Clostridium perfringens Enterotoxin: The Toxin Forms Highly Cation-Selective Channels in Lipid Bilayers



Abstract
:1. Introduction
2. Results
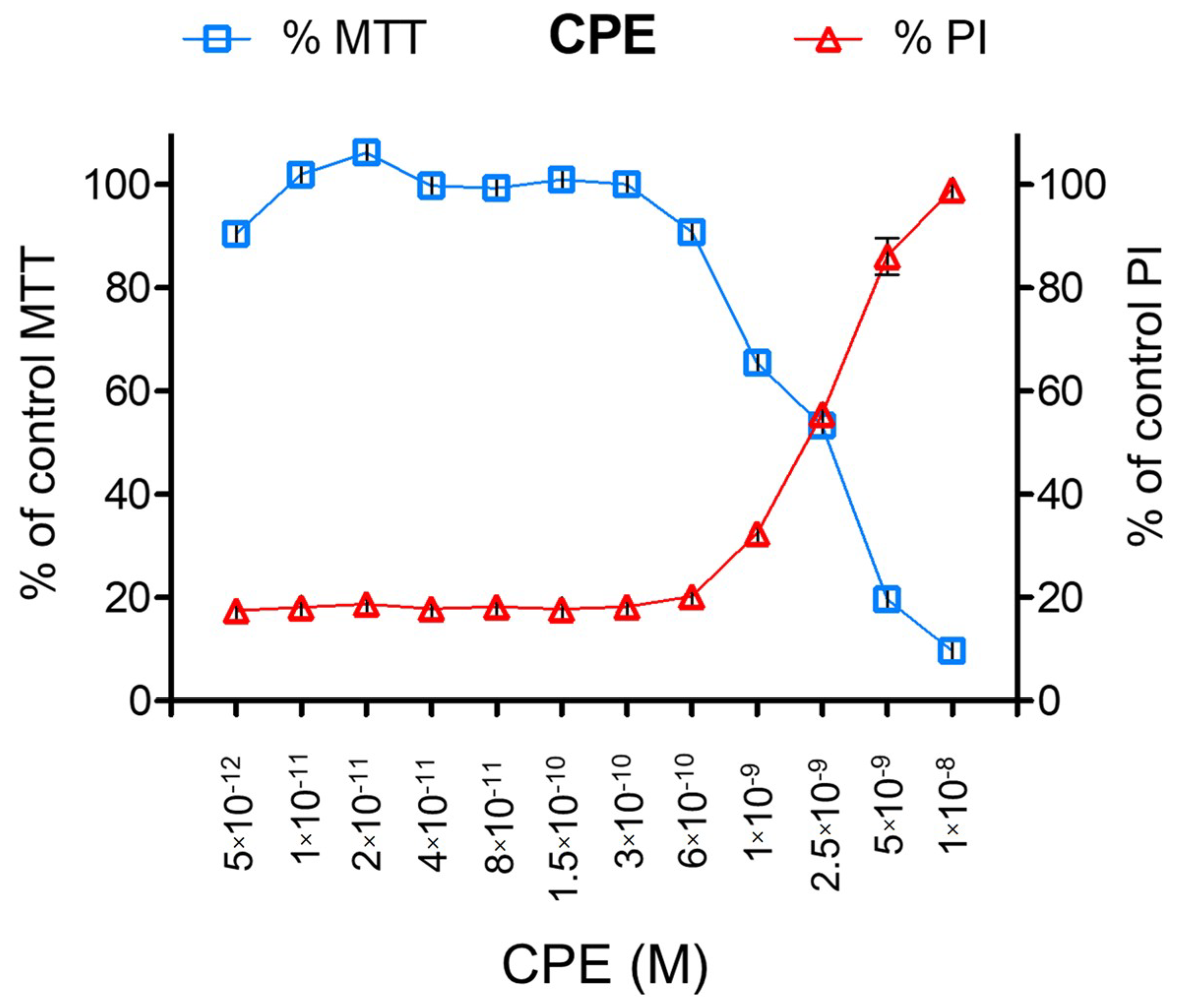
2.1. CPE Pore-Forming Activity on Vero Cells
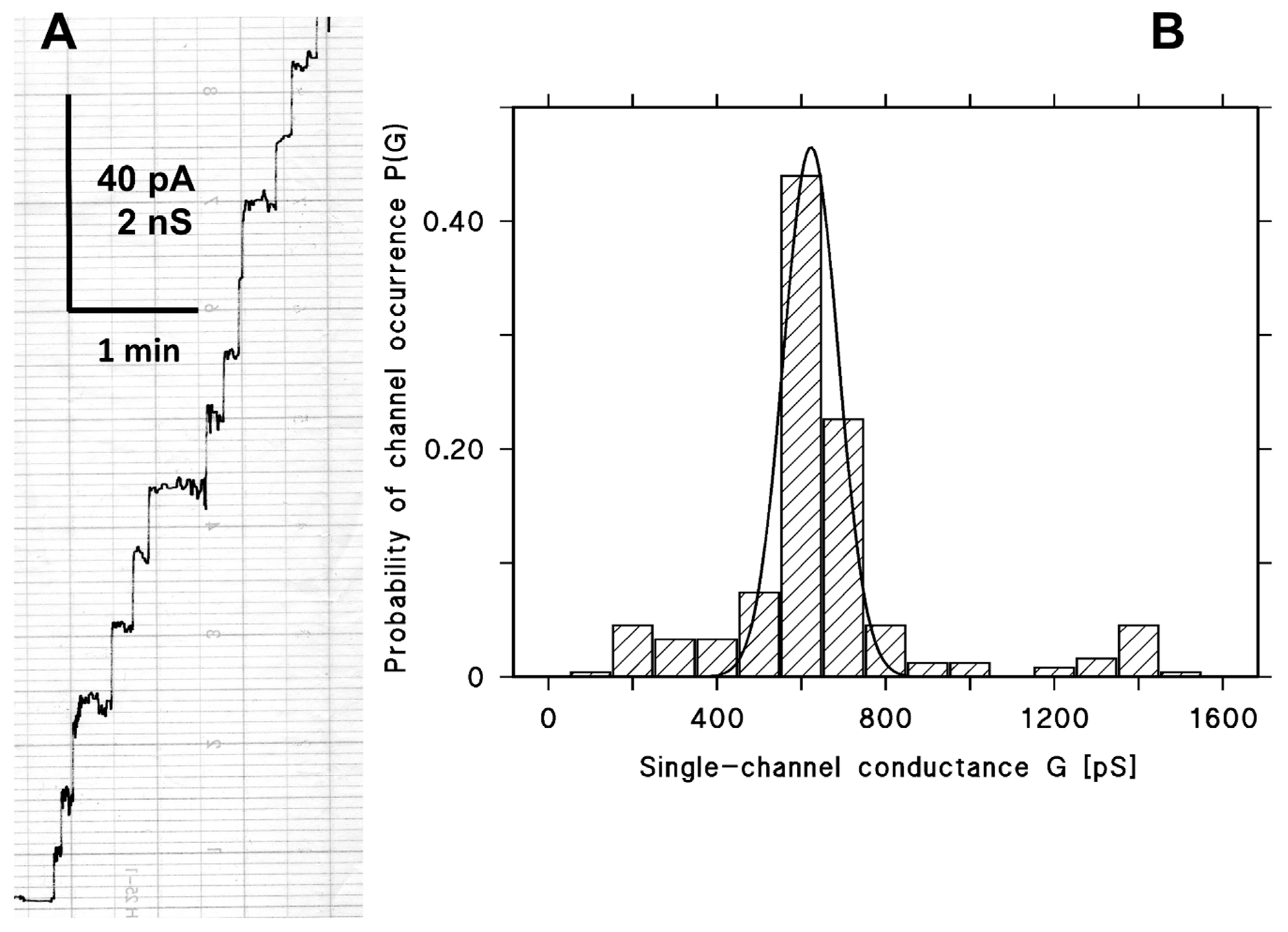
2.2. Pore-Forming Activity of CPE in Lipid Bilayer Membrane
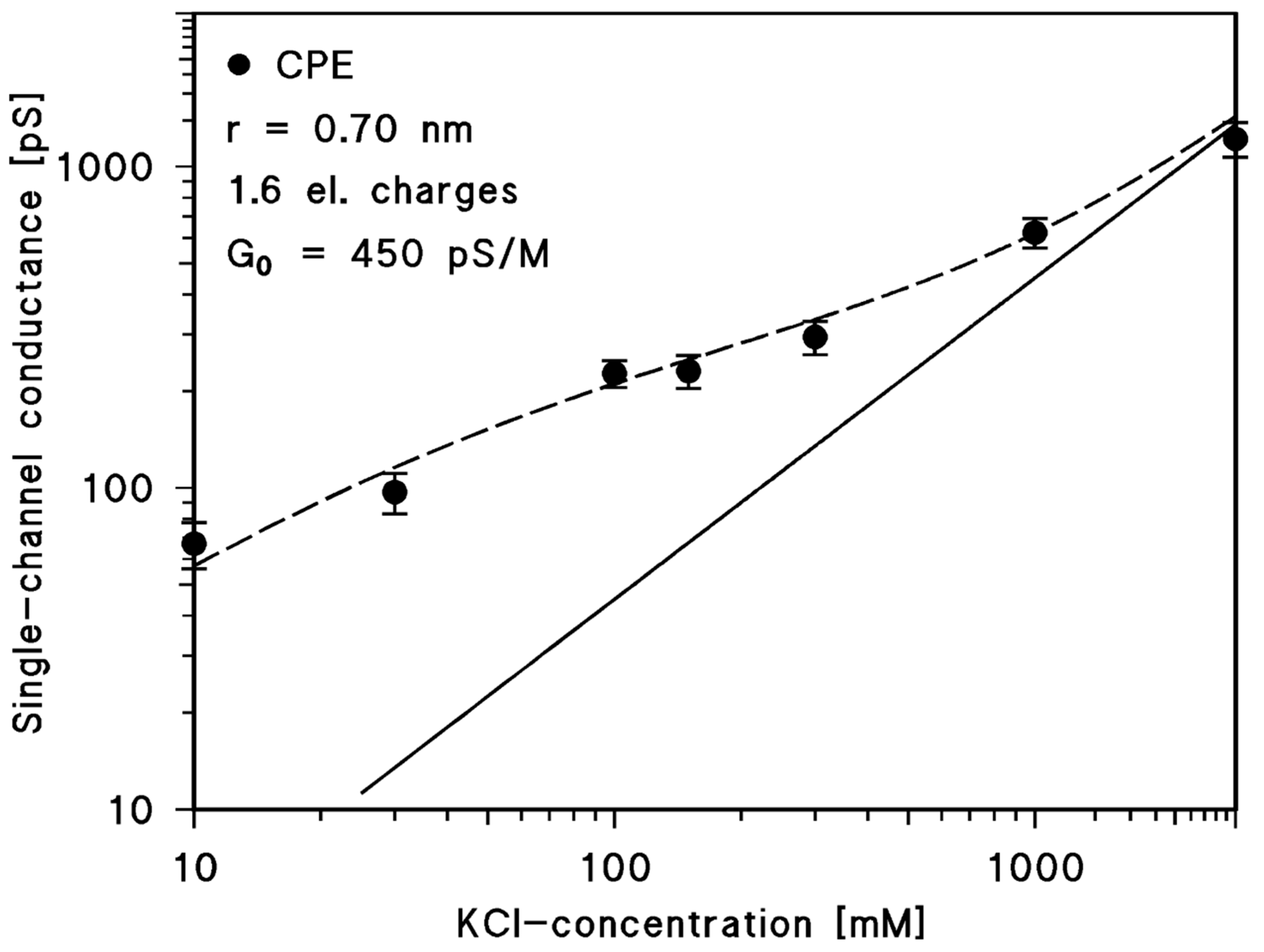
2.3. Single-Channel Conductance of CPE Channel
2.4. Selectivity of the CPE Channel
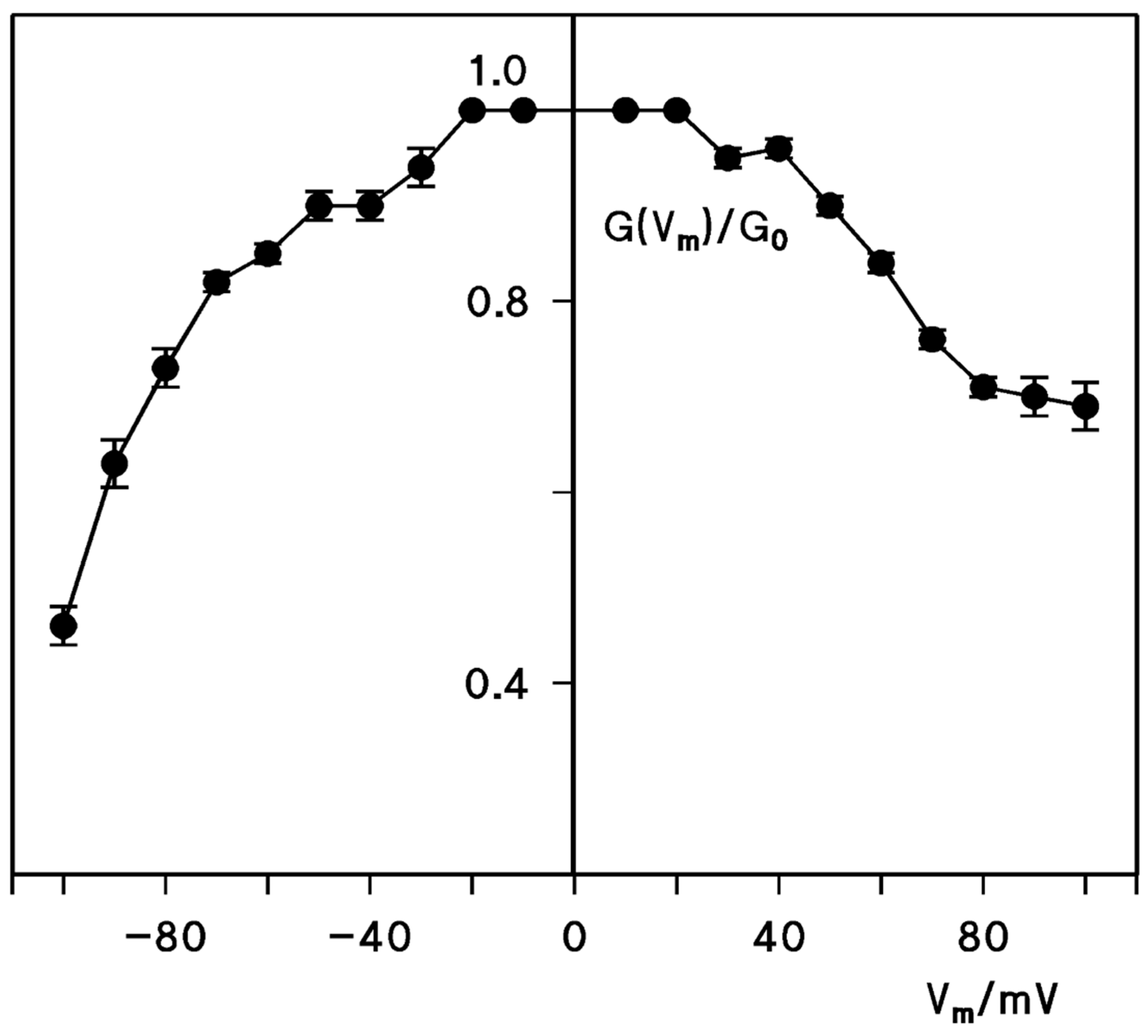
2.5. Voltage Dependence of the CPE Channel
3. Discussion
3.1. CPE Has a High Pore-Forming Capacity
3.2. The Channel Formed by CPE C. perfringens Is Cation Selective
3.3. Comparison with Other Pore-Forming Toxin (PFT) Channels
4. Materials and Methods
4.1. Materials
4.2. Lipid Bilayer Experiments
4.3. Effect of Negatively Charged Groups Attached to the Channel Mouth
4.4. Cell Culture
4.5. Propidium Iodide Influx
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CPE | Clostridium perfringens enterotoxin |
| Vero cells | African green monkey kidney epithelial cells |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| PI | propidium iodide |
References
- Petit, L.; Gibert, M.; Popoff, M.R. Clostridium perfringens enterotoxin and C. perfringens food poisoning. In Encyclopedia of Food Microbiology; Robinson, R., Batt, C., Patel, P., Eds.; Academic Press: London, UK, 1999; pp. 438–444. [Google Scholar]
- Freedman, J.C.; Shrestha, A.; McClane, B.A. Clostridium perfringens enterotoxin: Action, genetics, and translational applications. Toxins 2016, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G.; et al. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobe 2018. [Google Scholar] [CrossRef] [PubMed]
- Billington, S.J.; Wieckowski, E.U.; Sarker, M.R.; Bueschel, D.; Songer, J.G.; McClane, B.A. Clostridium perfringens type E animal enteritis isolates with highly conserved, silent enterotoxin gene sequences. Infect. Immun. 1998, 66, 4531–4536. [Google Scholar] [PubMed]
