, 569–580. [Google Scholar] [CrossRef]
- De Lucca, F.L.; Haddad, A.; Kochva, E.; Rothschild, A.M.; Valeri, V. Protein synthesis and morphological changes in the secretory epithelium of the venom gland of Crotalus durissus terrificus at different times after manual extraction of venom. Toxicon 1974, 12, 361–368. [Google Scholar] [CrossRef]
- Kochva, E. The origin of snakes and evolution of the venom apparatus. Toxicon 1987, 25, 65–106. [Google Scholar] [CrossRef]
- Mackessy, S.P. Morphology and ultrastructure of the venom gland of the Northern pacific rattlesnake Crotalus viridis oreganus. J. Morphol. 1991, 208, 109–128. [Google Scholar] [CrossRef]
- Rotenberg, D.; Bamberger, E.S.; Kochva, E. Studies on ribonucleic acid synthesis in the venom glands of Vipera palaestinae (Ophidia, Reptilia). Biochem. J. 1971, 121, 609–612. [Google Scholar] [CrossRef] [PubMed]
- Luna, M.S.; Hortencio, T.M.; Ferreira, Z.S.; Yamanouye, N. Sympathetic outflow activates the venom gland of the snake Bothrops jararaca by regulating the activation of transcription factors and the synthesis of venom gland proteins. J. Exp. Biol. 2009, 212, 1535–1543. [Google Scholar] [CrossRef] [PubMed]
- Kerchove, C.M.; Carneiro, S.M.; Markus, R.P.; Yamanouye, N. Stimulation of the alpha-adrenoceptor triggers the venom production cycle in the venom gland of Bothrops jararaca. J. Exp. Biol. 2004, 207, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Kerchove, C.M.; Luna, M.S.; Zablith, M.B.; Lazari, M.F.; Smaili, S.S.; Yamanouye, N. Alpha1-adrenoceptors trigger the snake venom production cycle in secretory cells by activating phosphatidylinositol 4,5-bisphosphate hydrolysis and ERK signaling pathway. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 150, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Yamanouye, N.; Britto, L.R.; Carneiro, S.M.; Markus, R.P. Control of venom production and secretion by sympathetic outflow in the snake Bothrops jararaca. J. Exp. Biol. 1997, 200, 2547–2556. [Google Scholar] [PubMed]
- Yamanouye, N.; Carneiro, S.M.; Scrivano, C.N.; Markus, R.P. Characterization of beta-adrenoceptors responsible for venom production in the venom gland of the snake Bothrops jararaca. Life Sci. 2000, 67, 217–226. [Google Scholar] [CrossRef]
- Gans, C.; Kochva, E. The accessory gland in the venom apparatus of viperid snakes. Toxicon 1965, 3, 61–63. [Google Scholar] [CrossRef]
- Gennaro, J.F., Jr.; Callahan, W.P., 3rd; Lorincz, A.E. The anatomy and biochemistry of a mucus-secreting cell type present in the poison apparatus of the pit viper Ancistrodon piscivorus piscivorus. Ann. N. Y. Acad. Sci. 1963, 106, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Gennaro, J.F.; Hall, H.P.; Casey, E.R.; Hayes, W.K. Neurotropic effects of venoms and other factors that promote prey acquisition. J. Exp. Zool. A Ecol. Genet. Physiol. 2007, 307, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, R.; Lorincz, A.E.; Gennaro, J.F., Jr. Polysaccharide content of the poison apparatus of the cottonmouth moccasin Agkistrodon piscivorus piscivorus. Toxicon 1967, 5, 125–131. [Google Scholar] [CrossRef]
