Co-Occurrence and Combinatory Effects of Alternaria Mycotoxins and Other Xenobiotics of Food Origin: Current Scenario and Future Perspectives

 ,
,  , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Natural Occurrence and Co-Occurrence of Alternaria Mycotoxins in Food
2.1. Co-Occurrence of Different Alternaria Toxins in Food
2.2. Co-Occurrence of Alternaria Toxins with Other Mycotoxins
3. Individual Toxicity of Main Alternaria Toxins and Combined Toxicity with Other Mycotoxins and Bioactive Compounds of Food Origin
3.1. Individual Toxicity of Alternaria Mycotoxins
3.1.1. Genotoxic Effects
3.1.2. Endocrine-Modulating and Other Toxic Effects
3.2. Combinatory Effects of Alternaria Mycotoxins
3.3. Combined Effects with Bioactive Food Constituents
4. The Key Role of Bioactive Compounds
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Berthiller, F.; Crews, C.; Dall’Asta, C.; de Saeger, S.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific Opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- Enyiukwu, D.N.; Awurum, A.N.; Nwaneri, J.A. Mycotoxins in Stored Agricultural Products: Implications to Food Safety and Health and Prospects of Plant-derived Pesticides as Novel Approach to their Management. Greener J. Microbiol. Antimicrob. 2014, 2, 32–48. [Google Scholar] [CrossRef]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; et al. Worldwide contamination of food-crops with mycotoxins : Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2019, 1–17. [Google Scholar] [CrossRef]
- Van der Fels-Klerx, H.J.; Liu, C.; Battilani, P. Modelling climate change impacts on mycotoxin contamination. World Mycotoxin J. 2016, 9, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Dellafiora, L.; Dall’Asta, C. Forthcoming challenges in mycotoxins toxicology research for safer food-a need for multi-omics approach. Toxins 2017, 9, 18. [Google Scholar] [CrossRef] [PubMed]
- Van Egmond, H.P.; Schothorst, R.C.; Jonker, M.A. Regulations relating to mycotoxins in food : Perspectives in a global and European context. Anal. Bioanal. Chem. 2007, 389, 147–157. [Google Scholar] [CrossRef]
- Ostry, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Escrivá, L.; Oueslati, S.; Font, G.; Manyes, L. Alternaria Mycotoxins in Food and Feed : An Overview. J. Food Qual. 2017, 2017, 1569748. [Google Scholar] [CrossRef]
- European Food Safety Authority; Arcella, D.; Eskola, M.; Gómez Ruiz, J.A. Dietary exposure assessment to Alternaria toxins in the European population. EFSA J. 2016, 14, 4654. [Google Scholar]
- European Commission. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. 2006, 5–24. [Google Scholar]
- Sanco, E. Summary report of the standing committee on the food chain and animal health held in brussels, 29 may 2012. European Commission 2012, 2012, 682729. [Google Scholar]
- EFSA. Call for Data on Alternaria Toxins in Food and Feed; EFSA: Parma, Italy, 2016; pp. 3–4. [Google Scholar]
- Alexander, J.; Benford, D.; Boobis, A.; Eskola, M.; Fink-Gremmels, J.; Fürst, P.; Heppner, C.; Schlatter, J.; van Leeuwen, R. Risk assessment of contaminants in food and feed. EFSA J. 2012, 10, s1004. [Google Scholar] [CrossRef]
- Bottalico, A.; Logrieco, A. Toxigenic Alternaria species of economic importance. In Mycotoxins in Agriculture and Food Safety; Sinha, K.K., Bhatnagar, D., Eds.; Marcel Dekker: New York, NY, USA, 1998; pp. 65–108. [Google Scholar]
- Scott, P.M. Analysis of Agricultural Commodities and Foods for Alternaria Mycotoxins. J. AOAC Int. 2001, 84, 1809–1817. [Google Scholar] [Green Version]
- Fernández-Cruz, M.L.; Mansilla, M.L.; Tadeo, J.L. Mycotoxins in fruits and their processed products: Analysis, occurrence and health implications. J. Adv. Res. 2010, 1, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Fraeyman, S.; Croubels, S.; Devreese, M.; Antonissen, G. Emerging fusarium and alternaria mycotoxins: Occurrence, toxicity and toxicokinetics. Toxins 2017, 9, 228. [Google Scholar] [CrossRef] [PubMed]
- Stinson, E.E.; Osman, S.F.; Heisler, E.G.; Siciliano, J.; Bills, D.D. Mycotoxin Production in Whole Tomatoes, Apples, Oranges, and Lemons. J. Agric. Food Chem. 1981, 29, 790–792. [Google Scholar] [CrossRef] [PubMed]
- Puntscher, H.; Cobankovic, I.; Marko, D.; Warth, B. Quantitation of free and modified Alternaria mycotoxins in European food products by LC-MS/MS. Food Control 2019, 102, 157–165. [Google Scholar] [CrossRef]
- Gotthardt, M.; Asam, S.; Gunkel, K.; Moghaddam, A.F.; Baumann, E.; Kietz, R.; Rychlik, M. Quantitation of Six Alternaria Toxins in Infant Foods Applying Stable Isotope Labeled Standards. Front. Microbiol. 2019, 10, 109. [Google Scholar] [CrossRef] [Green Version]
- Gratz, S.W. Do plant-bound masked mycotoxins contribute to toxicity? Toxins 2017, 9, 85. [Google Scholar] [CrossRef]