- Miyamoto, K.; Yumine, N.; Mimura, K.; Nagahama, M.; Li, J.; McClane, B.A.; Akimoto, S. Identification of novel Clostridium perfringens type e strains that carry an iota toxin plasmid with a functional enterotoxin gene. PLoS ONE 2011, 6, e20376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Paredes-Sabja, D.; Sarker, M.R.; McClane, B.A. Clostridium perfringens sporulation and sporulation-associated toxin production. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Gunzel, D.; Yu, A.S. Claudins and the modulation of tight junction permeability. Physiol. Rev. 2013, 93, 525–569. [Google Scholar] [CrossRef] [PubMed]
- Popoff, M.R.; Geny, B. Multifaceted role of Rho, Rac, Cdc42 and ras in intercellular junctions, lessons from toxins. Biochim. Biophys. Acta 2009, 1788, 797–812. [Google Scholar] [CrossRef] [PubMed]
- Van Itallie, C.M.; Anderson, J.M. Claudin interactions in and out of the tight junction. Tissue Barriers 2013, 1, e25247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, K.; Katahira, J.; Horiguchi, Y.; Sonoda, N.; Furuse, M.; Tsukita, S. Clostridium perfringens enterotoxin binds to the second extracellular loop of claudin-3, a tight junction integral membrane protein. FEBS Lett. 2000, 476, 258–261. [Google Scholar] [CrossRef]
- Mitchell, L.A.; Koval, M. Specificity of interaction between Clostridium perfringens enterotoxin and claudin-family tight junction proteins. Toxins 2010, 2, 1595–1611. [Google Scholar] [CrossRef] [PubMed]
- Eichner, M.; Protze, J.; Piontek, A.; Krause, G.; Piontek, J. Targeting and alteration of tight junctions by bacteria and their virulence factors such as Clostridium perfringens enterotoxin. Pflugers Arch. 2017, 469, 77–90. [Google Scholar] [CrossRef] [PubMed]
- McClane, B.A. The complex interactions between Clostridium perfringens enterotoxin and epithelial tight junctions. Toxicon 2001, 39, 1781–1791. [Google Scholar] [CrossRef]
- Lu, Z.; Ding, L.; Lu, Q.; Chen, Y.H. Claudins in intestines: Distribution and functional significance in health and diseases. Tissue Barriers 2013, 1, e24978. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.P.; Li, J.; Shrestha, A.; Freedman, J.C.; Beingesser, J.; McClane, B.A.; Uzal, F.A. Clostridium perfringens type A enterotoxin damages the rabbit colon. Infect. Immun. 2014, 82, 2211–2218. