Biological Activities of Cationicity-Enhanced and Hydrophobicity-Optimized Analogues of an Antimicrobial Peptide, Dermaseptin-PS3, from the Skin Secretion of Phyllomedusa sauvagii

 ,
, _Kwok.png) and
and 
Abstract
:1. Introduction
2. Results
2.1. Molecular Cloning of a DPS1 Precursor cDNA from a Skin Secretion-Derived cDNA Library
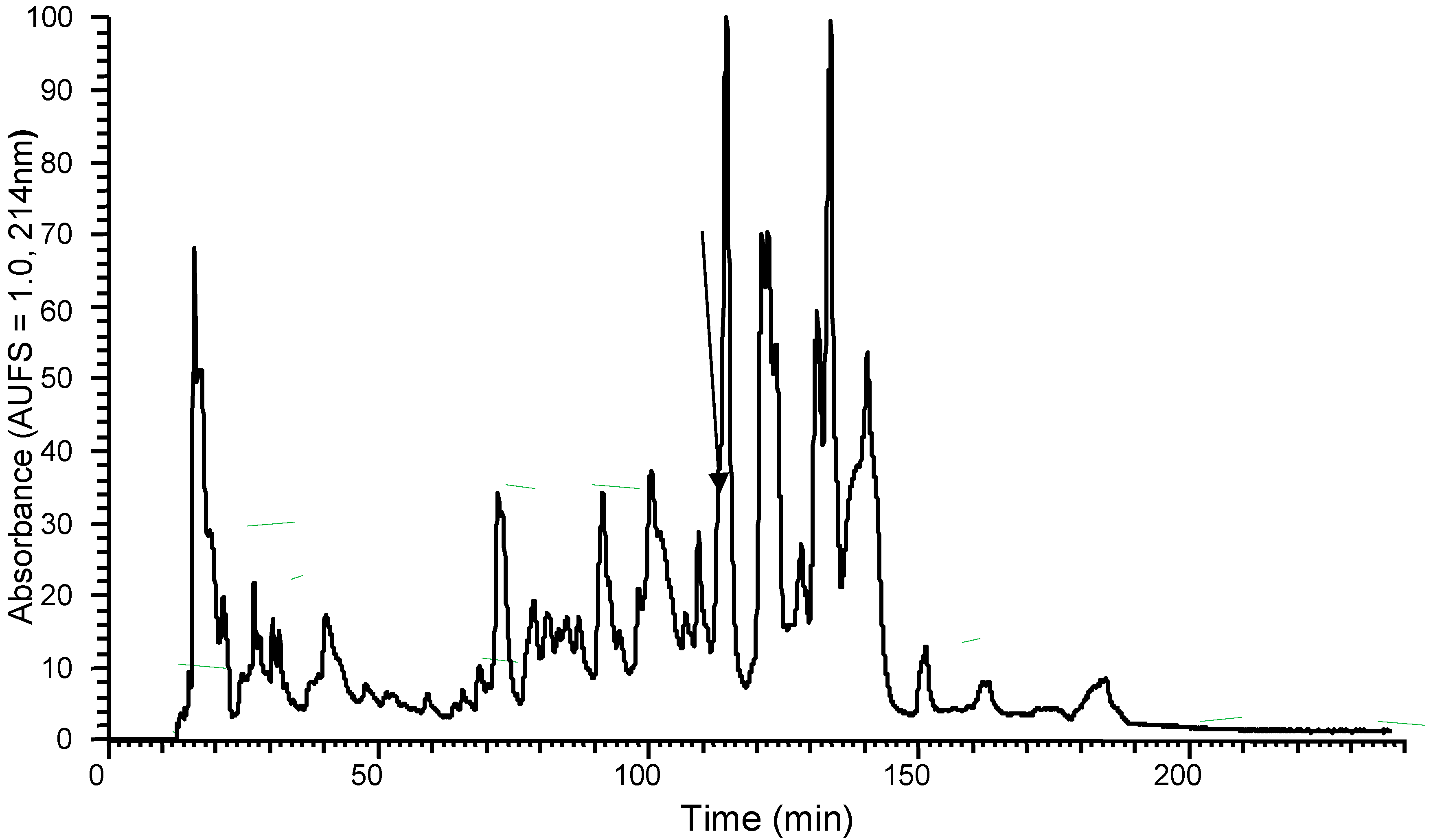
2.2. Isolation and Structural Characterisation of DPS3
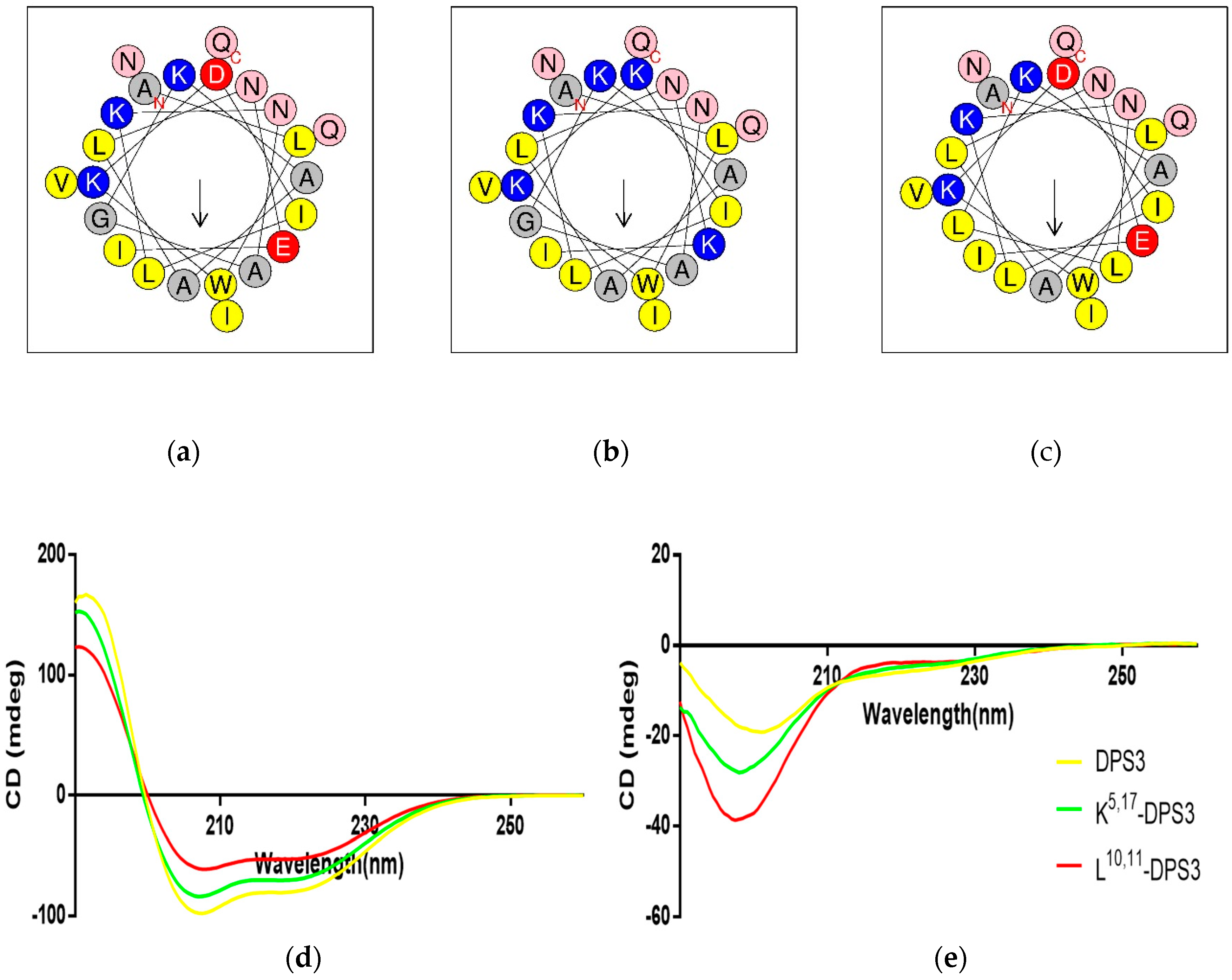
2.3. Physicochemical Properties and Secondary Structures of DPS3 and Its Analogues
2.4. Antimicrobial Activity
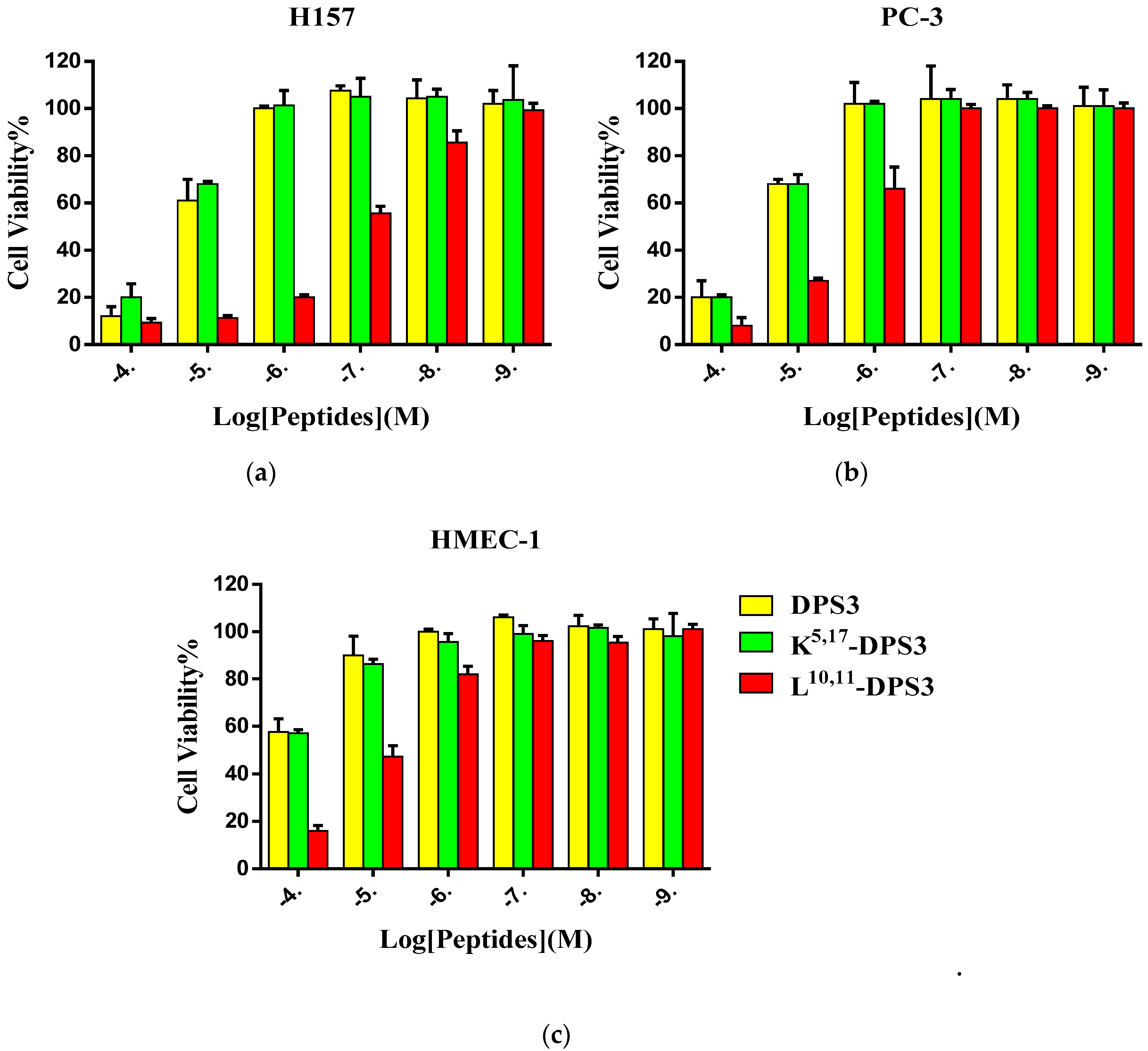
2.5. Cytotoxicity of Peptides on Human Cancer and Normal Cells
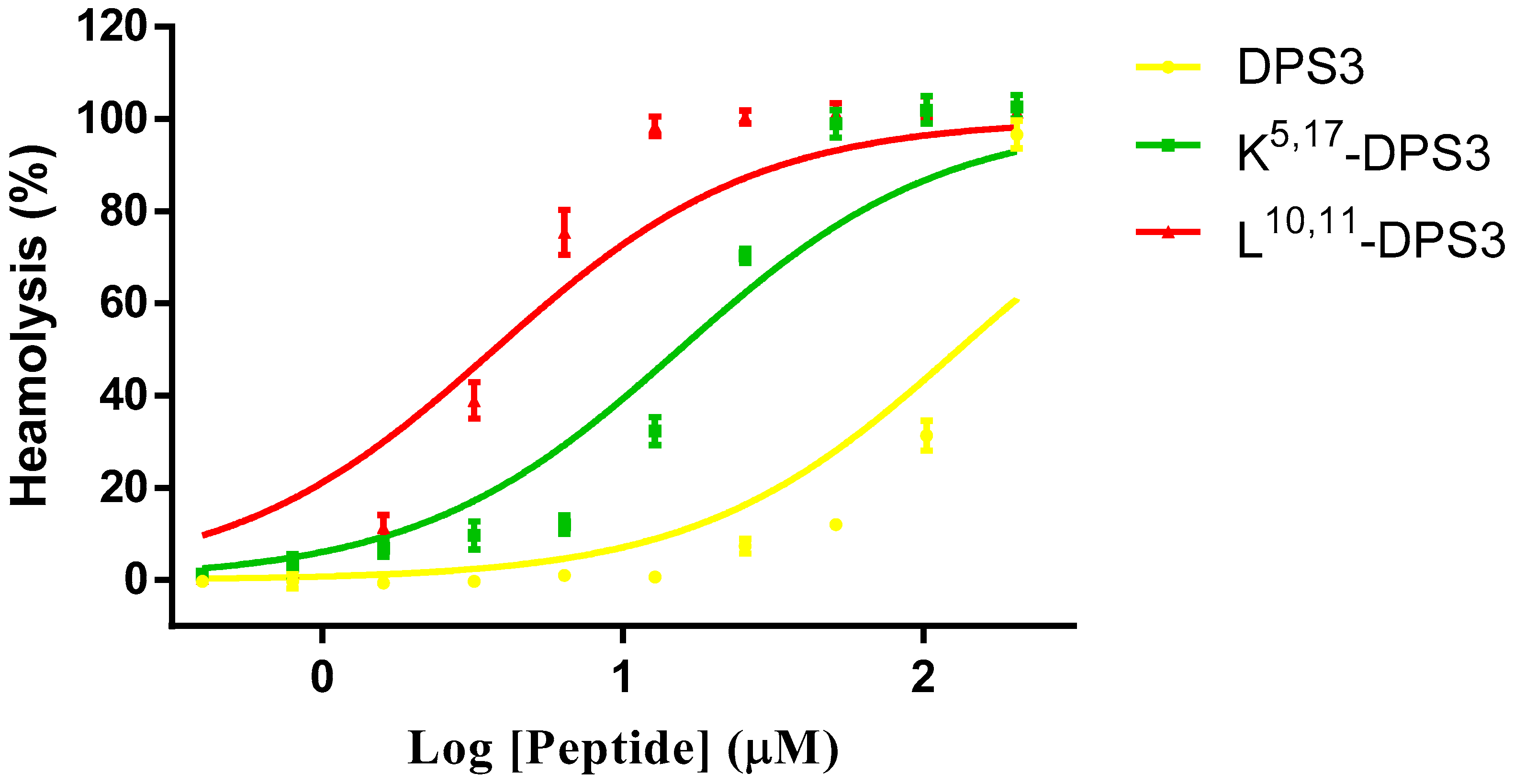
2.6. Haemolysis Activity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Acquisition of Phyllomedusa sauvagii Dermal Secretions
5.2. Shotgun Cloning of a cDNA Encoding DPS3 Peptide Biosynthetic Precursor
5.3. Identification and Analysis of Amino Acid Sequence
5.4. Design and Synthesis of DPS3 and Its Two Analogues
5.5. CD Analysis of Synthetic Peptides
5.6. Antimicrobial Assays
5.7. Cell Viability of Human Cancer and Normal Cells
5.8. Haemolysis Assay
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nicolas, P.; El Amri, C. The dermaseptin superfamily: A gene-based combinatorial library of antimicrobial peptides. Biochim. Biophys. Acta (BBA)-Biomembr. 2009, 1788, 1537–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas, P.; Ladram, A. Dermaseptins. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; Volume 3, pp. 350–363. ISBN 9780123850959. [Google Scholar]