- Sakai, F.; Carneiro, S.M.; Yamanouye, N. Morphological study of accessory gland of Bothrops jararaca and its secretory cycle. Toxicon 2012, 59, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Valente, R.H.; Luna, M.S.; de Oliveira, U.C.; Nishiyama, M.Y., Jr.; Junqueira de Azevedo Ide, L.; Portes-Junior, J.A.; Clissa, P.B.; Viana, L.G.; Sanches, L.; Moura da Silva, A.M.; et al. Bothrops jararaca accessory venom gland is an ancillary source of toxins to the snake. J. Proteom. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Young, B.A.; Blair, M.; Zahn, K.; Marvin, J. Mechanics of venom expulsion in Crotalus, with special reference to the role of the fang sheath. Anat. Rec. 2001, 264, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Nicolau, C.A.; Carvalho, P.C.; Junqueira-de-Azevedo, I.L.; Teixeira-Ferreira, A.; Junqueira, M.; Perales, J.; Neves-Ferreira, A.G.; Valente, R.H. An in-depth snake venom proteopeptidome characterization: Benchmarking Bothrops jararaca. J. Proteom. 2017, 151, 214–231. [Google Scholar] [CrossRef] [PubMed]
- Luna, M.S.; Valente, R.H.; Perales, J.; Vieira, M.L.; Yamanouye, N. Activation of Bothrops jararaca snake venom gland and venom production: A proteomic approach. J. Proteom. 2013, 94, 460–472. [Google Scholar] [CrossRef] [PubMed]
- Kardong, K. The evolution of the venom apparatus in snakes from colubrids to viperids & elapids. Mem. Inst. Butantan 1982, 46, 106–118. [Google Scholar]
- Breno, M.C.; Yamanouye, N.; Prezoto, B.C.; Lazari, M.F.M.; Toffoletto, O.; Picarelli, Z.P. Maintenance of the snake Bothrops jararaca (Wied, 1824) in captivity. Snake 1990, 22, 126–130. [Google Scholar]
- Secor, S.M.; Nagy, K. Bioenergetic correlates of foraging mode fo the snakes Crotalus cerastes and Masticophis flagellum. Ecology 1994, 75, 1600–1614. [Google Scholar] [CrossRef]
- Yamanouye, N.; Kerchove, C.M.; Moura-da-Silva, A.M.; Carneiro, S.M.; Markus, R.P. Long-term primary culture of secretory cells of Bothrops jararaca venom gland for venom production in vitro. Nat. Protoc. 2006, 1, 2763–2766. [Google Scholar] [CrossRef] [PubMed]
- Belloumini, H.E. Extraction and quantities of venom obtained from some Brazilian snakes. In Venomous Animals and Their Venoms; Buherl, W., Buckley, V., Deulofeu, V., Eds.; Academic Press: New York, NY, USA, 1968; pp. 97–117. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hemmi Valente, R.; Sakai, F.; Portes-Junior, J.A.; Godoy Viana, L.; Mendes Carneiro, S.; Perales, J.; Yamanouye, N. The Primary Duct of Bothrops jararaca Glandular Apparatus Secretes Toxins. Toxins 2018, 10, 121. https://doi.org/10.3390/toxins10030121
Hemmi Valente R, Sakai F, Portes-Junior JA, Godoy Viana L, Mendes Carneiro S, Perales J, Yamanouye N. The Primary Duct of Bothrops jararaca Glandular Apparatus Secretes Toxins. Toxins. 2018; 10(3):121. https://doi.org/10.3390/toxins10030121
Chicago/Turabian StyleHemmi Valente, Richard, Fernanda Sakai, José Antonio Portes-Junior, Luciana Godoy Viana, Sylvia Mendes Carneiro, Jonas Perales, and Norma Yamanouye. 2018. "The Primary Duct of Bothrops jararaca Glandular Apparatus Secretes Toxins" Toxins 10, no. 3: 121. https://doi.org/10.3390/toxins10030121
APA StyleHemmi Valente, R., Sakai, F., Portes-Junior, J. A., Godoy Viana, L., Mendes Carneiro, S., Perales, J., & Yamanouye, N. (2018). The Primary Duct of Bothrops jararaca Glandular Apparatus Secretes Toxins. Toxins, 10(3), 121. https://doi.org/10.3390/toxins10030121