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Uzal, F.A.; McClane, B.A. The interaction of Clostridium perfringens enterotoxin with receptor claudins. Anaerobe 2016, 41, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Freedman, J.C.; Shrestha, A.; Theoret, J.R.; Garcia, J.; Awad, M.M.; Adams, V.; Moore, R.J.; Rood, J.I.; McClane, B.A. Towards an understanding of the role of Clostridium perfringens toxins in human and animal disease. Future Microbiol. 2014, 9, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; McClane, B.A. Use of Clostridium perfringens enterotoxin and the enterotoxin receptor-binding domain (c-cpe) for cancer treatment: Opportunities and challenges. J. Toxicol. 2012, 2012, 981626. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.L.; Smedley, J.G., 3rd; Singh, U.; Chakrabarti, G.; Van Itallie, C.M.; Anderson, J.M.; McClane, B.A. Compositional and stoichiometric analysis of Clostridium perfringens enterotoxin complexes in caco-2 cells and claudin 4 fibroblast transfectants. Cell. Microbiol. 2007, 9, 2734–2755. [Google Scholar] [CrossRef] [PubMed]
- Veshnyakova, A.; Piontek, J.; Protze, J.; Waziri, N.; Heise, I.; Krause, G. Mechanism of Clostridium perfringens enterotoxin interaction with claudin-3/-4 protein suggests structural modifications of the toxin to target specific claudins. J. Biol. Chem. 2012, 287, 1698–1708. [Google Scholar] [CrossRef] [PubMed]
- Kimura, J.; Abe, H.; Kamitani, S.; Toshima, H.; Fukui, A.; Miyake, M.; Kamata, Y.; Sugita-Konishi, Y.; Yamamoto, S.; Horiguchi, Y. Clostridium perfringens enterotoxin interacts with claudins via electrostatic attraction. J. Biol. Chem. 2010, 285, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, G.; Zhou, X.; McClane, B.A. Death pathways activated in caco-2 cells by Clostridium perfringens enterotoxin. Infect. Immun. 2003, 71, 4260–4270. [Google Scholar] [CrossRef] [PubMed]