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef] [PubMed]
- Amiche, M.; Ladram, A.; Nicolas, P. A consistent nomenclature of antimicrobial peptides isolated from frogs of the subfamily Phyllomedusinae. Peptides 2008, 29, 2074–2082. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Van Huong, N.; Delfour, A.; Migliore-Samour, D.; Nicolas, P. Isolation, amino acid sequence and synthesis of dermaseptin, a novel antimicrobial peptide of amphibian skin. Biochemistry 1991, 30, 8824–8830. [Google Scholar] [CrossRef] [PubMed]
- Strahilevitz, J.; Mor, A.; Nicolas, P.; Shai, Y. Spectrum of antimicrobial activity and assembly of dermaseptin-b and its precursor form in phospholipid membranes. Biochemistry 1994, 33, 10951–10960. [Google Scholar] [CrossRef] [PubMed]
- Kustanovich, I.; Shalev, D.E.; Mikhlin, M.; Gaidukov, L.; Mor, A. Structural requirements for potent versus selective cytotoxicity for antimicrobial dermaseptin S4 derivatives. J. Biol. Chem. 2002, 277, 16941–16951. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, C.; Hamadat, S.; Le Saux, K.; Newton, C.; Mazouni, M.; Zargarian, L.; Miro-Padovani, M.; Zadigue, P.; Delbé, J.; Hamma-Kourbali, Y.; et al. Studies of the antitumor mechanism of action of dermaseptin B2, a multifunctional cationic antimicrobial peptide, reveal a partial implication of cell surface glycosaminoglycans. PLoS ONE 2017, 12, e0182926. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.L.; Hancock, R.E. Cationic host defense (antimicrobial) peptides. Curr. Opin. Immunol. 