Venom Proteome of Spine-Bellied Sea Snake (Hydrophis curtus) from Penang, Malaysia: Toxicity Correlation, Immunoprofiling and Cross-Neutralization by Sea Snake Antivenom

Abstract
:
1. Introduction
2. Results
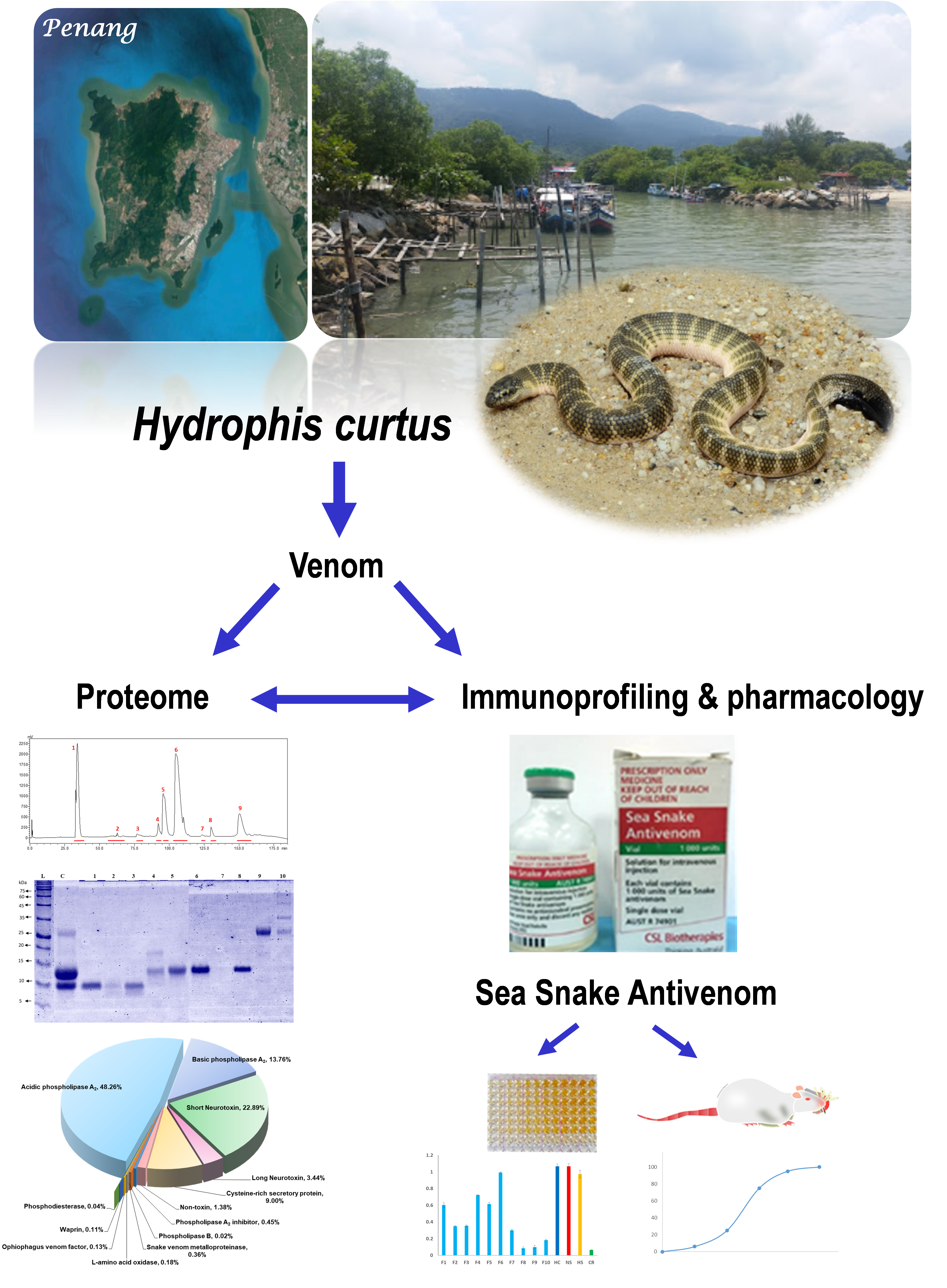
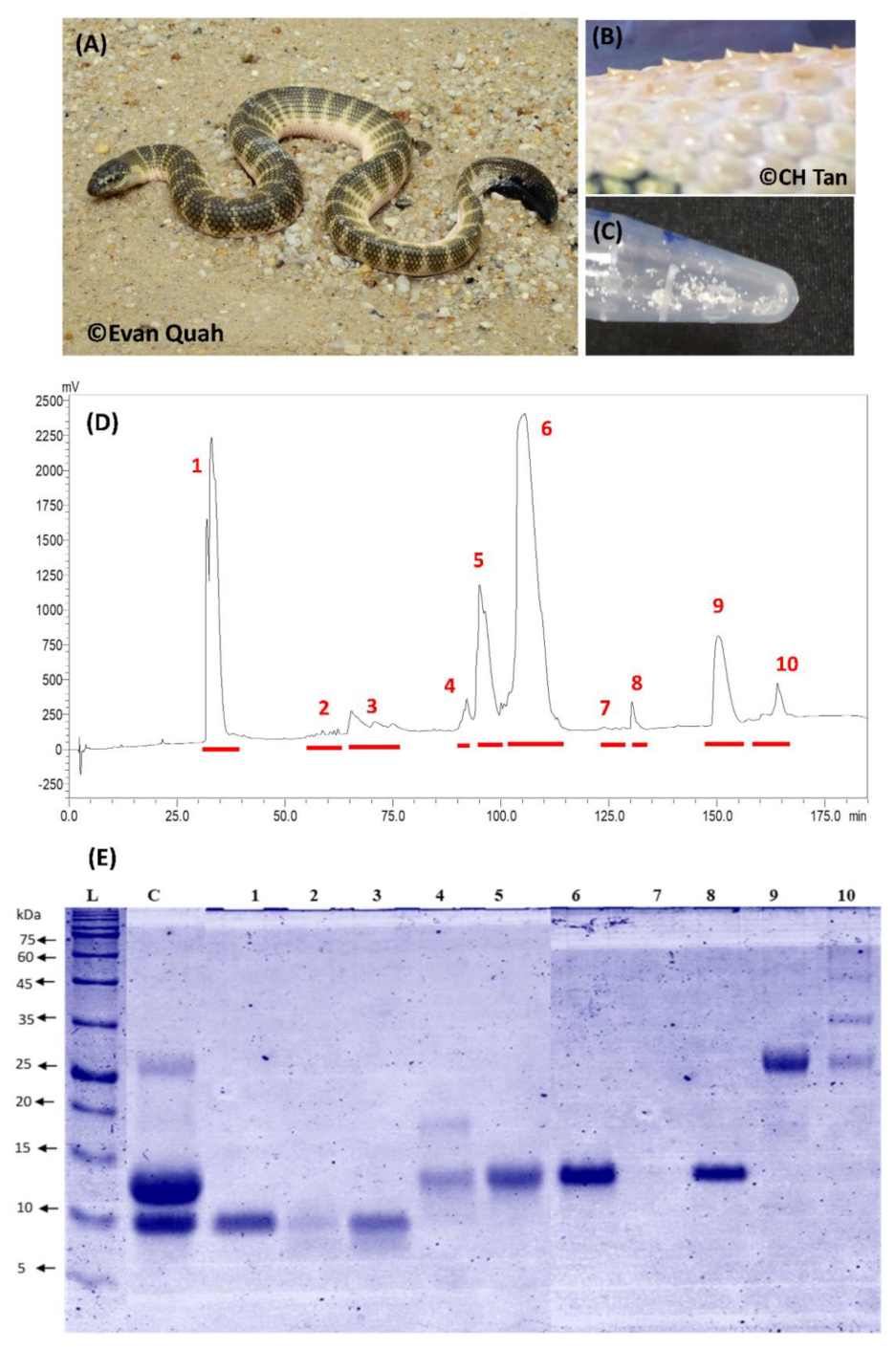
2.1. Decomplexation Proteomics of Penang H. curtus Venom
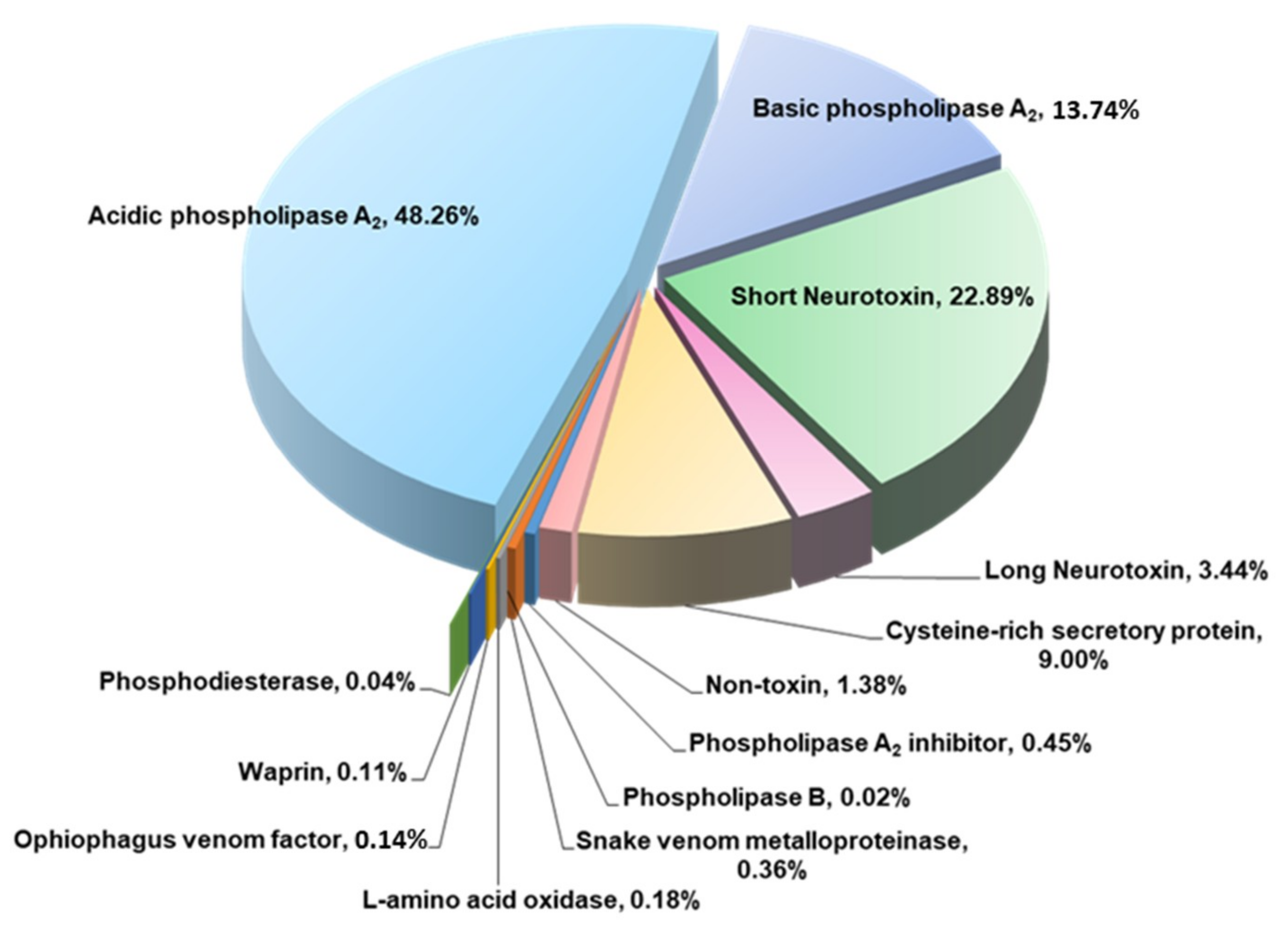
2.2. Proteome of Penang H. curtus Venom
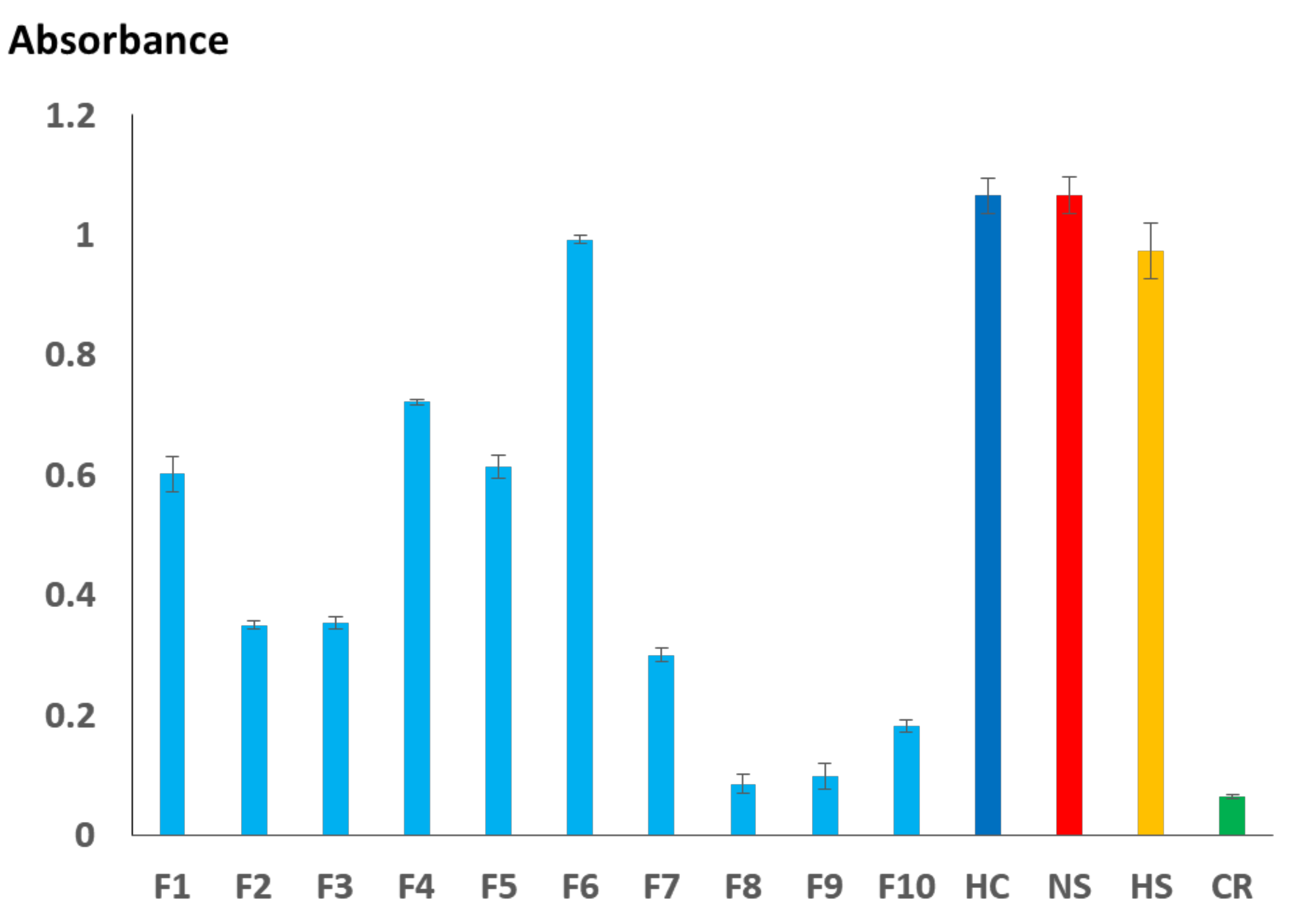
2.3. Immunoreactivity Profiling of H. curtus Venom Proteins
2.4. Toxicity of H. curtus Venom and Principal Toxins
2.5. Cross-Neutralization of H. curtus Venom and Toxins by SSAV
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Samples and Chemicals
5.2. Reverse-Phase High Performance Liquid Chromatography
5.3. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
5.4. In-Solution Protein Digestion with Trypsin
5.5. Nano-ESI-LCMS/MS of the Tryptic Digests and Label-Free LCMS/MS Protein Quantitation
5.6. Median Lethal Dose of Venom/Toxin and Efficacy of Neutralization by Antivenom
5.7. Immunological and Antigenic Profiling of H. curtus Venom Fractions
5.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Slowinski, J.B.; Keogh, J.S. Phylogenetic relationships of elapid snakes based on cytochrome b mtDNA sequences. Mol. Phylogenet. Evol. 2000, 15, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.L.; Lee, M.S.; Leys, R.; Foster, R.; Keogh, J.S. Molecular phylogeny and divergence dates for Australasian elapids and sea snakes (hydrophiinae): Evidence from seven genes for rapid evolutionary radiations. J. Evol. Biol. 2008, 21, 682–695. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.L.; Lee, M.S.; Mumpuni; Bertozzi, T.; Rasmussen, A.R. Multilocus phylogeny and recent rapid radiation of the viviparous sea snakes (Elapidae: Hydrophiinae). Mol. Phylogenet. Evol. 2013, 66, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Reid, H.A. Epidemiology of sea-snake bites. J. Trop. Med. Hyg. 1975, 78, 106–113. [Google Scholar]
- Reid, H.A.; Lim, K.J. Sea-snake bite; a survey of fishing villages in northwest Malaya. Br. Med. J. 1957, 2, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Kularatne, S.A.; Hettiarachchi, R.; Dalpathadu, J.; Mendis, A.S.; Appuhamy, P.D.; Zoysa, H.D.; Maduwage, K.; Weerasinghe, V.S.; de Silva, A. Enhydrina schistosa (Elapidae: Hydrophiinae) the most dangerous sea snake in Sri Lanka: Three case studies of severe envenoming. Toxicon 2014, 77, 78–86. [Google Scholar] [CrossRef]
- Vithanage, K.K.; Thirumavalavan, K. A case of a sea snake bite resulting in fatal envenoming. Ceylon Med. J. 2012, 57, 174–175. [Google Scholar] [CrossRef]
- Cao, N.; Tao, N.T.; Moore, A.; Montoya, A.; Rasmussen, A.R.; Broad, K.; Voris, H.K.; Takacs, Z. Sea Snake Harvest in the Gulf of Thailand. Conserv. Biol. 2014, 28, 1677–1687. [Google Scholar] [CrossRef]
- WHO. Guidelines for the Management of Snake Bites; Regional office for South-East Asia: New Delhi, India, 2016. [Google Scholar]
- Reid, H.A. Symptomatology, Pathology and Treatment of the Bites of Sea Snakes. In Snake Venoms; Lee, C.-Y., Ed.; Springer: Berlin/Heidelberg, Germany, 1979; Volume 52, pp. 922–955. [Google Scholar]