- Eichner, M.; Augustin, C.; Fromm, A.; Piontek, A.; Walther, W.; Bucker, R.; Fromm, M.; Krause, G.; Schulzke, J.D.; Gunzel, D.; et al. In colon epithelia, Clostridium perfringens enterotoxin causes focal leaks by targeting claudins which are apically accessible due to tight junction derangement. J. Infect. Dis. 2017, 217, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Ni, C.; Yang, Z.; Piontek, A.; Chen, H.; Wang, S.; Fan, Y.; Qin, Z.; Piontek, J. Specific binding of Clostridium perfringens enterotoxin fragment to claudin-b and modulation of zebrafish epidermal barrier. Exp. Dermatol. 2015, 24, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, N.; Furuse, M.; Sasaki, H.; Yonemura, S.; Katahira, J.; Horiguchi, Y.; Tsukita, S. Clostridium perfringens enterotoxin fragment removes specific claudin from tight junction strands: Evidence for direct involvement of claudin in tight junction barrier. J. Cell Biol. 1999, 147, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, T.; Shinya, N.; Ito, K.; Ohsawa, N.; Terada, T.; Hirata, K.; Kawano, Y.; Yamamoto, M.; Kimura-Someya, T.; Yokoyama, S.; et al. Structural basis for disruption of claudin assembly in tight junctions by an enterotoxin. Sci. Rep. 2016, 6, 33632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briggs, D.C.; Naylor, C.E.; Smedley, J.G., 3rd; Lukoyanova, N.; Robertson, S.; Moss, D.S.; McClane, B.A.; Basak, A.K. Structure of the food-poisoning Clostridium perfringens enterotoxin reveals similarity to the aerolysin-like pore-forming toxins. J. Mol. Biol. 2011, 413, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Kitadokoro, K.; Nishimura, K.; Kamitani, S.; Fukui-Miyazaki, A.; Toshima, H.; Abe, H.; Kamata, Y.; Sugita-Konishi, Y.; Yamamoto, S.; Karatani, H.; et al. Crystal structure of Clostridium perfringens enterotoxin displays features of beta-pore-forming toxins. J. Biol. Chem. 2011, 286, 19549–19555. [Google Scholar] [CrossRef] [PubMed]
- Smedley, J.G., 3rd; Uzal, F.A.; McClane, B.A. Identification of a prepore large-complex stage in the mechanism of action of Clostridium perfringens enterotoxin. Infect. Immun. 2007, 75, 2381–2390. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Theoret, J.R.; Shrestha, A.; Smedley, J.G., 3rd; McClane, B.A. Cysteine-scanning mutagenesis supports the importance of Clostridium perfringens enterotoxin amino acids 80 to 106 for membrane insertion and pore formation. Infect. Immun. 2012, 80, 4078–4088. [Google Scholar] [CrossRef] [PubMed]