2006, 18, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Schmidtchen, A.; Pasupuleti, M.; Malmsten, M. Effect of hydrophobic modifications in antimicrobial peptides. Adv. Colloid Interface Sci. 2014, 205, 265–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Chen, X.; Zhang, Y.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Chen, T. Identification of novel Amurin-2 variants from the skin secretion of Rana amurensis, and the design of cationicity-enhanced analogues. Biochem. Biophys. Res. Commun. 2018, 497, 943–949. [Google Scholar] [CrossRef] [PubMed]
- Galanth, C.; Abbassi, F.; Lequin, O.; Ayala-Sanmartin, J.; Ladram, A.; Nicolas, P.; Amiche, M. Mechanism of Antibacterial Action of Dermaseptin B2: Interplay between Helix–Hinge–Helix Structure and Membrane Curvature Strain. Biochemistry 2008, 48, 313–327. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.P.; Leite, J.R.S.; Brand, G.D.; Regis, W.B.; Tedesco, A.C.; Azevedo, R.B.; Freitas, S.M.; Bloch, C., Jr. Dermaseptins from Phyllomedusa oreades and Phyllomedusa distincta: Liposomes fusion and/or lysis investigated by fluorescence and atomic force microscopy. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 151, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta (BBA)-Biomembr. 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed]
- Giangaspero, A.; Sandri, L.; Tossi, A. Amphipathic α helical antimicrobial peptides. A systematic study of the effects of structural and physical properties on biological activity. Eur. J. Biochem. 2001, 268, 5589–5600. [Google Scholar] [CrossRef] [PubMed]