- Gopalakrishnakone, P.; Ponraj, D.; Thwinn, M.M. Myotoxic Phospholipases from Snake Venoms: General Myoglobinuric and Local Myonecrotic Toxins; John Wiley and Sons: New York, NY, USA, 1997. [Google Scholar]
- Tan, C.H.; Tan, K.Y.; Lim, S.E.; Tan, N.H. Venomics of the beaked sea snake, Hydrophis schistosus: A minimalist toxin arsenal and its cross-neutralization by heterologous antivenoms. J. Proteom. 2015, 126, 121–130. [Google Scholar] [CrossRef]
- Brook, G.A.; Torres, L.F.; Gopalakrishnakone, P.; Duchen, L.W. Effects of phospholipase of Enhydrina schistosa venom on nerve, motor end-plate and muscle of the mouse. Q. J. Exp. Physiol. 1987, 72, 571–591. [Google Scholar] [CrossRef]
- Mackessy, S.P.; Tu, A.T. Biology of the sea snakes and biochemistry of their venoms. In Toxin-Related Diseases: Poisons Originating from Plants, Animals and Spoilage; Tu, A.T., Ed.; Oxford & IBH Publishing Co.: New Delhi, India, 1993; pp. 305–351. [Google Scholar]
- Fry, B.G.; Wuster, W.; Ryan Ramjan, S.F.; Jackson, T.; Martelli, P.; Kini, R.M. Analysis of Colubroidea snake venoms by liquid chromatography with mass spectrometry: Evolutionary and toxinological implications. Rapid Commun. Mass Spectrom. 2003, 17, 2047–2062. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J.; Ghezellou, P.; Paiva, O.; Matainaho, T.; Ghassempour, A.; Goudarzi, H.; Kraus, F.; Sanz, L.; Williams, D.J. Snake venomics of two poorly known Hydrophiinae: Comparative proteomics of the venoms of terrestrial Toxicocalamus longissimus and marine Hydrophis cyanocinctus. J. Proteom. 2012, 75, 4091–4101. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Pla, D.; Sasa, M.; Tsai, W.C.; Solorzano, A.; Urena-Diaz, J.M.; Fernandez-Montes, M.L.; Mora-Obando, D.; Sanz, L.; Gutierrez, J.M.; et al. Two color morphs of the pelagic yellow-bellied sea snake, Pelamis platura, from different locations of Costa Rica: Snake venomics, toxicity, and neutralization by antivenom. J. Proteom. 2014, 103, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Gutierrez, J.M.; Rasmussen, A.R.; Engmark, M.; Gravlund, P.; Sanders, K.L.; Lohse, B.; Lomonte, B. Danger in the reef: Proteome, toxicity, and neutralization of the venom of the olive sea snake, Aipysurus laevis. Toxicon 2015, 107, 187–196. [Google Scholar] [CrossRef]
- Tan, C.H.; Wong, K.Y.; Tan, K.Y.; Tan, N.H. Venom proteome of the yellow-lipped sea krait, Laticauda colubrina from Bali: Insights into subvenomic diversity, venom antigenicity and cross-neutralization by antivenom. J. Proteom. 2017, 166, 48–58. [Google Scholar] [CrossRef]
- Neale, V.; Sotillo, J.; Seymour, J.E.; Wilson, D. The venom of the spine-bellied sea snake (Hydrophis curtus): Proteome, Toxin diversity and intraspecific variation. Int. J. Mol. Sci. 2017, 18, 2695. [Google Scholar] [CrossRef]
- Pahari, S.; Bickford, D.; Fry, B.G.; Kini, R.M. Expression pattern of three-finger toxin and phospholipase A2 genes in the venom glands of two sea snakes, Lapemis curtus and Acalyptophis peronii: Comparison of evolution of these toxins in land snakes, sea kraits and sea snakes. BMC Evol. Biol. 2007, 7, 175. [Google Scholar] [CrossRef]