- Nathanail, A.V.; Syvähuoko, J.; Malachová, A.; Jestoi, M.; Varga, E.; Michlmayr, H.; Adam, G.; Sieviläinen, E.; Berthiller, F.; Peltonen, K. Simultaneous determination of major type A and B trichothecenes, zearalenone and certain modified metabolites in Finnish cereal grains with a novel liquid chromatography-tandem mass spectrometric method. Anal. Bioanal. Chem. 2015, 407, 4745–4755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellafiora, L.; Perotti, A.; Galaverna, G.; Buschini, A.; Dall’Asta, C. On the masked mycotoxin zearalenone-14-glucoside. Does the mask truly hide? Toxicon 2016, 111, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Dellafiora, L.; Galaverna, G.; Righi, F.; Cozzini, P.; Dall’Asta, C. Assessing the hydrolytic fate of the masked mycotoxin zearalenone-14-glucoside—A warning light for the need to look at the “maskedome”. Food Chem. Toxicol. 2017, 99, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Puntscher, H.; Kütt, M.L.; Skrinjar, P.; Mikula, H.; Podlech, J.; Fröhlich, J.; Marko, D.; Warth, B. Tracking emerging mycotoxins in food: Development of an LC-MS/MS method for free and modified Alternaria toxins. Anal. Bioanal. Chem. 2018, 410, 4481–4494. [Google Scholar] [CrossRef]
- Walravens, J.; Mikula, H.; Rychlik, M.; Asam, S.; Devos, T.; Ediage, E.N.; Di Mavungu, J.D.; Jacxsens, L.; Van Landschoot, A.; Vanhaecke, L.; et al. Validated UPLC-MS/MS Methods to Quantitate Free and Conjugated Alternaria Toxins in Commercially Available Tomato Products and Fruit and Vegetable Juices in Belgium. J. Agric. Food Chem. 2016, 64, 5101–5109. [Google Scholar] [CrossRef]
- Sanzani, S.M.; Gallone, T.; Garganese, F.; Caruso, A.G.; Amenduni, M.; Ippolito, A. Contamination of fresh and dried tomato by Alternaria toxins in southern Italy. Food Addit. Contam. Part A 2019, 36, 789–799. [Google Scholar] [CrossRef]
- Gambacorta, L.; Magistà, D.; Perrone, G.; Murgolo, S.; Logrieco, A.F.; Solfrizzo, M. Co-occurrence of toxigenic moulds, aflatoxins, ochratoxin A, Fusarium and Alternaria mycotoxins in fresh sweet peppers (Capsicum annuum) and their processed products. World Mycotoxin J. 2018, 11, 159–174. [Google Scholar] [CrossRef]
- Ansari, A.A.; Shrivastava, A.K. Natural occurrence of Alternaria mycotoxins in sorghum and ragi from North Bihar, India. Food Addit. Contam. 1990, 7, 815–820. [Google Scholar] [CrossRef]
- Müller, M.E.H.; Korn, U. Alternaria mycotoxins in wheat—A 10 years survey in the Northeast of Germany. Food Control 2013, 34, 191–197. [Google Scholar] [CrossRef]
- Lou, J.; Fu, L.; Peng, Y.; Zhou, L. Metabolites from Alternaria fungi and their bioactivities. Molecules 2013, 18, 5891–5935. [Google Scholar] [CrossRef]
- Shimshoni, J.A.; Cuneah, O.; Sulyok, M.; Krska, R.; Galon, N.; Sharir, B.; Shlosberg, A. Mycotoxins in corn and wheat silage in Israel. Food Addit. Contam. Part A 2013, 30, 1614–1625. [Google Scholar] [CrossRef] [PubMed]
- Varga, E.; Glauner, T.; Berthiller, F.; Krska, R.; Schuhmacher, R.; Sulyok, M. Development and validation of a (semi-) quantitative UHPLC-MS/MS method for the determination of 191 mycotoxins and other fungal metabolites in almonds, hazelnuts, peanuts and pistachios. Anal. Bioanal. Chem. 2013, 405, 5087–5104. [Google Scholar] [CrossRef] [PubMed]
- Mikušová, P.; Sulyok, M.; Šrobárová, A. Alternaria mycotoxins associated with grape berries in vitro and in situ. Biologia 2014, 69, 173–177. [Google Scholar] [CrossRef]
- Asam, S.A.; Konitzer, K.K.; Schieberle, P.; Rychlik, M. Stable Isotope Dilution Assays of Alternariol and Alternariol Monomethyl Ether in Beverages. J. Agric. Food Chem. 2009, 57, 5152–5160. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.; Manes, J.; Font, G.; Juan-García, A. Determination of mycotoxins in fruit berry by-products using QuEChERS extraction method. LWT Food Sci. Technol. 2017, 86, 344–351. [Google Scholar] [CrossRef]
- Zhao, K.; Shao, B.; Yang, D.; Li, F. Natural Occurrence of Four Alternaria Mycotoxins in Tomato- and Citrus-Based Foods in China. J. Agric. Food Chem. 2014, 63, 343–348. [Google Scholar] [CrossRef]
- Scott, P.M.; Lawrence, G.A.; Lau, B.P. Analysis of wines, grape juices and cranberry juices for Alternaria toxins. Mycotoxin Res. 2006, 22, 142–147. [Google Scholar] [CrossRef]
- Prelle, A.; Spadaro, D.; Garibaldi, A.; Gullino, L.M. A new method for detection of five alternaria toxins in food matrices based on LC—APCI-MS. Food Chem. 2013, 140, 161–167. [Google Scholar] [CrossRef]
- López, P.; Venema, D.; de Rijk, T.; de Kok, A.; Scholten, J.M.; Mol, H.G.J.; de Nijs, M. Occurrence of Alternaria toxins in food products in The Netherlands. Food Control 2016, 60, 196–204. [Google Scholar] [CrossRef]
- Veprikova, Z.; Zachariasova, M.; Dzuman, Z.; Zachariasova, A.; Fenclova, M.; Slavikova, P.; Vaclavikova, M.; Mastovska, K.; Hengst, D.; Hajslova, J. Mycotoxins in Plant-Based Dietary Supplements: Hidden Health Risk for Consumers. J. Agric. Food Chem. 2015, 63, 6633–6643. [Google Scholar] [CrossRef]