- Timofeeva, L.; Kleshcheva, N. Antimicrobial polymers: Mechanism of action, factors of activity, and applications. Appl. Microbiol. Biotechnol. 2011, 89, 475–492. [Google Scholar] [CrossRef] [PubMed]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2012, 11, 37. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, F. Cationic amphiphilic peptides with cancer-selective toxicity. Eur. J. Pharmacol. 2009, 625, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Nicolas, P. Isolation and structure of novel defensive peptides from frog skin. Eur. J. Biochem. 1994, 219, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charpentier, S.; Amiche, M.; Mester, J.; Vouille, V.; Le Caer, J.P.; Nicolas, P.; Delfour, A. Structure, synthesis, and molecular cloning of dermaseptins B, a family of skin peptide antibiotics. J. Biol. Chem. 1998, 273, 14690–14697. [Google Scholar] [CrossRef] [PubMed]
- Riedl, S.; Zweytick, D.; Lohner, K. Membrane-active host defense peptides–challenges and perspectives for the development of novel anticancer drugs. Chem. Phys. Lipids 2011, 164, 766–781. [Google Scholar] [CrossRef] [PubMed]
- Van Zoggel, H.; Carpentier, G.; Dos Santos, C.; Hamma-Kourbali, Y.; Courty, J.; Amiche, M.; Delbé, J. Antitumor and angiostatic activities of the antimicrobial peptide dermaseptin B2. PLoS ONE 2012, 7, e44351. [Google Scholar] [CrossRef] [PubMed]
- Belmadani, A.; Semlali, A.; Rouabhia, M. Dermaseptin-S1 decreases Candida albicans growth, biofilm formation and the expression of hyphal wall protein 1 and aspartic protease genes. J. Appl. Microbiol. 2018, 125, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Nicolas, P. The NH2-terminal alpha-helical domain 1–18 of dermaseptin is responsible for antimicrobial activity. J. Biol. Chem. 1994, 269, 1934–1939. [Google Scholar] [PubMed]