- Takasaki, C.; Kimura, S.; Kokubun, Y.; Tamiya, N. Isolation, properties and amino acid sequences of a phospholipase A2 and its homologue without activity from the venom of a sea snake, Laticauda colubrina, from the Solomon Islands. Biochem. J. 1988, 253, 869–875. [Google Scholar] [CrossRef]
- Ratanabanangkoon, K.; Tan, K.Y.; Eursakun, S.; Tan, C.H.; Simsiriwong, P.; Pamornsakda, T.; Wiriyarat, W.; Klinpayom, C.; Tan, N.H. A simple and novel strategy for the production of a pan-specific antiserum against Elapid snakes of Asia. PLoS Negl. Trop. Dis. 2016, 10, e0004565. [Google Scholar] [CrossRef]
- Chetty, N.; Du, A.; Hodgson, W.C.; Winkel, K.; Fry, B.G. The in vitro neuromuscular activity of Indo-Pacific sea-snake venoms: Efficacy of two commercially available antivenoms. Toxicon 2004, 44, 193–200. [Google Scholar] [CrossRef]
- Tan, K.Y.; Tan, C.H.; Fung, S.Y.; Tan, N.H. neutralization of the principal toxins from the venoms of Thai Naja kaouthia and Malaysian Hydrophis schistosus: Insights into toxin-specific neutralization by two different antivenoms. Toxins 2016, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Tan, N.H.; Tan, K.Y.; Kwong, K.O. Antivenom cross-neutralization of the venoms of Hydrophis schistosus and Hydrophis curtus, two common sea snakes in Malaysian waters. Toxins 2015, 7, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Tan, K.Y.; Tan, N.H. A Protein Decomplexation strategy in snake venom proteomics. Methods Mol. Biol. 2019, 1871, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.Y.; Tan, C.H.; Sim, S.M.; Fung, S.Y.; Tan, N.H. Geographical venom variations of the Southeast Asian monocled cobra (Naja kaouthia): Venom-induced neuromuscular depression and antivenom neutralization. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016, 185–186, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Barber, C.M.; Isbister, G.K.; Hodgson, W.C. Alpha neurotoxins. Toxicon 2013, 66, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.H.; Wong, K.Y.; Tan, C.H. Venomics of Naja sputatrix, the Javan spitting cobra: A short neurotoxin-driven venom needing improved antivenom neutralization. J. Proteom. 2017, 157, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.Y.; Tan, C.H.; Tan, N.H. Venom and Purified Toxins of the Spectacled Cobra (Naja naja) from Pakistan: Insights into Toxicity and Antivenom Neutralization. Am. J. Trop. Med. Hyg. 2016, 94, 1392–1399. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.Y.; Tan, C.H.; Tan, K.Y.; Quraishi, N.H.; Tan, N.H. Elucidating the biogeographical variation of the venom of Naja naja (spectacled cobra) from Pakistan through a venom-decomplexing proteomic study. J. Proteom. 2018, 175, 156–173. [Google Scholar] [CrossRef] [PubMed]
- Oh, A.M.F.; Tan, C.H.; Ariaranee, G.C.; Quraishi, N.; Tan, N.H. Venomics of Bungarus caeruleus (Indian krait): Comparable venom profiles, variable immunoreactivities among specimens from Sri Lanka, India and Pakistan. J. Proteom. 2017, 164, 1–18. [Google Scholar] [CrossRef]