- Hardy, S.P.; Ritchie, C.; Allen, M.C.; Ashley, R.H.; Granum, P.E. Clostridium perfringens type A enterotoxin forms mepacrine-sensitive pores in pure phospholipid bilayers in the absence of putative receptor proteins. Biochim. Biophys. Acta 2001, 1515, 38–43. [Google Scholar] [CrossRef]
- Sugimoto, N.; Takagi, M.; Ozutsumi, K.; Harada, S.; Matsuda, M. Enterotoxin of Clostridium perfringens type A forms ion-permeable channels in a lipid bilayer membrane. Biochem. Biophys. Res. Commun. 1988, 156, 551–556. [Google Scholar] [CrossRef]
- Hardy, S.P.; Denmead, M.; Parekh, N.; Granum, P.E. Cationic currents induced by Clostridium perfringens type a enterotoxin in human intestinal caco-2 cells. J. Med. Microbiol. 1999, 48, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Riess, F.G.; Lichtinger, T.; Cseh, R.; Yassin, A.F.; Schaal, K.P.; Benz, R. The cell wall porin of Nocardia farcinica: Biochemical identification of the channel-forming protein and biophysical characterization of the channel properties. Mol. Microbiol. 1998, 29, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Trias, J.; Benz, R. Characterization of the channel formed by the mycobacterial porin in lipid bilayer membranes. Demonstration of voltage gating and of negative point charges at the channel mouth. J. Biol. Chem. 1993, 268, 6234–6240. [Google Scholar] [PubMed]
- Benz, R.; Maier, E.; Ladant, D.; Ullmann, A.; Sebo, P. Adenylate cyclase toxin (cyaa) of Bordetella pertussis. Evidence for the formation of small ion-permeable channels and comparison with hlya of Escherichia coli. J. Biol. Chem. 1994, 269, 27231–27239. [Google Scholar] [PubMed]
- Hu, D.; Sugii, S.; Kusunoki, H.; Uemura, T. Flow cytometric assay for cytotoxic activity of crude Clostridium perfringens enterotoxin using non-adherent cell fm3a. FEMS Immunol. Med. Microbiol. 1995, 12, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Petit, L.; Maier, E.; Gibert, M.; Popoff, M.R.; Benz, R. Clostridium perfringens epsilon-toxin induces a rapid change in cell membrane permeability to ions and forms channels in artificial lipid bilayers. J. Biol. Chem. 2001, 276, 15736–15740. [Google Scholar] [CrossRef] [PubMed]