- Hancock, R.E.; Chapple, D.S. Peptide antibiotics. Antimicrob. Agents Chemother. 1999, 43, 1317–1323. [Google Scholar] [CrossRef]
- Lequin, O.; Ladram, A.; Chabbert, L.; Bruston, F.; Convert, O.; Vanhoye, D.; Chassaing, G.; Nicolas, P.; Amiche, M. Dermaseptin S9, an α-helical antimicrobial peptide with a hydrophobic core and cationic termini. Biochemistry 2006, 45, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Andreev, K.; Bianchi, C.; Laursen, J.S.; Citterio, L.; Hein-Kristensen, L.; Gram, L.; Kuzmenko, I.; Olsen, C.A.; Gidalevitz, D. Guanidino groups greatly enhance the action of antimicrobial peptidomimetics against bacterial cytoplasmic membranes. Biochim. Biophys. Acta (BBA)-Biomembr. 2014, 1838, 2492–2502. [Google Scholar] [CrossRef] [PubMed]
- Andreev, K.; Martynowycz, M.W.; Huang, M.L.; Kuzmenko, I.; Bu, W.; Kirshenbaum, K.; Gidalevitz, D. Hydrophobic interactions modulate antimicrobial peptoid selectivity towards anionic lipid membranes. Biochim. Biophys. Acta (BBA)-Biomembr. 2018, 1860, 1414–1423. [Google Scholar] [CrossRef] [PubMed]
- Miltz, J.; Rydlo, T.; Mor, A.; Polyakov, V. Potency evaluation of a dermaseptin S4 derivative for antimicrobial food packaging applications. Packag. Technol. Sci. Int. J. 2006, 19, 345–354. [Google Scholar] [CrossRef]