- Lau, B.P.; Scott, P.M.; Lewis, D.A.; Kanhere, S.R.; Cleroux, C.; Roscoe, V.A. Liquid chromatography—Mass spectrometry and liquid chromatography—Tandem mass spectrometry of the Alternaria mycotoxins alternariol and alternariol monomethyl ether in fruit juices and beverages. J. Chromatogr. A 2003, 998, 119–131. [Google Scholar] [CrossRef]
- Delgado, T.; Gόmez-Cordovés, C. Natural occurrence of alternariol and alternariol methyl ether in Spanish apple juice concentrates. J. Chromatogr. A 1998, 815, 93–97. [Google Scholar] [CrossRef]
- Pavón, M.Á.; Luna, A.; de la Cruz, S.; González, I.; Martín, R.; García, T. PCR-based assay for the detection of Alternaria species and correlation with HPLC determination of altenuene, alternariol and alternariol monomethyl ether production in tomato products. Food Control 2012, 25, 45–52. [Google Scholar] [CrossRef]
- Hickert, S.; Bergmann, M.; Ersen, S.; Cramer, B.; Humpf, H. Survey of Alternaria toxin contamination in food from the German market, using a rapid HPLC-MS/MS approach. Mycotoxin Res. 2016, 32, 7–18. [Google Scholar] [CrossRef]
- da Cruz Cabral, L.; Terminiello, L.; Pinto, V.F.; Nielsen, K.F.; Patriarca, A. Natural occurrence of mycotoxins and toxigenic capacity of Alternaria strains from mouldy peppers. Int. J. Food Microbiol. 2016, 236, 155–160. [Google Scholar] [CrossRef]
- Oviedo, M.S.; Barros, G.G.; Chulze, S.N.; Ramirez, M.L. Natural occurrence of alternariol and alternariol monomethyl ether in soya beans. Mycotoxin Res. 2012, 28, 169–174. [Google Scholar] [CrossRef]
- Juan, C.; Oueslati, S.; Mañes, J. Evaluation of Alternaria mycotoxins in strawberries : Quantification and storage condition. Food Addit. Contam. Part A 2016, 33, 861–868. [Google Scholar] [CrossRef]
- Magnani, R.F.; De Souza, G.D.; Rodrigues-Filho, E. Analysis of Alternariol and Alternariol Monomethyl Ether on Flavedo and Albedo Tissues of Tangerines (Citrus reticulata) with Symptoms of Alternaria Brown Spot. J. Agric. Food Chem. 2007, 55, 4980–4986. [Google Scholar] [CrossRef]
- Hasan, H.A.H. Alternaria mycotoxins in black rot lesion of tomato fruit: Conditions and regulation of their production. Mycopathologia 1995, 130, 171–177. [Google Scholar] [CrossRef]
- Noser, J.; Schneider, P.; Rother, M.; Schmutz, H. Determination of six Alternaria toxins with UPLC-MS/MS and their occurrence in tomatoes and tomato products from the Swiss market. Mycotoxin Res. 2011, 27, 265–271. [Google Scholar] [CrossRef]
- Zhao, K.; Shao, B.; Yang, D.; Li, F.; Zhu, J. Natural occurrence of Alternaria toxins in wheat-based products and their dietary exposure in China. PLoS ONE 2015, 10, e0132019. [Google Scholar] [CrossRef] [PubMed]
- Kayode, O.F.; Sulyok, M.; Fapohunda, S.O.; Ezekiel, C.N.; Krska, R.; Oguntona, C.R.B. Surveillance Mycotoxins and fungal metabolites in groundnut- and maize-based snacks from Nigeria. Food Addit. Contam. Part B 2013, 6, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Azcarate, M.P.; Patriarca, A.; Terminiello, L.; Fernandez Pinto, V. Alternaria Toxins in Wheat during the 2004 to 2005 Argentinean Harvest. J. Food Prot. 2008, 71, 1262–1265. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yoshizawa, T. Alternaria Mycotoxins in Weathered Wheat from China. J. Agric. Food Chem. 2000, 48, 2920–2924. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nie, J.; Yan, Z.; Li, Z.; Cheng, Y.; Chang, W. Occurrence and co-occurrence of mycotoxins in nuts and dried fruits from China. Food Control 2018, 88, 181–189. [Google Scholar] [CrossRef]
- Wei, D.; Wang, Y.; Jiang, D.; Feng, X.; Li, J.; Wang, M. Survey of Alternaria Toxins and Other Mycotoxins in Dried Fruits in China. Toxins 2017, 9, 200. [Google Scholar] [CrossRef]
- Ezekiel, C.N.; Sulyok, M.; Warth, B.; Krska, R. Multi-microbial metabolites in fonio millet (acha) and sesame seeds in Plateau State, Nigeria. Eur. Food Res. Technol. 2012, 235, 285–293. [Google Scholar] [CrossRef]
- Chulze, S.N.; Torres, A.M.; Dalcero, A.N.A.M.; Etcheverry, M.G.; Ramirez, M.L.; Farnochi, M.C. Alternaria Mycotoxins in Sunflower Seeds : Incidence and Distribution of the Toxins in Oil and Meal. J. Food Prot. 1995, 58, 1133–1135. [Google Scholar] [CrossRef]
- Hickert, S.; Hermes, L.; Marques, L.M.M.; Focke, C.; Cramer, B.; Lopes, N.P.; Flett, B.; Humpf, H.-U. Alternaria toxins in South African sunflower seeds : Cooperative study. Mycotoxin Res. 2017, 33, 309–321. [Google Scholar] [CrossRef]
- Bhat, R.; Rai, R.V.; Karim, A.A. Mycotoxins in Food and Feed: Present Status and Future Concerns. Compr. Rev. Food Sci. Food Saf. 2010, 9, 57–81. [Google Scholar] [CrossRef]
- Janić Hajnal, E.; Čolović, R.; Pezo, L.; Orčić, D.; Vukmirović, D.; Mastilović, J. Possibility of Alternaria toxins reduction by extrusion processing of whole wheat flour. Food Chem. 2016, 213, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Bauer, J.I.; Gross, M.; Gottschalk, C.; Usleber, E. Investigations on the occurrence of mycotoxins in beer. Food Control 2016, 63, 135–139. [Google Scholar] [CrossRef]