- Benz, R. Solute uptake through bacterial outer membrane. In Bacterial Cell Wall; Ghuyen, J.M., Hakenbeck, R., Eds.; Elsevier Sciences B. V.: Amsterdam, The Netherlands, 1994; pp. 397–423. [Google Scholar]
- Benz, R.; Janko, K.; Läuger, P. Ionic selectivity of pores formed by the matrix protein (porin) of Escherichia coli. Biochim. Biophys. Acta 1979, 551, 238–247. [Google Scholar] [CrossRef]
- Benz, R.; Schmid, A.; Wagner, W.; Goebel, W. Pore formation by the Escherichia coli hemolysin: Evidence for an association-dissociation equilibrium of the pore-forming aggregates. Infect. Immun. 1989, 57, 887–895. [Google Scholar] [PubMed]
- Schmid, A.; Benz, R.; Just, I.; Aktories, K. Interaction of Clostridium botulinum C2 toxin with lipid bilayer membranes. J. Biol. Chem. 1994, 269, 16706–16711. [Google Scholar] [PubMed]
- Cole, A.R.; Gibert, M.; Popoff, M.R.; Moss, D.S.; Titball, R.W.; Basak, A. Clostridium perfringens ε-toxin shows structural similarity to the pore-forming toxin aerolysin. Nat. Struct. Mol. Biol. 2004, 11, 797–798. [Google Scholar] [CrossRef] [PubMed]
- Knapp, O.; Benz, R.; Popoff, M.R. Pore-forming activity of clostridial binary toxins. Biochim. Biophys. Acta 2016, 1858, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Orlik, F.; Schiffler, B.; Benz, R. Anthrax toxin protective antigen: Inhibition of channel function by chloroquine and related compounds and study of binding kinetics using the current noise analysis. Biophys. J. 2005, 88, 1715–1724. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.P.; McQuarrie, D.A. The effect of discrete charges on the electrical properties of a membrane. I. J. Theor. Biol. 1975, 55, 13–27. [Google Scholar] [CrossRef]
- Popoff, M.R. Clostridial pore-forming toxins: Powerful virulence factors. Anaerobe 2014, 30, 220–238. [Google Scholar] [CrossRef] [PubMed]
- Podobnik, M.; Kisovec, M.; Anderluh, G. Molecular mechanism of pore formation by aerolysin-like proteins. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef] [PubMed]