- Tyler, M.J.; Stone, D.J.; Bowie, J.H. A novel method for the release and collection of dermal, glandular secretions from the skin of frogs. J. Pharmacol. Toxicol. Methods 1992, 28, 199–200. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Zhou, M.; Duan, J.; Bininda-Emonds, O.R.; Chen, T.; et al. Targeted modification of a novel amphibian antimicrobial peptide from Phyllomedusa tarsius to enhance its activity against MRSA and microbial biofilm. Front. Microbiol. 2017, 8, 628. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, L.; Wu, Y.; Wang, L.; Ma, C.; Xi, X.; Bininda-Emonds, O.R.; Shaw, C.; Chen, T.; Zhou, M. Evaluation of the bioactivity of a mastoparan peptide from wasp venom and of its analogues designed through targeted engineering. Int. J. Biol. Sci. 2018, 14, 599. [Google Scholar] [CrossRef] [PubMed]
- Micsonai, A.; Wien, F.; Bulyáki, É.; Kun, J.; Moussong, É.; Lee, Y.H.; Goto, Y.; Réfrégiers, M.; Kardos, J. BeStSel: A web server for accurate protein secondary structure prediction and fold recognition from the circular dichroism spectra. Nucleic Acids Res. 2018, 46, W315–W322. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| #1 | b (1+) | b (2+) | Seq. | y (1+) | y (2+) | #2 |
|---|---|---|---|---|---|---|
| 1 | 72.04440 | 36.52584 | A | 23 | ||
| 2 | 185.12847 | 93.06787 | L | 2478.41923 | 1239.71325 | 22 |
| 3 | 371.20779 | 186.10753 | W | 2365.33516 | 1183.17122 | 21 |
| 4 | 499.30276 | 250.15502 | K | 2179.25584 | 1090.13156 | 20 |
| 5 | 614.32971 | 307.66849 | D | 2051.16087 | 1026.08407 | 19 |
| 6 | 727.41378 | 364.21053 | I | 1936.13392 | 968.57060 | 18 |
| 7 | 840.49785 | 420.75256 | L | 1823.04985 | 912.02856 | 17 |