- Fanning, R.A.; Sheehan, F.; Leyden, C.; Duffy, N.; Iglesias-Martinez, L.F.; Carey, M.F.; Campion, D.P.; O’Connor, J.J. A role for adrenergic receptors in the uterotonic effects of ergometrine in isolated human term nonlaboring myometrium. Anesth. Analg. 2017, 124, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.E.H.; Urban, K.; Köppen, R.; Siegel, D.; Korn, U.; Koch, M. Mycotoxins as antagonistic or supporting agents in the interaction between phytopathogenic Fusarium and Alternaria fungi. World Mycotoxin J. 2015, 8, 311–321. [Google Scholar] [CrossRef]
- Juan, C.; Covarelli, L.; Beccari, G.; Colasante, V.; Mañes, J. Simultaneous analysis of twenty-six mycotoxins in durum wheat grain from Italy. Food Control 2016, 62, 322–329. [Google Scholar] [CrossRef]
- Zwickel, T.; Kahl, S.M.; Rychlik, M.; Müller, M.E.H. Chemotaxonomy of mycotoxigenic small-spored Alternaria fungi—Do multitoxin mixtures act as an indicator for species differentiation? Front. Microbiol. 2018, 9, 1368. [Google Scholar] [CrossRef]
- Pero, R.W.; Posner, H.; Blois, M.; Harvan, D.; Spalding, J.W. Toxicity of metabolites produced by the “Alternaria”. Environ. Health Perspect. 1973, 4, 87–94. [Google Scholar] [CrossRef]
- Smith, E.R.; Fredrickson, T.N.; Hadidian, Z. Toxic effects of the sodium and the N,N’-dibenzylethylenediamine salts of tenuazonic acid (NSC-525816 and NSC-82260). Cancer Chemother. Rep. 1968, 52, 579. [Google Scholar]
- Woodey, M.A.; Chu, F.S. Toxicology of Alternaria mycotoxins. In Alternaria Biology, Plant Diseases and Metabolites; Elsevier: Amsterdam, The Netherlands, 1992; pp. 409–434. [Google Scholar]
- Griffin, G.; Chu, F. Toxicity of the Alternaria Metabolites Alternariol, Alternariol Methyl Ether, Altenuene, and Tenuazonic Acid in the Chicken Embryo Assay. Appl. Environ. Microbiol. 1983, 46, 1420–1422. [Google Scholar]
- Giambrone, J.J.; Davis, N.D.; Diener, U.L. Effect of tenuazonic acid on young chickens. Poult. Sci. 1978, 57, 1554–1558. [Google Scholar] [CrossRef]
- Benford, D.J. Risk Assessment of Chemical Contaminants and Residues in Food. In Persistent Organic Pollutants and Toxic Metals in Foods; Woodhead Publishing: Cambridge, UK, 2017; pp. 3–13. ISBN 9780081006740. [Google Scholar]
- Chou, T.-C. Theoretical Basis, Experimental Design, and Computerized Simulation of Synergism and Antagonism in Drug Combination Studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef] [PubMed]
- Webb, J.L. Enzyme and Metabolic Inhibitors; Academic Press: New York, NY, USA, 1963. [Google Scholar]
- Chou, T.-C.; Talalayi, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzyme Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Fehr, M.; Pahlke, G.; Fritz, J.; Christensen, M.O.; Boege, F.; Altemöller, M.; Podlech, J.; Marko, D. Alternariol acts as a topoisomerase poison, preferentially affecting the IIα isoform. Mol. Nutr. Food Res. 2009, 53, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, L.; Wagner, J.; Metzler, M. Estrogenic and clastogenic potential of the mycotoxin alternariol in cultured mammalian cells. Food Chem. Toxicol. 2006, 44, 398–408. [Google Scholar] [CrossRef]
- Brugger, E.M.; Wagner, J.; Schumacher, D.M.; Koch, K.; Podlech, J.; Metzler, M.; Lehmann, L. Mutagenicity of the mycotoxin alternariol in cultured mammalian cells. Toxicol. Lett. 2006, 164, 221–230. [Google Scholar] [CrossRef]
- Schuchardt, S.; Ziemann, C.; Hansen, T. Combined toxicokinetic and in vivo genotoxicity study on Alternaria toxins. EFSA Support. Publ. 2014, 11, 679E. [Google Scholar] [CrossRef]
- Pommier, Y. Drugging Topoisomerases: Lessons and Challenges. ACS Chem. Biol. 2013, 8, 82–95. [Google Scholar] [CrossRef]
- Fernández-Blanco, C.; Font, G.; Ruiz, M.J. Role of quercetin on Caco-2 cells against cytotoxic effects of alternariol and alternariol monomethyl ether. Food Chem. Toxicol. 2016, 89, 60–66. [Google Scholar] [CrossRef]
- Tiessen, C.; Ellmer, D.; Mikula, H.; Pahlke, G.; Warth, B.; Gehrke, H.; Zimmermann, K.; Heiss, E.; Fröhlich, J.; Marko, D. Impact of phase I metabolism on uptake, oxidative stress and genotoxicity of the emerging mycotoxin alternariol and its monomethyl ether in esophageal cells. Arch. Toxicol. 2017, 91, 1213–1226. [Google Scholar] [CrossRef]
- Dellafiora, L.; Dall’Asta, C.; Cruciani, G.; Galaverna, G.; Cozzini, P. Molecular modelling approach to evaluate poisoning of topoisomerase I by alternariol derivatives. Food Chem. 2015, 189, 93–101. [Google Scholar] [CrossRef]
- Schwarz, C.; Kreutzer, M.; Marko, D. Minor contribution of alternariol, alternariol monomethyl ether and tenuazonic acid to the genotoxic properties of extracts from Alternaria alternata infested rice. Toxicol. Lett. 2012, 214, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, C.; Tiessen, C.; Kreutzer, M.; Stark, T.; Hofmann, T.; Marko, D. Characterization of a genotoxic impact compound in alternaria alternata infested rice as altertoxin II. Arch. Toxicol. 2012, 86, 1911–1925. [Google Scholar] [CrossRef] [PubMed]