- Tan, K.Y.; Liew, J.L.; Tan, N.H.; Quah, E.S.H.; Ismail, A.K.; Tan, C.H. Unlocking the secrets of banded coral snake (Calliophis intestinalis, Malaysia): A venom with proteome novelty, low toxicity and distinct antigenicity. J. Proteom. 2018. [Google Scholar] [CrossRef]
- Lomonte, B.; Rey-Suarez, P.; Fernandez, J.; Sasa, M.; Pla, D.; Vargas, N.; Benard-Valle, M.; Sanz, L.; Correa-Netto, C.; Nunez, V.; et al. Venoms of Micrurus coral snakes: Evolutionary trends in compositional patterns emerging from proteomic analyses. Toxicon 2016, 122, 7–25. [Google Scholar] [CrossRef] [PubMed]
- Doley, R.; Zhou, X.; Kini, R.M. Snake Venom Phospholipase A2 Enzymes; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Geh, S.L.; Toh, H.T. Ultrastructural changes in skeletal muscle caused by a phospholipase A2 fraction isolated from the venom of a sea snake, Enhydrina schistosa. Toxicon 1978, 16, 633–643. [Google Scholar] [CrossRef]
- Tan, K.Y.; Tan, C.H.; Chanhome, L.; Tan, N.H. Comparative venom gland transcriptomics of Naja kaouthia (monocled cobra) from Malaysia and Thailand: Elucidating geographical venom variation and insights into sequence novelty. PeerJ 2017, 5, e3142. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Tan, K.Y.; Fung, S.Y.; Tan, N.H. Venom-gland transcriptome and venom proteome of the Malaysian king cobra (Ophiophagus hannah). BMC Genom. 2015, 16, 687. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Morita, T. Structure and function of snake venom cysteine-rich secretory proteins. Toxicon 2004, 44, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Heyborne, W.H.; Mackessy, S.P. Cysteine-rich secretory proteins in reptile venoms. In Handbook of Venoms and Toxins of Reptiles; Mackessy, S.P., Ed.; CRC Press: Boca Raton, FL, USA, 2009; pp. 325–336. [Google Scholar]
- Tan, C.H.; Tan, N.H. Toxinology of Snake Venoms: The Malaysian Context. In Snake Venoms; Gopalakrishnakone, P., Inagaki, H., Mukherjee, A.K., Rahmy, T.R., Vogel, C.-W., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 1–37. [Google Scholar] [CrossRef]
- Tan, N.H.; Ponnudurai, G. A comparative study of the biological properties of some sea snake venoms. Comp. Biochem. Physiol. B 1991, 99, 351–354. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, K.Y.; Tan, N.H. Revisiting Notechis scutatus venom: On shotgun proteomics and neutralization by the "bivalent" Sea Snake Antivenom. J. Proteom. 2016, 144, 33–38. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Lomonte, B.; León, G.; Alape-Girón, A.; Flores-Díaz, M.; Sanz, L.; Angulo, Y.; Calvete, J.J. Snake venomics and antivenomics: Proteomic tools in the design and control of antivenoms for the treatment of snakebite envenoming. J. Proteom. 2009, 72, 165–182. [Google Scholar] [CrossRef] [Green Version]
- Faisal, T.; Tan, K.Y.; Sim, S.M.; Quraishi, N.; Tan, N.H.; Tan, C.H. Proteomics, functional characterization and antivenom neutralization of the venom of Pakistani Russell’s viper (Daboia russelii) from the wild. J. Proteom. 2018, 183, 1–13. [Google Scholar] [CrossRef]
- Tan, C.H.; Tan, K.Y.; Yap, M.K.; Tan, N.H. Venomics of Tropidolaemus wagleri, the sexually dimorphic temple pit viper: Unveiling a deeply conserved atypical toxin arsenal. Sci. Rep. 2017, 7, 43237. [Google Scholar] [CrossRef]