| 8 | 968.59282 | 484.80005 | K | 1709.96578 | 855.48653 | 16 |
| 9 | 1082.63575 | 541.82151 | N | 1581.87081 | 791.43904 | 15 |
| 10 | 1153.67287 | 577.34007 | A | 1467.82788 | 734.41758 | 14 |
| 11 | 1210.69434 | 605.85081 | G | 1396.79076 | 698.89902 | 13 |
| 12 | 1338.78931 | 669.89829 | K | 1339.76929 | 670.38828 | 12 |
| 13 | 1409.82643 | 705.41685 | A | 1211.67432 | 606.34080 | 11 |
| 14 | 1480.86355 | 740.93541 | A | 1140.63720 | 570.82224 | 10 |
| 15 | 1593.94762 | 797.47745 | L | 1069.60008 | 535.30368 | 9 |
| 16 | 1707.99055 | 854.49891 | N | 956.51601 | 478.76164 | 8 |
| 17 | 1837.03315 | 919.02021 | E | 842.47308 | 421.74018 | 7 |
| 18 | 1950.11722 | 975.56225 | I | 713.43048 | 357.21888 | 6 |
| 19 | 2064.16015 | 1032.58371 | N | 600.34641 | 300.67684 | 5 |
| 20 | 2192.21873 | 1096.61300 | Q | 486.30348 | 243.65538 | 4 |
| 21 | 2305.30280 | 1153.15504 | I | 358.24490 | 179.62609 | 3 |
| 22 | 2404.37122 | 1202.68925 | V | 245.16083 | 123.08405 | 2 |
| 23 | Q-Amidated | 146.09241 | 73.54984 | 1 |
| Peptide | Hydrophobicity (H) | Hydrophobic Moment (µH) | % Helix 1 | Net Charge |
|---|---|---|---|---|
| ALWKDILKNAGKAALNEINQIVQ-NH2 | 0.373 | 0.437 | 44.9 | +2 |
| ALWKKILKNAGKAALNKINQIVQ-NH2 | 0.349 | 0.437 | 39 | +6 |
| ALWKDILKNLLKAALNEINQIVQ-NH2 | 0.508 | 0.517 | 28.8 | +2 |
| Microorganisms | DPS3 | K5, 17-DPS3 | L10, 11-DPS3 |
|---|---|---|---|
| MIC (µM) | |||
| S. aureus (NCTC 10788) | 256 | 8 | 8 |
| E. coli (NCTC 10418) | 32 | 8 | 16 |
| C. albicans (NCYC 1467) | 64 | 4 | 16 |
| Peptide | IC50 for H157 (µM) | IC50 for PC3 (µM) | IC50 for HMEC-1 (µM) |