- Fleck, S.C.; Burkhardt, B.; Pfeiffer, E.; Metzler, M. Alternaria toxins: Altertoxin II is a much stronger mutagen and DNA strand breaking mycotoxin than alternariol and its methyl ether in cultured mammalian cells. Toxicol. Lett. 2012, 214, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Fleck, S.C.; Sauter, F.; Pfeiffer, E.; Metzler, M.; Hartwig, A.; Köberle, B. DNA damage and repair kinetics of the Alternaria mycotoxins alternariol, altertoxin II and stemphyltoxin III in cultured cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2016, 798–799, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Aichinger, G.; Krüger, F.; Puntscher, H.; Preindl, K.; Warth, B.; Marko, D. Naturally occurring mixtures of Alternaria toxins: Anti-estrogenic and genotoxic effects in vitro. Arch. Toxicol. 2019, 93, 3021–3031. [Google Scholar] [CrossRef]
- Vejdovszky, K.; Hahn, K.; Braun, D.; Warth, B.; Marko, D. Synergistic estrogenic effects of Fusarium and Alternaria mycotoxins in vitro. Arch. Toxicol. 2017, 91, 1447–1460. [Google Scholar] [CrossRef]
- Dellafiora, L.; Warth, B.; Schmidt, V.; Del Favero, G.; Mikula, H.; Fröhlich, J.; Marko, D. An integrated in silico/in vitro approach to assess the xenoestrogenic potential of Alternaria mycotoxins and metabolites. Food Chem. 2018, 248, 253–261. [Google Scholar] [CrossRef]
- Stypuła-Trębas, S.; Minta, M.; Radko, L.; Jedziniak, P.; Posyniak, A. Nonsteroidal mycotoxin alternariol is a full androgen agonist in the yeast reporter androgen bioassay. Environ. Toxicol. Pharmacol. 2017, 55, 208–211. [Google Scholar] [CrossRef]
- Dellafiora, L.; Galaverna, G.; Cruciani, G.; Dall’Asta, C. A computational study toward the “personalized” activity of alternariol—Does it matter for safe food at individual level? Food Chem. Toxicol. 2019, 130, 199–206. [Google Scholar] [CrossRef]
- Frizzell, C.; Ndossi, D.; Kalayou, S.; Eriksen, G.S.; Verhaegen, S.; Sørlie, M.; Elliott, C.T.; Ropstad, E.; Connolly, L. An in vitro investigation of endocrine disrupting effects of the mycotoxin alternariol. Toxicol. Appl. Pharmacol. 2013, 271, 64–71. [Google Scholar] [CrossRef]
- Grover, S.; Lawrence, C.B. The Alternaria alternata mycotoxin alternariol suppresses lipopolysaccharide-induced inflammation. Int. J. Mol. Sci. 2017, 18, 1577. [Google Scholar] [CrossRef] [PubMed]
- Kollarova, J.; Cenk, E.; Schmutz, C.; Marko, D. The mycotoxin alternariol suppresses lipopolysaccharide-induced inflammation in THP-1 derived macrophages targeting the NF-κB signalling pathway. Arch. Toxicol. 2018, 92, 3347–3358. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Qiang, S. Environmental, genetic and cellular toxicity of tenuazonic acid isolated from Alternaria alternata. Afr. J. Biotechnol. 2008, 7, 1151–1156. [Google Scholar]
- Bensassi, F.; Gallerne, C.; El Dein, O.S.; Hajlaoui, M.R.; Bacha, H.; Lemaire, C. Combined effects of alternariols mixture on human colon carcinoma cells. Toxicol. Mech. Methods 2015, 25, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Vejdovszky, K.; Sack, M.; Jarolim, K.; Aichinger, G.; Somoza, M.M.; Marko, D. In vitro combinatory effects of the Alternaria mycotoxins alternariol and altertoxin II and potentially involved miRNAs. Toxicol. Lett. 2017, 267, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Blanco, C.; Font, G.; Ruiz, M.J. Interaction effects of enniatin B, deoxinivalenol and alternariol in Caco-2 cells. Toxicol. Lett. 2016, 241, 38–48. [Google Scholar] [CrossRef]
- McKinney, J.D.; Richard, A.; Waller, C.; Newman, M.C.; Gerberick, F. The Practice of Structure Activity Relationships (SAR) in Toxicology. Toxicol. Sci. 2000, 56, 8–17. [Google Scholar] [CrossRef] [Green Version]
- Juan-García, A.; Juan, C.; Manyes, L.; Ruiz, M.J. Binary and tertiary combination of alternariol, 3-acetyl-deoxynivalenol and 15-acetyl-deoxynivalenol on HepG2 cells: Toxic effects and evaluation of degradation products. Toxicol. In Vitro 2016, 34, 264–273. [Google Scholar] [CrossRef]
- Vejdovszky, K.; Warth, B.; Sulyok, M.; Marko, D. Non-synergistic cytotoxic effects of Fusarium and Alternaria toxin combinations in Caco-2 cells. Toxicol. Lett. 2016, 241, 1–8. [Google Scholar] [CrossRef]
- Rocha, O.; Ansari, K.; Doohan, F.M. Effects of trichothecene mycotoxins on eukaryotic cells: A review. Food Addit. Contam. 2005, 22, 369–378. [Google Scholar] [CrossRef]
- Dellafiora, L.; Galaverna, G.; Dall’Asta, C. In silico analysis sheds light on the structural basis underlying the ribotoxicity of trichothecenes—A tool for supporting the hazard identification process. Toxicol. Lett. 2017, 270, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Solhaug, A.; Karlsøen, L.M.; Holme, J.A.; Kristoffersen, A.B.; Eriksen, G.S. Immunomodulatory effects of individual and combined mycotoxins in the THP-1 cell line. Toxicol. In Vitro 2016, 36, 120–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pistol, G.C.; Gras, M.A.; Marin, D.E.; Israel-Roming, F.; Stancu, M.; Taranu, I. Natural feed contaminant zearalenone decreases the expressions of important pro-and anti-inflammatory mediators and mitogen-activated protein kinase/NF-κB signalling molecules in pigs. Br. J. Nutr. 2014, 111, 452–464. [Google Scholar] [CrossRef] [PubMed]