- Knapp, O.; Maier, E.; Benz, R.; Geny, B.; Popoff, M.R. Identification of the channel-forming domain of Clostridium perfringens epsilon-toxin (etx). Biochim. Biophys. Acta 2009, 1788, 2584–2593. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, T.; Schmid, A.; Notermans, S.; Benz, R. Aerolysin of Aeromonas sobria: Evidence for the formation of ion-permeable channels and comparison with alpha-toxin of Staphylococcus aureus. Infect. Immun. 1990, 58, 2127–2132. [Google Scholar] [PubMed]
- Song, L.; Hobaugh, M.R.; Shustak, C.; Cheley, S.; Bayley, H.; Gouaux, J.E. Structure of staphylococcal alpha-hemolysin, a heptameric transmembrane pore. Science 1996, 274, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Bokori-Brown, M.; Savva, C.G.; Fernandes da Costa, S.P.; Naylor, C.E.; Basak, A.K.; Titball, R.W. Molecular basis of toxicity of Clostridium perfringens epsilon toxin. FEBS J. 2011, 278, 4589–4601. [Google Scholar] [CrossRef] [PubMed]
- Hang’ombe, M.B.; Mukamoto, M.; Kohda, T.; Sugimoto, N.; Kozaki, S. Cytotoxicity of Clostridium septicum alpha-toxin: Its oligomerization in detergent resistant membranes of mammalian cells. Microb. Pathog. 2004, 37, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Knapp, O.; Maier, E.; Mkaddem, S.B.; Benz, R.; Bens, M.; Chenal, A.; Geny, B.; Vandewalle, A.; Popoff, M.R. Clostridium septicum alpha-toxin forms pores and induces rapid cell necrosis. Toxicon 2010, 55, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Degiacomi, M.T.; Iacovache, I.; Pernot, L.; Chami, M.; Kudryashev, M.; Stahlberg, H.; van der Goot, F.G.; Dal Peraro, M. Molecular assembly of the aerolysin pore reveals a swirling membrane-insertion mechanism. Nat. Chem. Biol. 2013, 9, 623–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yelland, T.S.; Naylor, C.E.; Bagoban, T.; Savva, C.G.; Moss, D.S.; McClane, B.A.; Blasig, I.E.; Popoff, M.; Basak, A.K. Structure of a C. perfringens enterotoxin mutant in complex with a modified claudin-2 extracellular loop 2. J. Mol. Biol. 2014, 426, 3134–3147. [Google Scholar] [CrossRef] [PubMed]