- Tan, C.H.; Liew, J.L.; Tan, K.Y.; Tan, N.H. Assessing SABU (Serum Anti Bisa Ular), the sole Indonesian antivenom: A proteomic analysis and neutralization efficacy study. Sci. Rep. 2016, 6, 37299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard-Jones, N. A CIOMS ethical code for animal experimentation. WHO Chron 1985, 39, 51–56. [Google Scholar] [PubMed]
- Finney, D.J. Probit Analysis, 2nd ed.; Cambridge University Press: Cambridge, UK, 1952. [Google Scholar]
- Tan, K.Y.; Tan, N.H.; Tan, C.H. Venom proteomics and antivenom neutralization for the Chinese eastern Russell’s viper, Daboia siamensis from Guangxi and Taiwan. Sci. Rep. 2018, 8, 8545. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | Distinct Peptides | MS/MS Search Score | Species (as Per Annotation in Database) | Database Accession a | Protein Name (as Per Annotation in Database) | Relative Abundance (%) b |
|---|---|---|---|---|---|---|
| 1 | 2 | 28.12 | Hydrophis curtus | P68416 | Short neurotoxin 1 | 22.89 |
| 2 | 3 | 62.47 | Hydrophis curtus | A3FM53 | Long neurotoxin 2 | 0.49 |
| 3 | 5 | 114.27 | Hydrophis curtus | Q8UW29 | Long neurotoxin 1 | 1.10 |
| 4 | 84.14 | Hydrophis curtus | A3FM53 | Long neurotoxin 2 | 1.32 | |
| 2 | 40.42 | Ophiophagus hannah | Q53B58 | Long neurotoxin OH-55 | 0.33 | |
| 1 | 21.62 | Ophiophagus hannah | Q53B57 | Long neurotoxin OH-56 | 0.21 | |
| 4 | 5 | 92.36 | Hydrophis curtus | Q8UW31 | Acidic phospholipase A2 57 | 0.08 |
| 2 | 46.47 | Enhydrina schistosa | Unigene22561_ESM | Acidic phospholipase A2 57 | 0.34 | |
| 2 | 43.43 | Hydrophis curtus | CL4079.Contig1_HCM | Basic phospholipase A2 73 | 0.07 | |
| 2 | 35.82 | Calliophis intestinalis | CL2932.Contig3_CIM | Acidic phospholipase A2 57 | 0.16 | |
| 3 | 66.02 | Enhydrina schistosa | P00610 | Basic phospholipase A2 | 0.45 | |
| 2 | 50.61 | Hydrophis curtus | Q8UW30 | Basic phospholipase A2 73 | 0.32 | |
| 1 | 20.76 | Hydrophis curtus | Unigene14087_HCM | Waprin-Rha1 | 0.11 | |
| 5 | 4 | 85.11 | Enhydrina schistosa | P00610 | Basic phospholipase A2 | 2.90 |
| 3 | 64.58 | Hydrophis curtus | Q8UW08 | Basic phospholipase A2 | 1.68 | |
| 3 | 60.39 | Hydrophis curtus | Q8UW30 | Basic phospholipase A2 73 | 2.78 | |
| 2 | 46.67 | Enhydrina schistosa | Unigene22561_ESM | Acidic phospholipase A2 57 | 1.80 | |
| 2 | 45.14 | Naja Naja | CL339.Contig1_NNSL | Basic phospholipase A2 | 2.85 | |
| 3 | 47.42 | Hydrophis curtus | Q8UW31 | Acidic phospholipase A2 57 | 0.48 | |
| 2 | 31.85 | Bungurus caeruleus | Unigene17389_BCSL | Acidic phospholipase A2 57 | 0.05 | |
| 2 | 24.1 | Calliophis intestinalis | CL2932.Contig3_CIM | Acidic phospholipase A2 57 | 0.15 | |
| 6 | 7 | 131.08 | Hydrophis curtus | Q8UW31 | Acidic phospholipase A2 57 | 28.11 |
| 2 | 46.96 | Hydrophis curtus | Q8UW08 | Basic phospholipase A2 | 0.98 | |
| 2 | 42.22 | Enhydrina schistosa | P00610 | Basic phospholipase A2 | 0.80 | |
| 2 | 41.03 | Bungarus caeruleus | Unigene17389_BCSL | Acidic phospholipase A2 57 | 15.26 | |
| 2 | 38.92 | Hydrophis curtus | Q8UW30 | basic phospholipase A2 73 | 0.91 | |
| 2 | 31.08 | Denisonia devisi | R4G2S8 | PLA-2-Den-2 | 1.82 | |