- Pestka, J.J. Deoxynivalenol-induced proinflammatory gene expression: Mechanisms and pathological sequelae. Toxins 2010, 2, 1300–1317. [Google Scholar] [CrossRef] [PubMed]
- Vejdovszky, K.; Schmidt, V.; Warth, B.; Marko, D. Combinatory estrogenic effects between the isoflavone genistein and the mycotoxins zearalenone and alternariol in vitro. Mol. Nutr. Food Res. 2017, 61, 1600526. [Google Scholar] [CrossRef]
- Dellafiora, L.; Aichinger, G.; Geib, E.; Sánchez-Barrionuevo, L.; Brock, M.; Cánovas, D.; Dall’Asta, C.; Marko, D. Hybrid in silico/in vitro target fishing to assign function to “orphan” compounds of food origin—The case of the fungal metabolite atromentin. Food Chem. 2019, 270, 61–69. [Google Scholar] [CrossRef]
- Dellafiora, L.; Ruotolo, R.; Perotti, A.; Cirlini, M.; Galaverna, G.; Cozzini, P.; Buschini, A.; Dall’Asta, C. Molecular insights on xenoestrogenic potential of zearalenone-14-glucoside through a mixed in vitro/in silico approach. Food Chem. Toxicol. 2017, 108, 257–266. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Alexander, J.; Barregard, L.; Bignami, M.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Appropriateness to set a group health-based guidance value for zearalenone and its modified forms. EFSA J. 2016, 14, 4425. [Google Scholar]
- Aichinger, G.; Beisl, J.; Marko, D. Genistein and delphinidin antagonize the genotoxic effects of the mycotoxin alternariol in human colon carcinoma cells. Mol. Nutr. Food Res. 2017, 61, 1600462. [Google Scholar] [CrossRef]
- Chen, W.; Lin, Y.C.; Ma, X.Y.; Jiang, Z.Y.; Lan, S.P. High concentrations of genistein exhibit pro-oxidant effects in primary muscle cells through mechanisms involving 5-lipoxygenase-mediated production of reactive oxygen species. Food Chem. Toxicol. 2014, 67, 72–79. [Google Scholar] [CrossRef]
- Fukumoto, L.R.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef] [PubMed]
- Braga, A.R.C.; Murador, D.C.; de Souza Mesquita, L.M.; de Rosso, V.V. Bioavailability of anthocyanins: Gaps in knowledge, challenges and future research. J. Food Compos. Anal. 2018, 68, 31–40. [Google Scholar] [CrossRef]
- Bitsch, R.; Netzel, M.; Frank, T.; Strass, G.; Bitsch, I. Bioavailability and Biokinetics of Anthocyanins From Red Grape Juice and Red Wine. J. Biomed. Biotechnol. 2004, 2004, 293–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aichinger, G.; Puntscher, H.; Beisl, J.; Kütt, M.L.; Warth, B.; Marko, D. Delphinidin protects colon carcinoma cells against the genotoxic effects of the mycotoxin altertoxin II. Toxicol. Lett. 2018, 284, 136–142. [Google Scholar] [CrossRef]
- Jan, A.T.; Kamli, M.R.; Murtaza, I.; Singh, J.B.; Ali, A.; Haq, Q.M.R. Dietary flavonoid quercetin and associated health benefits-An overview. Food Rev. Int. 2010, 26, 302–317. [Google Scholar] [CrossRef]
- Jeong, Y.M.; Choi, Y.G.; Kim, D.S.; Park, S.H.; Yoon, J.A.; Kwon, S.B.; Park, E.S.; Park, K.C. Cytoprotective effect of green tea extract and quercetin against hydrogen peroxide-induced oxidative stress. Arch. Pharm. Res. 2005, 28, 1251–1256. [Google Scholar] [CrossRef]
- Vila-donat, P.; Fernández-blanco, C.; Sagratini, G.; Font, G.; Ruiz, M.-J. Effects of Soyasaponin I and soyasaponins-rich extract on the Alternariol- induced cytotoxicity on Caco-2 cells. Food Chem. Toxicol. 2015, 77, 44–49. [Google Scholar] [CrossRef]
- Rosenblum, E.R.; Stauber, R.E.; Van Thiel, D.H.; Campbell, I.M.; Gavaler, J.S. Assessment of the Estrogenic Activity of Phytoestrogens Isolated from Bourbon and Beer. Alcohol. Clin. Exp. Res. 1993, 17, 1207–1209. [Google Scholar] [CrossRef]
- Karabín, M.; Hudcová, T.; Jelínek, L.; Dostálek, P. Biologically Active Compounds from Hops and Prospects for Their Use. Compr. Rev. Food Sci. Food Saf. 2016, 15, 542–567. [Google Scholar] [CrossRef] [Green Version]
- Milligan, S.R.; Kalita, J.C.; Pocock, V.; Van De Kauter, V.; Stevens, J.F.; Deinzer, M.L.; Rong, H.; De Keukeleire, D. The Endocrine Activities of 8-Prenylnaringenin and Related Hop (Humulus lupulus L.) Flavonoids. J. Clin. Endocrinol. Metab. 2000, 85, 4912–4915. [Google Scholar] [CrossRef]
- Aichinger, G.; Beisl, J.; Marko, D. The Hop Polyphenols Xanthohumol and 8-Prenyl-Naringenin Antagonize the Estrogenic Effects of Fusarium Mycotoxins in Human Endometrial Cancer Cells. Front. Nutr. 2018, 5, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, R.E.; Bomser, J.A.; Min, D.B. Bioactivity of Resveratrol. Compr. Rev. Food Sci. Food Saf. 2006, 5, 65–70. [Google Scholar] [CrossRef]
- De Waard, W.J.; Aarts, J.M.M.J.G.; Peijnenburg, A.C.M.; De Kok, T.M.C.M.; Van Schooten, F.J.; Hoogenboom, L.A.P. Ah receptor agonist activity in frequently consumed food items. Food Addit. Contam. Part A 2008, 25, 779–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreck, I.; Deigendesch, U.; Burkhardt, B.; Marko, D.; Weiss, C. The Alternaria mycotoxins alternariol and alternariol methyl ether induce cytochrome P450 1A1 and apoptosis in murine hepatoma cells dependent on the aryl hydrocarbon receptor. Arch. Toxicol. 2012, 86, 625–632. [Google Scholar] [CrossRef]