- Benz, R.; Janko, K.; Boos, W.; Läuger, P. Formation of large, ion-permeable membrane channels by the matrix protein (porin) of Escherichia coli. Biochim. Biophys. Acta 1978, 511, 305–319. [Google Scholar] [CrossRef]
- Petit, L.; Gibert, M.; Gillet, D.; Laurent-Winter, C.; Boquet, P.; Popoff, M.R. Clostridium perfringens epsilon-toxin acts on mdck cells by forming a large membrane complex. J. Bacteriol. 1997, 179, 6480–6487. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Electrolyte | Concentration (M) | G (pS) | N |
|---|---|---|---|
| KCL | 0.01 | 67 ± 11 | 113 |
| 0.03 | 97 ± 14 | 94 | |
| 0.1 | 227 ± 22 | 118 | |
| 0.15 | 231 ± 27 | 154 | |
| 0.3 | 295 ± 35 | 149 | |
| 1.0 | 623 ± 65 | 243 | |
| 3.0 | 1220 ± 150 | 261 | |
| LiCl | 1.0 | 211 ± 22 | 96 |
| KCH3COO (pH 7) | 1.0 | 619 ± 71 | 162 |
| Electrolyte | Vm (mV) | Permeability Ratios Pcation/Panion |
|---|---|---|
| KCl | 33.3 ± 2.4 | 11.4 |
| LiCl | 29.7 ± 1.7 | 8.3 |
| KCH3COO (pH 7) | 34.3 ± 2.3 | 13.0 |
| Toxin | Single-Channel Conductance in 1 M KCl | Zero-Current Membrane Potential for 5-Fold KCl Gradients | Permeability Ratio PK/PCl |
|---|---|---|---|
| G [pS] | Vm [mV] | PK/PCl | |
| CPE | 623 | +33 | 11.4 |
| Aerolysin | 650 | −24 | 0.21 |
| S. aureus α-toxin | 820 | −22 | 0.25 |
| ε-toxin | 550 | −19 | 0.30 |
| C. septicum alpha-toxin | 1250 | −15 | 0.38 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benz, R.; Popoff, M.R. Clostridium perfringens Enterotoxin: The Toxin Forms Highly Cation-Selective Channels in Lipid Bilayers. Toxins 2018, 10, 341. https://doi.org/10.3390/toxins10090341
Benz R, Popoff MR. Clostridium perfringens Enterotoxin: The Toxin Forms Highly Cation-Selective Channels in Lipid Bilayers. Toxins. 2018; 10(9):341. https://doi.org/10.3390/toxins10090341
Chicago/Turabian StyleBenz, Roland, and Michel R. Popoff. 2018. "Clostridium perfringens Enterotoxin: The Toxin Forms Highly Cation-Selective Channels in Lipid Bilayers" Toxins 10, no. 9: 341. https://doi.org/10.3390/toxins10090341
APA StyleBenz, R., & Popoff, M. R. (2018). Clostridium perfringens Enterotoxin: The Toxin Forms Highly Cation-Selective Channels in Lipid Bilayers. Toxins, 10(9), 341. https://doi.org/10.3390/toxins10090341