| 7 | 13 | 242.58 | Hydrophis curtus | CL2848.Contig2_HCM | Extracellular matrix protein 1 | 0.16 |
| 8 | 8 | 143.01 | Hydrophis curtus | CL2323.Contig2_HCM | Lysozyme C, milk isozyme-like | 0.77 |
| 9 | 8 | 148.33 | Hydrophis curtus | Q8UW11 | Cysteine-rich venom protein 2 | 3.28 |
| 6 | 111.6 | Enhydrina schistosa | CL131.Contig1_ESM | Cysteine-rich secretory protein | 3.42 | |
| 3 | 51.38 | Hydrophis curtus | Q8UW25 | Cysteine-rich venom protein 1 | 1.49 | |
| 10 | 23 | 394.95 | Ophiophagus hannah | P81383 | L-amino-acid oxidase | 0.18 |
| 20 | 351.67 | Hydrophis curtus | Unigene7803_HCM | Transferrin | 0.15 | |
| 12 | 197.37 | Ophiophagus hannah | I2C090 | Ophiophagus venom factor | 0.06 | |
| 8 | 139.98 | Ophiophagus hannah | CL304.Contig1_OHM | OVF precursor protein | 0.05 | |
| 9 | 144.94 | Hydrophis curtus | CL4561.Contig1_HCM | Glutathione peroxidase 3 | 0.07 | |
| 8 | 138.68 | Enhydrina schistosa | CL1665.Contig1_ESM | Sulfhydryl oxidase 1-like | 0.07 | |
| 7 | 124.2 | Hydrophis curtus | Q8UW11 | Cysteine-rich venom protein 2 | 0.81 | |
| 7 | 122.36 | Hydrophis curtus | CL4690.Contig9_HCM | Phospholipase A2 inhibitor | 0.20 | |
| 6 | 89.52 | Ovophis okinavensis | U3TDL2 | glutaminyl_cyclase | 0.02 | |
| 4 | 69.73 | Hydrophis curtus | Unigene19328_HCM | Phospholipase A2 inhibitor beta | 0.25 | |
| 4 | 54.96 | Hydrophis curtus | CL2048.Contig1_HCM | Multiple inositol polyphosphate phosphatase 1 | 0.05 | |
| 3 | 46.73 | Hydrophis curtus | CL1278.Contig2_HCM | Phospholipase B | 0.02 | |
| 3 | 45.91 | Enhydrina schistosa | CL560.Contig2_ESM | Carinatease-1 | 0.09 | |
| 2 | 37.92 | Hydrophis curtus | CL4690.Contig1_HCM | Scutatease-1 | 0.22 | |
| 2 | 35.54 | Hydrophis curtus | Unigene390_HCM | Zinc metalloproteinase-disintegrin-like NaMP | 0.05 | |
| 2 | 36.44 | Enhydrina schistosa | CL79.Contig2_ESM | ADP-ribosyl cyclase 1 | 0.01 | |
| 2 | 34.29 | Enhydrina schistosa | CL98.Contig1_ESM | Lysosomal Pro-X carboxypeptidase-like | 0.04 | |
| 2 | 32.92 | Hydrophis curtus | Unigene20804_HCM | Phosphodiesterase | 0.04 | |
| 2 | 32.47 | Hydrophis curtus | CL1263.Contig2_HCM | Golgi apparatus protein 1 | 0.02 | |
| 2 | 30.78 | Hydrophis curtus | Unigene23143_HCM | N-acetylglucosamine-6-sulfatase | 0.02 | |
| 2 | 29.28 | Ophiophagus hannah | CL2083.Contig1_OHM | OVF precursor protein | 0.03 |
| Protein Family/Protein Subtype | Fraction | Accession Code a | Relative Abundance (%) b |
|---|---|---|---|
| Phospholipases A2 | 62.00 | ||
| Acidic PLA2 | 48.26 | ||
| Acidic phospholipase A2 57 | 4,5,6 | Q8UW31 | 28.68 |
| PLA-2-Den-2 | 6 | R4G2S8 | 1.82 |
| Acidic phospholipase A2 57 | 5,6 | Unigene17389_BCSL | 15.31 |
| Acidic phospholipase A2 57 | 4,5 | Unigene22561_ESM | 2.15 |
| Acidic phospholipase A2 57 | 4,5 | CL2932.Contig3_CIM | 0.31 |
| Basic PLA2 | 13.74 | ||
| Basic phospholipase A2 | 4,5,6 | P00610 | 4.15 |
| Basic phospholipase A2 | 5,6 | Q8UW08 | 2.66 |
| Basic phospholipase A2 73 | 4,5,6 | Q8UW30 | 4.01 |
| Basic phospholipase A2 | 5 | CL339.Contig1_NNSL | 2.85 |
| Basic phospholipase A2 73 | 4 | CL4079.Contig1_HCM | 0.07 |
| Three-finger toxins | 26.33 | ||
| Short Neurotoxin | 22.89 | ||