- Zhang, S.; Qin, C.; Safe, S.H. Flavonoids as aryl hydrocarbon receptor agonists/antagonists: Effects of structure and cell context. Environ. Health Perspect. 2003, 111, 1877–1882. [Google Scholar] [CrossRef]
- Ashida, H.; Fukuda, I.; Yamashita, T.; Kanazawa, K. Flavones and flavonols at dietary levels inhibit a transformation of aryl hydrocarbon receptor induced by dioxin. FEBS Lett. 2000, 476, 213–217. [Google Scholar] [CrossRef]
- Bandele, O.J.; Clawson, S.J.; Osheroff, N. Dietary polyphenols as topoisomerase II poisons: B ring and C ring substituents determine the mechanism of enzyme-mediated DNA cleavage enhancement. Chem. Res. Toxicol. 2008, 21, 1253–1260. [Google Scholar] [CrossRef]
- López-Lázaro, M.; Willmore, E.; Austin, C.A. The dietary flavonoids myricetin and fisetin act as dual inhibitors of DNA topoisomerases I and II in cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2010, 696, 41–47. [Google Scholar] [CrossRef]
- Bandele, O.J.; Osheroff, N. Bioflavonoids as poisons of human topoisomerase II alpha and II beta. Biochemistry 2007, 46, 6097–6108. [Google Scholar] [CrossRef]
- Leone, S.; Basso, E.; Polticelli, F.; Cozzi, R. Resveratrol acts as a topoisomerase II poison in human glioma cells. Int. J. Cancer 2012, 131, E173–E178. [Google Scholar] [CrossRef]


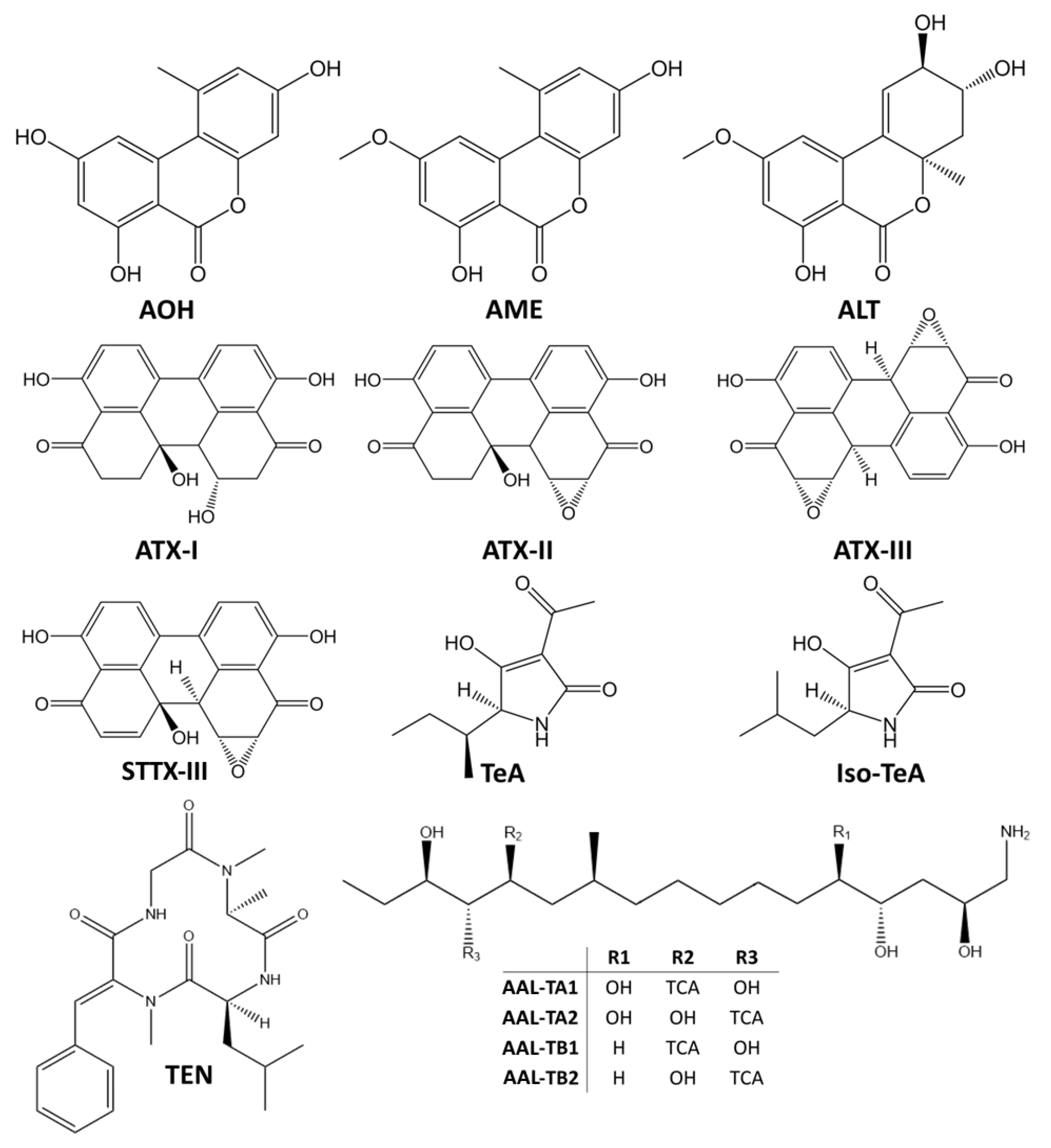{kind=link}
| Food/Foodstuff | Alternaria Mycotoxins | Reference | ||||||
|---|---|---|---|---|---|---|---|---|
| AOH | AME | ALT | TeA | TEN | ATX-I | Other | ||
| Fruits, Vegetables and Derivatives | ||||||||
| Apple | X | X | X | X | X | [19] | ||
| Apple juice | X | X | [36,43] | |||||
| Apple juice (concentrated) | X | X | [44] | |||||
| Apple-pear-cherry (puree infant formula) | X | X | X | X | – | ALP (–) | [21] | |
| Berry juice | X | X | – | [37] | ||||
| Cherry-banana (puree infant formula) | X | X | X | – | – | ALP (–) | [21] | |
| Cranberry juice | X | X | [39] | |||||
| Cranberry nectar | X | X | [43] | |||||
| Citrus juice | – | X a | X a | – | [38] | |||
| Grape juice | X | X | [39] | |||||
| Ketchup | X a | – | X a | [45] | ||||
| Ketchup | X | X | X | X | [38] | |||
| Mixed juice (fruits and vegetables) | X | X a | X a | X | – | Iso-ALT (–), AAL TA1 (–), AAL TA2(–) | [46] | |
| Orange juice | X | X | [36] | |||||
| Pepper | X | X | X | [47] | ||||
| Prune nectar | X | X | [43] | |||||
| Soya beans | X | X | [48] | |||||
| Strawberry | X a | X a | [49] | |||||
| Sweet pepper | X | X | X | X | [29] | |||
| Tangerine (flavedo) | X | X | [50] | |||||
| Tomato | X | X | X | X | – | [19] | ||
| Tomato | X a | X a | X a | – | ATX-II (–) | [51] | ||
| Tomato | X | X | X | [45] | ||||
| Tomato (dried) | X a | X a | X a | X a | [52] | |||
| Tomato (puree and ketchup) | X | X | – | X | X a | – | Iso-ALT (–), AAL TA1 (–), AAL TA2(–) | [46] |
| Tomato (sun-dried) | X | X | X | [45] | ||||
| Tomato juice | – | X | X | X | [38] | |||
| Tomato sauce | X | X | X a | X | X a | X | ALP (X), AOH-3-S (X a), AME-3-S (X) | [20] |
| Tomato sauce (puree infant formula) | X | X | X | X | – | ALP (–) | [21] | |
| Tomato soup (puree infant formula) | – | X | X | – | – | ALP (–) | [21] | |
| Vegetable juice | X | X | [36] | |||||
| Cereals and Derivatives | ||||||||
| Bakery products (wheat- and rye- based) | X | X | – | X | X | – | Iso-ALT (–), AAL TA1 (–), AAL TA2(–) | [46] |
| Bread | X a | X | X | X | [53] | |||
| Cereal grains | X a | X a | – | X a | [41] | |||
| Corn silage | X a | X a | X a | MACRO (X a) | [33] | |||
| Dried noodles | X a | X | X | X | [53] | |||
| Maize-based snacks | X | X | [54] | |||||
| Millet (infant formula) | X | X | X | X | – | ALP (–) | [21] | |
| Oat (infant formula) | – | X | X | X | – | ALP (–) | [21] | |
| Ragi | – | X | X | X | – | [30] | ||
| Rice (infant formula) | X | X | X | X | – | ALP (–) | [21] | |
| Sorghum | – | X | X | X | – | [30] | ||
| Spelt (infant formula) | X | X | X | X | X | ALP (–) | [21] | |
| Wheat | X a | X a | X a | X a | [31] | |||
| Wheat | X a | X a | X a | [55] | ||||
| Wheat (infant formula) | X | X | X | X | X | ALP (–) | [21] | |
| Wheat flour | X a | X | X | X | [53] | |||
| Wheat flour | X a | X | – | X | X a | X | ALP (X), AOH-3-S (-), AME-3-S (-) | [20] |
| Wheat silage | X a | X a | X a | MACRO (X a) | [33] | |||
| Weathered wheat | X | X | X | [56] | ||||
| Dried Fruits and Nuts | ||||||||
| Almonds | X a | X a | X a | [57] | ||||
| Almonds | X a | X a | X a | – | MACRO (X a) | [34] | ||
| Chestnuts | X a | X a | X a | [57] | ||||
| Dried figs | X a | X a | – | [57] | ||||
| Dried grape berries | X | X | X | X | X | X | ATX-II (X), MACRO (X) | [35] |
| Dried jujubes | X a | X a | X a | [57] | ||||
| Dried persimmons | X a | X a | – | [57] | ||||