|---|---|---|---|
| DPS3 | 15.67 | 18.20 | 132.10 |
| K5, 17-DPS3 | 18.20 | 18.20 | 123.00 |
| L10, 11-DPS3 | 0.12 | 1.85 | 8.76 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, Y.; Chen, X.; Ma, C.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Kwok, H.F.; Chen, T. Biological Activities of Cationicity-Enhanced and Hydrophobicity-Optimized Analogues of an Antimicrobial Peptide, Dermaseptin-PS3, from the Skin Secretion of Phyllomedusa sauvagii. Toxins 2018, 10, 320. https://doi.org/10.3390/toxins10080320
Tan Y, Chen X, Ma C, Xi X, Wang L, Zhou M, Burrows JF, Kwok HF, Chen T. Biological Activities of Cationicity-Enhanced and Hydrophobicity-Optimized Analogues of an Antimicrobial Peptide, Dermaseptin-PS3, from the Skin Secretion of Phyllomedusa sauvagii. Toxins. 2018; 10(8):320. https://doi.org/10.3390/toxins10080320
Chicago/Turabian StyleTan, Yining, Xiaoling Chen, Chengbang Ma, Xinping Xi, Lei Wang, Mei Zhou, James F. Burrows, Hang Fai Kwok, and Tianbao Chen. 2018. "Biological Activities of Cationicity-Enhanced and Hydrophobicity-Optimized Analogues of an Antimicrobial Peptide, Dermaseptin-PS3, from the Skin Secretion of Phyllomedusa sauvagii" Toxins 10, no. 8: 320. https://doi.org/10.3390/toxins10080320
APA StyleTan, Y., Chen, X., Ma, C., Xi, X., Wang, L., Zhou, M., Burrows, J. F., Kwok, H. F., & Chen, T. (2018). Biological Activities of Cationicity-Enhanced and Hydrophobicity-Optimized Analogues of an Antimicrobial Peptide, Dermaseptin-PS3, from the Skin Secretion of Phyllomedusa sauvagii. Toxins, 10(8), 320. https://doi.org/10.3390/toxins10080320