| Short neurotoxin 1 | 1 | P68416 | 22.89 |
| Long Neurotoxin | 3.44 | ||
| Long neurotoxin 2 | 2,3 | A3FM53 | 1.81 |
| Long neurotoxin OH-56 | 3 | Q53B57 | 0.21 |
| Long neurotoxin OH-55 | 3 | Q53B58 | 0.33 |
| Long neurotoxin 1 | 3 | Q8UW29 | 1.10 |
| Cysteine-rich secretory protein | 9.00 | ||
| Cysteine-rich venom protein 2 | 9,10 | Q8UW11 | 4.09 |
| Cysteine-rich venom protein 1 | 9 | Q8UW25 | 1.49 |
| Cysteine-rich secretory protein | 9 | CL131.Contig1_ESM | 3.42 |
| Phospholipase A2 inhibitors | 0.45 | ||
| Phospholipase A2 inhibitor beta | 10 | Unigene19328_HCM | 0.25 |
| Phospholipase A2 inhibitor | 10 | CL4690.Contig9_HCM | 0.20 |
| Snake venom metalloproteinases | 0.36 | ||
| Zinc metalloproteinase-disintegrin-like NaMP | 10 | Unigene390_HCM | 0.05 |
| Scutatease-1 | 10 | CL4690.Contig1_HCM | 0.22 |
| Carinatease-1 | 10 | CL560.Contig2_ESM | 0.09 |
| L-amino acid oxidase | 0.18 | ||
| L-amino-acid oxidase | 10 | P81383 | 0.18 |
| Ophiophagus venom factor | 0.14 | ||
| Ophiophagus venom factor | 10 | I2C090 | 0.06 |
| OVF precursor protein | 10 | CL304.Contig1_OHM | 0.05 |
| OVF precursor protein | 10 | CL2083.Contig1_OHM | 0.03 |
| Waprin | 0.11 | ||
| Waprin-Rha1 | 4 | Unigene14087_HCM | 0.11 |
| Phosphodiesterase | 0.04 | ||
| Phosphodiesterase | 10 | Unigene20804_HCM | 0.04 |
| Phospholipase B | 0.02 | ||
| Phospholipase B | 10 | CL1278.Contig2_HCM | 0.02 |
| Non-toxin | 1.38 | ||
| Extracellular matrix protein 1 | 7 | CL2848.Contig2_HCM | 0.16 |
| Lysozyme C, milk isozyme-like | 8 | CL2323.Contig2_HCM | 0.77 |
| Transferrin | 10 | Unigene7803_HCM | 0.15 |
| Glutathione peroxidase 3 | 10 | CL4561.Contig1_HCM | 0.07 |
| Sulfhydryl oxidase 1-like | 10 | CL1665.Contig1_ESM | 0.07 |
| Glutaminyl_cyclase | 10 | U3TDL2 | 0.02 |
| Multiple inositol polyphosphate phosphatase 1 | 10 | CL2048.Contig1_HCM | 0.05 |
| ADP-ribosyl cyclase 1 | 10 | CL79.Contig2_ESM | 0.01 |
| Lysosomal Pro-X carboxypeptidase-like | 10 | CL98.Contig1_ESM | 0.04 |
| Golgi apparatus protein 1 | 10 | CL1263.Contig2_HCM | 0.02 |
| N-acetylglucosamine-6-sulfatase | 10 | Unigene23143_HCM | 0.02 |
| Species | H. curtus (Penang) n = 10 Adults Dry Weight Per Milking: ~1–8 mg | H. curtus (January, Weipa) n = 11 Adults | H. curtus (June, Weipa) n = 10 Subadults | H. schistosus (Penang) n = 10 Adults | L. colubrina (Bali) n = Several | Aipysurus laevus (Broome) n = 4 | H. platura (Costa Rica) n = 84 | H. cyanocinctus (Hara) n = Several |
|---|---|---|---|---|---|---|---|---|
| i.v. LD50 (µg/g mouse) | 0.20 (0.18–0.24) | NA | NA | 0.07 (0.05–0.09) | 0.10 (0.08–0.12) | 0.15 (0.08–0.25) | 0.23 (note: 0.13 by i.p. route) | 0.132 |
| Reference | Current study | Neale et al. (2017) [20] | Tan et al. (2015b) [12] | Tan et al. (2017) [19] | Laustsen et al. (2015) [18] | Lomonte et al. (2014) [17] | Calvete et al. (2012) [16] | |
| Methods | C18 rpHPLC, in-solution tryptic digestion, LCMS/MS | C4 rpHPLC, in-solution tryptic digestion, LCMS/MS | C4 rpHPLC, in-solution tryptic digestion, LCMS/MS | C18 rpHPLC, in-gel tryptic digestion, MALDI TOF/TOF | C18 rpHPLC, in-solution tryptic digestion, LCMS/MS | C18 rpHPLC, in-gel tryptic digestion, MALDI TOF/TOF | C18 rpHPLC, in-gel tryptic digestion, LCMS/MS | C18 rpHPLC, N-terminal sequencing, in-gel tryptic digestion, CID-MS/MS |