| Dried raisins | X a | X a | – | [57] | ||||
| Dried raisins | X | X | – | X | – | [58] | ||
| Dried wolfberries | X a | X a | – | X | X | [58] | ||
| Hazelnuts | X a | X a | X a | [57] | ||||
| Hazelnuts | X | X | X | X a | MACRO (X) | [34] | ||
| Peanuts | X a | X a | X a | – | MACRO (X a) | [34] | ||
| Pine nuts | X a | X a | X a | [57] | ||||
| Pistachios | – | – | – | – | MACRO (X a) | [34] | ||
| Walnuts | X a | X a | X a | [57] | ||||
| Other Food and Foodstuff | ||||||||
| Beer | X a | – | X a | – | X a | [40] | ||
| Food supplement (antioxidants) | X a | X a | X a | X a | [42] | |||
| Food supplement (milk thistle) | X | X | X | X | [42] | |||
| Food supplement (phytoestrogens) | X | X | X a | X | [42] | |||
| Red wines | X | X | [39] | |||||
| Sesame seeds | X | X | X | [59] | ||||
| Sunflower seed oil | X | X | – | X | X | X a | ALP (X a), AOH-3-S (-), AME-3-S (-) | [20] |
| Sunflower seeds | – | – | – | X a | X a | [41] | ||
| Sunflower seeds | X | X | X | [60] | ||||
| Sunflower seeds | X | X | X a | X | X | X a | Iso-ALT (X a), AAL TA1 (–), AAL TA2(–) | [46] |
| Sunflower seeds | X a | X a | X | X | ALTSOH (X a), Val-TeA (X a) | [61] | ||
| Vegetable oils (rapeseed and sunflower seeds) | X | X | – | X a | X | – | Iso-ALT (–), AAL TA1 (–), AAL TA2(–) | [46] |
| White wines | X | X | [39] | |||||
| Wines | X a | – | – | X a | – | [41] | ||
| Food/Foodstuff | Co-Occurring Mycotoxins | Reference | |||
|---|---|---|---|---|---|
| 2 Mycotoxins | 3 Mycotoxins | 4 Mycotoxins | >4 Mycotoxins | ||
| Berry juices | AFB2, AME AFB2, AOH AOH, AME | AFB2, AFG2, AOH | AFB2, AFG2, AME, AOH AFG1, AFG2, AME, AOH AFB2, AFG1, AFG2, AME | [37] | |
| AFB2, AOH, OTA | |||||
| AFB2, AFG2, AME | |||||
| AFB2, AFG2, AOH | |||||
| AFG2, AME, AOH | |||||
| AFB2, AME, AOH | |||||
| Sweet pepper (Capsicum annuum) | AFB1, TeA | ZEN, TEN, TeA | FB2, TEN, TeA, ZEN | n 16 (NIV, AOH, TeA, HT-2, FB2, OTA, T-2, FB1, TEN, AME, AFB1, DON, AFG1, AFB2, AFG2 and ZEN) | [29] |
| Durum wheat | n.a. | n.a. | n.a. | n 7 (EN B, EN B1, EN A1, AME, DON, HT2 and T2) | [67] |
| Dried fruits (raisins, dried apricots, dates and wolfberries) | TeA, MPA TeA, TEN | n.a. | n.a. | n.a. | [58] |
| Maize-based snacks | n.a. | n.a. | n.a. | n 6 (FB1, FB2, FB3, BEAU, AME, EMOD) | [54] |
| Nuts and dried fruits | AFB2, TEN | AFB2, TEN, AME | ZEN, TEN, AOH, AME | n 8 (AFB1, AFB2, ENB, ENB1, OTB, TEN, AOH, AME) | [57] |
| AFG1, AME | AFB2, AOH, AME | ENA1, ENB, TEN, AME | |||
| ZEN, AOH | ENA1, ENB1, AME | AFB2, TEN, AOH, AME | |||
| BEA, AME | BEA, AOH, AME | AFB1, BEA, AOH, AME | |||
| T-2, AME | T-2, BEA, AME | AFG1, AFG2, ENB1, TEN | |||
| ENB1, TEN | AFB2, ENB, AOH | AFG1, ENB1, AOH, AME | |||
| Beer | AOH, ZEN | n.a. | Ergometrine, AOH, ZEN, DON | n.a. | [64] |
| Food supplements (milk thistle - based) | n.a. | n.a. | AOH, AME, TEN, MPA Other | n 14 (AOH, AME, TEN, 3-ADON, FUS-X, ENN-B, ENN-B1, ENN-A, ENN-A1, BEA, DON, HT-2, T-2, ZEN) etc. | [42] |
| Mycotoxin | Animal Species | Route of Exposure | LD50 (mg/kg b.w.) | Reference |
|---|---|---|---|---|
| AOH | Mouse (DBA/2) | intraperitoneal | >400 1 | [69] |
| AME | Mouse (DBA/2) | intraperitoneal | >400 1 | |
| TeA | Mouse | intravenous | 115 (female) | [70] |
| 162 (male) | ||||
| oral | 81 (female) | |||
| 186 (male) | ||||
| Mouse (ICR) | intravenous | 125 (male) | [71] | |
| intraperitoneal | 150 (male) | |||
| subcutaneous | 145 (male) | |||
| oral | 225 (male) | |||
| Rat | intravenous | 157 (female) | [70] | |
| 146 (male) | ||||
| oral | 168 (female) | |||
| 180 (male) | ||||
| Chicken embryo | injection | 548 2 | [72] | |
| White leghorn chicken | oral | 37.5 3 | [73] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crudo, F.; Varga, E.; Aichinger, G.; Galaverna, G.; Marko, D.; Dall’Asta, C.; Dellafiora, L. Co-Occurrence and Combinatory Effects of Alternaria Mycotoxins and Other Xenobiotics of Food Origin: Current Scenario and Future Perspectives. Toxins 2019, 11, 640. https://doi.org/10.3390/toxins11110640
Crudo F, Varga E, Aichinger G, Galaverna G, Marko D, Dall’Asta C, Dellafiora L. Co-Occurrence and Combinatory Effects of Alternaria Mycotoxins and Other Xenobiotics of Food Origin: Current Scenario and Future Perspectives. Toxins. 2019; 11(11):640. https://doi.org/10.3390/toxins11110640
Chicago/Turabian StyleCrudo, Francesco, Elisabeth Varga, Georg Aichinger, Gianni Galaverna, Doris Marko, Chiara Dall’Asta, and Luca Dellafiora. 2019. "Co-Occurrence and Combinatory Effects of Alternaria Mycotoxins and Other Xenobiotics of Food Origin: Current Scenario and Future Perspectives" Toxins 11, no. 11: 640. https://doi.org/10.3390/toxins11110640
APA StyleCrudo, F., Varga, E., Aichinger, G., Galaverna, G., Marko, D., Dall’Asta, C., & Dellafiora, L. (2019). Co-Occurrence and Combinatory Effects of Alternaria Mycotoxins and Other Xenobiotics of Food Origin: Current Scenario and Future Perspectives. Toxins, 11(11), 640. https://doi.org/10.3390/toxins11110640