| 3FTX | 26.33 | 30.44 | 40.43 | 70.5 | 66.14 | 25.3 | ~49.9 | 81.1 |
| SNTX | 22.89 | 8.33 | 20.76 | 55.8 | 16.94 | 25.3 | ~36 | 51.7 |
| LNTX | 3.44 | 22.11 | 19.67 | 14.7 | 48.9 | - | ~14 | 29.4 |
| CTX | - | - | - | - | 0.3 | - | - | |
| PLA2 | 62.00 | 66.7 | 54.50 | 27.5 | 33.3 | 71.2 | 32.9 | 18.9 |
| Acidic | 48.26 | 57.93 | 41.21 | 6.1 | 0.04 | 46.4 | NA | NA |
| Basic | 13.74 | 8.76 | 13.28 | 21.4 | 33.26 | 24.8 | NA | NA |
| Neutral | - | 0.01 | 0.007 | - | - | - | NA | NA |
| CRISP | 9.00 | 2.53 | 4.95 | 1.3 | 0.05 | 2.5 | 9.1 | - |
| LAAO | 0.18 | - | - | 0.2 | - | - | - | - |
| SVMP | 0.36 | 0.004 | 0.01 | 0.5 | - | - | ~0.9 | - |
| NUC | - | - | - | - | - | - | ~0.8 | - |
| CTL | - | 0.09 | 0.003 | - | - | - | - | - |
| PDE | 0.04 | - | - | - | - | - | - | - |
| PLB | 0.02 | - | - | - | - | - | - | - |
| PLA2 inhibitor | 0.45 | 0.01 | - | - | - | - | - | - |
| OVF | 0.14 | - | - | - | - | - | - | - |
| Waprin | 0.11 | - | - | - | - | - | - | - |
| Non-toxins/cellular proteins | 1.38 | 0.231 | 0.12 | - | - | 0.2 | 5.0 | - |
| Unknown | - | - | - | - | 0.57 | 0.8 | 1.4 | - |
| Venom/Fraction | Challenge Dose | i.v. LD50 (µg/g) a | ED50 (µL) b | ER50 (mg/mL) c | Potency, P (mg/mL) d | SSAV Protein Concentration (mg/mL) | Normalized P, n-P (mg/g) e |
|---|---|---|---|---|---|---|---|
| Venom | 5 | 0.20 (0.18–0.24) | 9.87 (7.98–12.21) | 2.53 (2.28–3.04) | 2.03 (1.83–2.43) | 217.2 ± 3.0 | 9.35 |
| F1_SNTx | 5 | 0.10 (0.08–0.12) | 25.82 (22.38–29.79) | 0.41 (0.34–0.51) | 0.34 (0.27–0.41) | 217.2 ± 3.0 | 1.57 |
| F3_LNTx | 5 | 0.24 (0.21–0.28) | 27.90 (25.36–30.70) | 0.95 (0.83–1.10) | 0.76 (0.66–0.88) | 217.2 ± 3.0 | 3.50 |
| F5 | - | >1 | - | - | - | - | - |
| F6 | - | >1 | - | - | - | - | - |
| F9 | - | >1 | - | - | - | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, C.H.; Tan, K.Y.; Ng, T.S.; Sim, S.M.; Tan, N.H. Venom Proteome of Spine-Bellied Sea Snake (Hydrophis curtus) from Penang, Malaysia: Toxicity Correlation, Immunoprofiling and Cross-Neutralization by Sea Snake Antivenom. Toxins 2019, 11, 3. https://doi.org/10.3390/toxins11010003
Tan CH, Tan KY, Ng TS, Sim SM, Tan NH. Venom Proteome of Spine-Bellied Sea Snake (Hydrophis curtus) from Penang, Malaysia: Toxicity Correlation, Immunoprofiling and Cross-Neutralization by Sea Snake Antivenom. Toxins. 2019; 11(1):3. https://doi.org/10.3390/toxins11010003
Chicago/Turabian StyleTan, Choo Hock, Kae Yi Tan, Tzu Shan Ng, Si Mui Sim, and Nget Hong Tan. 2019. "Venom Proteome of Spine-Bellied Sea Snake (Hydrophis curtus) from Penang, Malaysia: Toxicity Correlation, Immunoprofiling and Cross-Neutralization by Sea Snake Antivenom" Toxins 11, no. 1: 3. https://doi.org/10.3390/toxins11010003
APA StyleTan, C. H., Tan, K. Y., Ng, T. S., Sim, S. M., & Tan, N. H. (2019). Venom Proteome of Spine-Bellied Sea Snake (Hydrophis curtus) from Penang, Malaysia: Toxicity Correlation, Immunoprofiling and Cross-Neutralization by Sea Snake Antivenom. Toxins, 11(1), 3. https://doi.org/10.3390/toxins11010003